
深圳湾红树林老鼠簕灌丛的光合固碳及其影响因子
2020-04-16 05:32:58林秋莲古伟唐
海洋技术学报 2020年1期
魏 龙,蔡 坚*,林秋莲,杨 琼,古伟唐
(1. 广东省林业科学研究院 广东省森林培育与保护利用重点实验室,广东 广州 510520; 2. 厦门大学 环境与生态学院,福建 厦门 361102; 3. 广东内伶仃-福田国家级自然保护区管理局,广东 深圳 518040;4. 深圳大学 生命与海洋科学学院,广东 深圳 518040)
红树林是生长在热带和亚热带海岸潮间带的木本植物群落[1]。老鼠簕(Acanthus ilicifolius)是爵床科老鼠簕属的常绿亚灌木,喜光而不耐荫蔽,在阳光充足处生长茂盛,高可达2 m,植株叶片坚硬光滑,为长椭圆形,边缘一般具刺状利齿,有泌盐现象。在我国,老鼠簕常构成红树林的林下灌丛[1-2]。过去,对于老鼠簕的研究主要集中在其地理分布特征、群落外貌及其结构、药用价值等方面[1-3];另一些研究则针对老鼠簕的种子萌发[4]、幼苗耐盐性[5]、潮汐淹水及其光照的适应性[6]等方面展开。温室内的模拟研究表明:由于适应于林下的生境,老鼠簕幼苗在光照强度达到45%时,叶片达到最大光合固碳能力,生长迅速;而极低的光照强度使老鼠簕幼苗生长缓慢[6]。
老鼠簕灌丛的生物量碳库是红树林林下植被碳库的主要组成,约占广东内伶仃-福田国家级自然保护区(114°03'E,22°32'N)红树林植被碳库的5%~10%[7-8]。野外条件下,老鼠簕还是先锋植物,在光照较弱的红树林及前沿光滩亦可分布。近10 年来,在深圳湾南岸的香港米铺湿地,老鼠簕的分布面积逐年增加,迅速占领中、高潮位的滩涂[9]。因此,随着该区域的老鼠簕灌丛的扩张,其生物量碳库对于红树林生物量碳库的贡献将大幅增加。在位于深圳湾北岸的福田红树林区,老鼠簕主要分布于林下、林窗和林缘,少数也分布于红树林前沿光滩[10]。然而,不同生境的环境因子将对老鼠簕植株的固碳效率产生不同影响,进而可能改变其生物量碳库的累积。在以造林增汇为目标的红树林造林中,物种配置和群落结构组成对于生物量碳库的影响极为显著,适当的林分改造大大提高了红树林碳储量[11]。在热带地区,如密克罗尼西亚的帕劳和雅浦,红树林的碳储量分别高达479~1 068 t·hm-2和853~1 385 t·hm-2,其复杂的林分结构和地被层碳库具有很大的贡献[12]。因此,本研究对不同生境下老鼠簕的固碳效率的比较,不仅能揭示其生物量固碳的演变特征和响应机制,还将对红树林造林的生物量碳库累积评估和造林实践提供理论依据。
1 材料与方法
1.1 样地概述
研究样地位于广东省深圳的广东内伶仃-福田国家级自然保护区(114°03'E,22°32'N)的观鸟屋西侧。该区域属于南亚热带季风气候,年均温22.5 ℃,最冷月(1 月)均温14.1 ℃;雨季为每年的3 月至9 月,年均降水量1 962 mm,年均相对湿度79%;平均日照时数为2 134 h;潮汐为不规则半日潮,平均潮差1.36 m(香港天文台,http://www.hko.gov.hk)。该区域内分布的天然林为秋茄(Kandelia obovata)-桐花树(Aegicerascorniculatum)群落、白骨壤(Avicennia marina)群落和人工引种的海桑(Sonneratia caseolaris)-无瓣海桑(Sonneratia apetala)群落。根据曾雪琴等[13]的调查结果,在天然秋茄-桐花树群落(秋茄林)、天然白骨壤群落(白骨壤林)、林中空地(林窗)、人工海桑-无瓣海桑群落的林缘和光滩等5 种生境下,各选取4 个1 m × 1 m 的样方,随机选取样方内2 棵植株测定。5 种生境的分布如图1 所示。

图1 深圳福田红树林的样地分布图
1.2 研究方法
1.2.1 生物量测定 在5 种样方内,各采集成熟老鼠簕植株2 棵作为水平样。将新鲜的植株分成根、茎、叶3 部分,把样品放入烘箱用105 ℃烘10 min进行杀青,60 ℃烘干至恒重,分别称取各植株根、茎、叶的干重。
1.2.2 光合特性和样品元素含量测定 选取5 种样方内的成熟植株各2 棵作为水平样,测定成熟叶片的光合特性。采用Li-6400(LI-6400XT; Li-Cor,Inc., Lincoln, NE, USA)便携式光合作用测定系统的外接红蓝LED 光源叶室,在晴天10:00-15:00 进行测定。选取枝条顶端向下的第二对健康成熟的叶片,在光照强度(PPFD)为1 000 μmol photons m-2s-1的光照条件下测定叶片的光合参数,主要包括叶片净光合速率(A),蒸腾速率(E)和气孔导度(gs)等。仪器使用开放式气路,CO2浓度为380 μmol mol-1。叶片WUE为同化速率和蒸腾速率比值(A/E),光合N 利用率PNUE为同化速率和叶片N 含量的比值(A/[N])。光响应曲线的测定采用外接红蓝LED 光源,设置10 个光照强度梯度,分别为1 600, 1 200, 800,400,200,100,50,20,10,0 μmol photons m-2s-1,每个梯度的光照强度持续至少2 min。最大光合速率(Amax),光补偿点(LCP)和光饱和点(LSP)通过Long等[14]的方法计算。所有参数可以通过公式(1)进行拟合[15]:
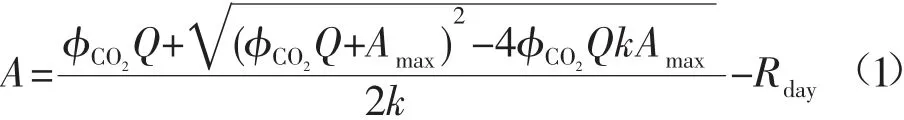
式中:A为净光合速率,μmol m-2s-1;ΦCO2为最大的表观量子产量,mol mol-1;Amax为最大光合速率,μmol CO2m-2s-1;Q为光照强度,μmol m-2s-1;k为常数(0<k<1);Rday为暗呼吸速率,μmol m-2s-1。
收集进行光合特性测定的叶片在60 ℃下烘干,研磨过60 目筛。用稳定同位素质谱仪(Thermo Finnigan MAT DELTAplus XP) 测 定 样 品 的δ13C(‰)。δ13C 用公式(2)表示:
式中:Rsample为样品的13C/12C 比率;Rstandard为Pee-Dee Belemnite 标准物的13C/12C 比率;叶片的碳稳定同位素比率(δ13C 值)代表了植物整个的生长季的水分利用率(WUE)[16]。通过稳定同位素δ13C 分析植物水分利用率(WUE)的公式为:
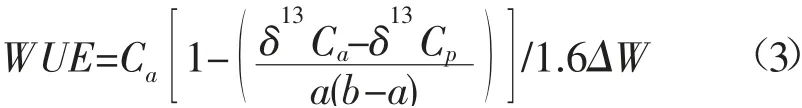
式中:Ca为大气CO2浓度;δ13Ca为大气CO2碳同位素比率;δ13Cp为植物叶片碳同位素比率;a为CO2从空气中扩散过程的碳同位素分差(4.4‰);b为RuBP 羧化酶在同化CO2过程中对13CO2的判别值(≈29‰);ΔW为叶片内外水蒸汽压差(可根据实验期间观测的大气温度和湿度计算获得)。
用德国Elementar 公司Vario EL III 型元素分析仪测定叶片总N 的含量(%)。叶片光合氮利用率
(PNUE)为光合速率A和叶片总N 含量的比值。
1.2.3 环境因子测定 分别选取夏秋季的每月月中的天气晴朗、光照稳定的中午,测定样地的实际光照强度,与空地光照强度比较,即获得其相对光照强度每月重复测定样方内的光照强度。对每月测定的日照强度计算平均值。在生物量测定的样方内,采集每个样地内的表层土壤(0~10 cm)样品,带回实验室自然风干、研磨。通过硝化法、分光光度法测定其N,P 含量[17]。按比例(土:水=1:5)取样,用去离子水充分浸泡、振荡,离心或过滤后取上清,分别用盐度计和pH 计测定其盐度和pH 值。5 种生境的环境因子如表1 所示。
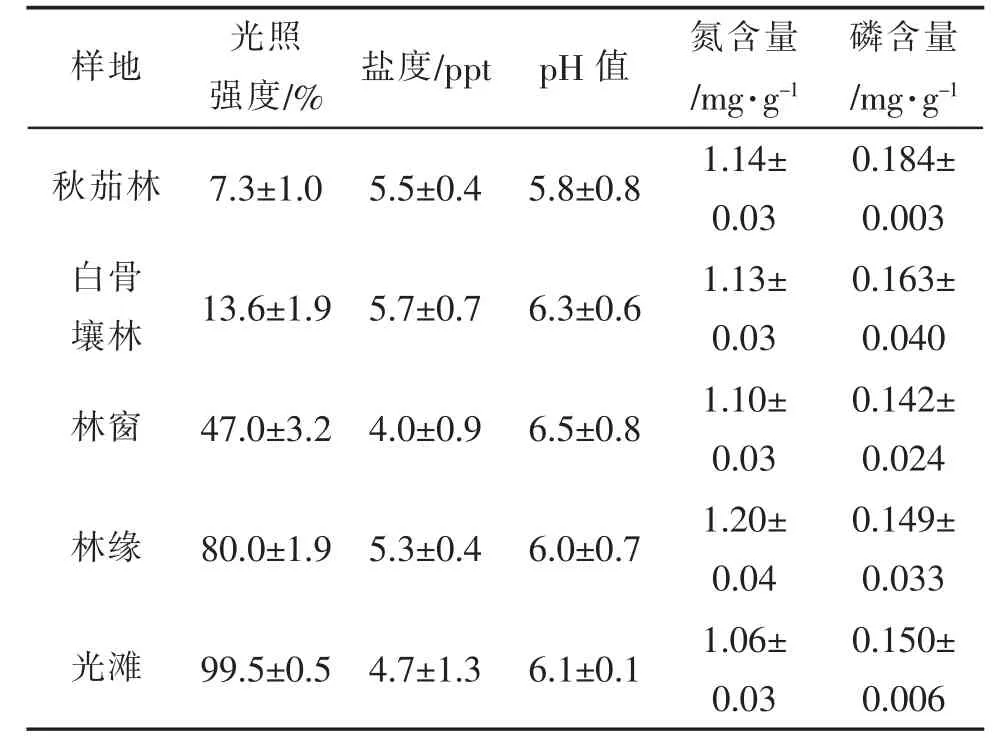
表1 5 种生境样地的环境因子
1.3 数据分析
应用统计软件SPSS17.0 对数据进行统计分析。对不同生境下的生物量和光合作用参数进行比较和差异显著性分析(One-way ANOVA,LSD 法),当p>0.05 为没有显著差异;采用线性回归对叶片水分利用效率(WUE)和叶片δ13C 值、植株总生物量和叶片光合氮利用效率(PNUE)进行回归;采用逐步回归对植株生物量、光合作用参数和生境的光照强度、土壤盐度和pH、土壤元素含量等进行回归分析。使用Sigmaplot10.0 绘图。
2 结果与分析
2.1 植株生物量及其分配特征
5 种生境的老鼠簕植株的地上和地下生物量及其分配情况如图2 所示。在林窗生境中的地上生物量显著高于其他样地,而秋茄和白骨壤林下的地上生物量最低。地下生物量最高值(4.62±0.93 g)出现在光滩的样方中。

图2 不同生境老鼠簕植株的地上生物量(AGB)和地下生物量(BGB)
2.2 植株成熟叶片的光合特性
2.2.1 叶片光合特性和水分利用率 5 种生境下老鼠簕成熟叶片的净光合速率A、气孔导度gs、蒸腾速率E和水分利用率WUE的变化趋势如图3 所示。其中,林窗、林缘和光滩3 种生境下A和gs达到最高,而林下生境不利于叶片的光合作用。蒸腾速率E在林下较低,而在光滩生境中达到最大值。WUE在林窗达到最大,而在林下和光滩生境最低。
在5 种不同生境中,老鼠簕成熟叶片WUE与δ13C 值呈现显著正相关(R2=0.301,P=0.015);总生物量与叶片PNUE具有极显著的正相关(R2=0.525,P=0.001),如图4 所示。
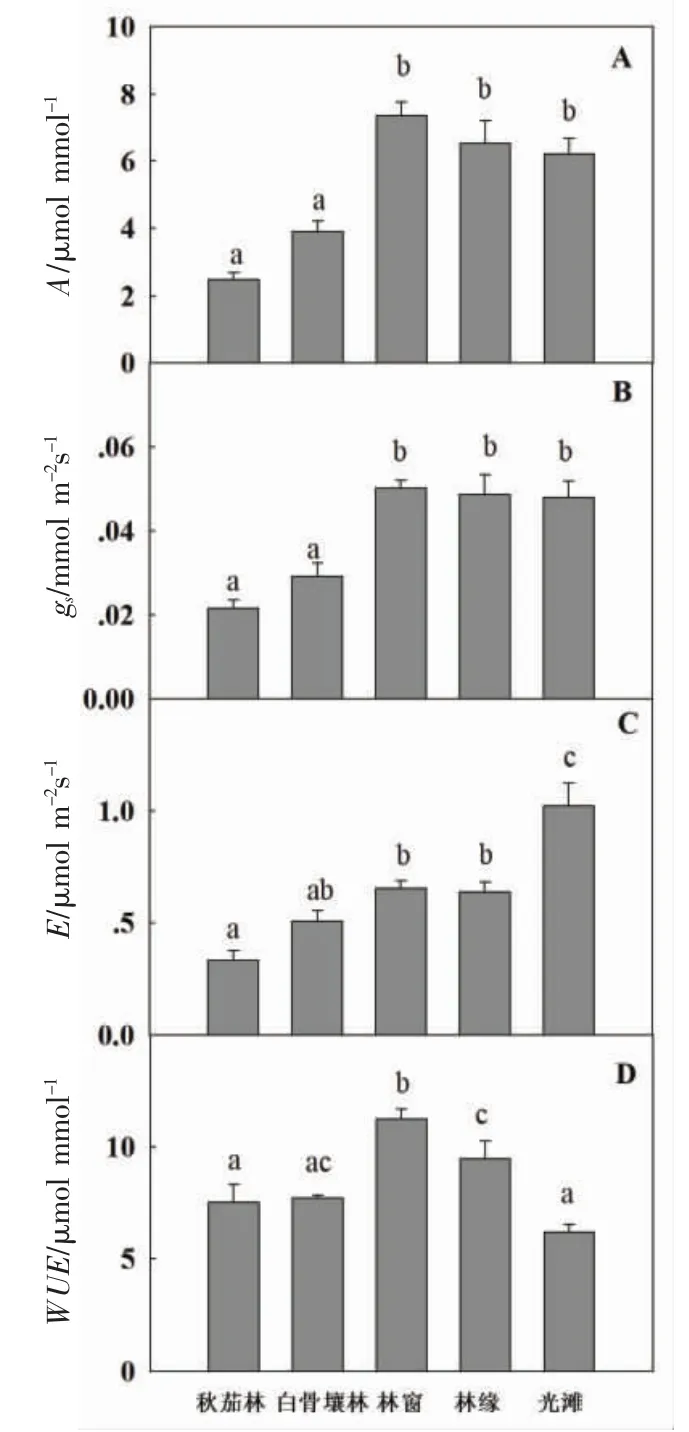
图3 不同生境老鼠簕植株成熟叶片的光合特性和水分利用率

图4 老鼠簕植株成熟叶片水分利用效率(WUE)和叶片δ13C值(A)、叶片的光合氮分利用效率(PNUE)和植株总生物量(B)的相关关系
2.2.2 叶片光响应曲线 5 种生境的老鼠簕植株成熟叶片的光响应曲线如图5 所示,图中,Amax为最大光合速率,LCP 为光补偿点,LSP 为光饱合点。林窗生境中的植株叶片最大光合速率(Amax)最高;而两种林下生境中的Amax均最低,叶片光合能力最弱。光补偿点(LCP)和光饱和点(LSP)的变化趋势与Amax相似。在林窗、林缘和光滩的生境中,叶片光合能力都处于较高水平,林窗的光照强度最有利于老鼠簕的光合作用。
2.3 影响植株光合固碳的关键生态因子
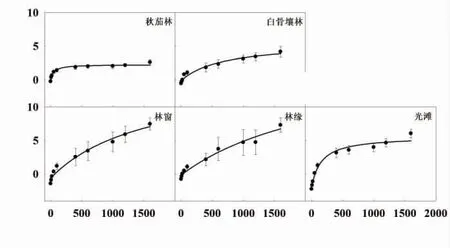
图5 不同生境下老鼠簕植株成熟叶片的光响应曲线
在光照强度、土壤pH 值、土壤盐度、土壤总氮和总磷含量等诸多环境因子中,老鼠簕植株的总生物量与光照强度存在显著的相关性(R2=0.515,P=0.009),与叶片光合速率也存在显著的相关性(R2=0.557,P<0.001)(参见表2);其他指标均不显著相关。
3 讨论
红树林是热带地区碳储量最丰富的森林之一[18]。近年来,国内外对红树林碳库的评估成为研究热点,并为红树林造林提供诸多信息。我国红树林是全球红树林天然分布的北界,在植被生物量碳库上较热带地区的碳库为低[19]。老鼠簕灌丛作为我国红树林的主要下木层植被,大大增加了红树林的植被碳库[8]。因此,对于红树林内、林缘和光滩等不同生境条件下老鼠簕的光合固碳能力开展研究,有助于从固碳机制上揭示老鼠簕灌丛碳库累积的影响因子,为红树林人工造林的物种配置模式和造林增汇措施提供理论依据。

表2 不同生境老鼠簕的生长/光合特性与主要环境因子的逐步回归分析结果
深圳福田红树林区的老鼠簕灌丛主要分布在秋茄林、白骨壤林内,也出现在林间空地、林缘和光滩上(图1)。从生物量的分配来看,在林窗、林缘和光滩的老鼠簕植株地上生物量累积大大高于林下生境(图2)。这与其叶片的光合速率有关(图3):在林窗、林缘和光滩生境下,成熟叶片能获得更多的光能,从而提高光合作用效率;同时,光滩的植株也增加了根生物量的比重,提高其对营养元素的吸收以支持高的光合作用消耗。植物的生物量分配反映了植物生长中资源和能量分配的一般特征[20-21]。在本研究中,在秋茄林和白骨壤林内,老鼠簕植株叶片的光合速率低、植株生物量较低,而林窗生境具有最高的总生物量。可见,老鼠簕植株的生物量分配策略与光照强度密切相关。
植物的气孔是叶片获取CO2的通道,也是叶片水分蒸腾的门户。叶片光合速率增大,消耗CO2增多,叶片通过调节气孔开度,增大气孔导度(gs)来加速外界CO2向叶细胞内的扩散,弥补CO2的消耗[16]。植物的光合速率和水分利用率的权衡,与植株有机碳固定密切相关,体现了植物生物量形成的物质基础[22]。在本研究中,林窗、林缘和光滩的老鼠簕叶片光合同化速率最大;但光滩的老鼠簕叶片蒸腾速率最大,说明光滩的高光强显著提高了叶片水分蒸腾,使得叶片的水分利用率降低。林窗和林缘的老鼠簕植株的水分利用更具保守性,这一特点符合老鼠簕在的分布特征,即老鼠簕在林窗和林缘生境下能进行最佳的光合同化和生物量积累。叶片δ13C反映了同化速率与气孔导度的平衡(内在水分利用效率),叶片的δ13C 值越高,植物水分利用效率就越高[23]。本研究中,老鼠簕叶片δ13C 值的变化与叶片WUE的变化是趋势一致,两者之间存在显著的正相关性(图4)。可见,中等光照强度下老鼠簕叶片较高的WUE是提高其碳同化的有效手段,而低光照条件对光合速率的抑制、高光照条件对蒸腾作用的促进均降低了其生物量积累。另外,叶片中N 直接或是间接参与光合作用的各个环节,对植物叶绿素、光合速率、暗反应的主要酶等有所影响[24]。PNUE代表单位叶片N 的净碳同化率,有效地指示了植物捕获碳源而付出的N。本研究中,老鼠簕叶片PNUE与植株总生物量呈现显著的正相关(图4),暗示了其投入光合作用的N 越多,光合捕获的碳也更多。
从叶片光合能力上,5 种不同生境中老鼠簕叶片光响应曲线的结果显示林下Amax低下;林窗生境的叶片Amax最高;林缘和光滩的Amax有所下降的趋势(图5);LCP 和LSP 的变化趋势也有类似趋势,且LSP 的最大值出现在林缘生境。这一结果与光合固碳速率的变化趋势较为一致,说明林窗和林缘是老鼠簕光合能力最佳的生境,最适于老鼠簕植株的生物量碳累积。
综合老鼠簕生长的5 种生境的环境因子,本研究发现光照强度是影响其生物量积累和光合碳同化的关键生态因子(表2)。在郁闭的冠层下,老鼠簕植株的生长和光合能力均受到抑制,而林窗和林缘生境的光合强度使其光合固碳达到最适。在光滩的强光照生境中,老鼠簕也能进行较高的光合固碳;但强光加速叶片水分丧失,降低WUE,进而抑制地上生物量累积。可见,光照强度是决定老鼠簕林下生长的重要因子。在林窗和林缘,光合速率和与光合速率有关的各项指标均处于高水平,说明在光强40%~80%的林窗和林缘环境中,老鼠簕可以通过较高的光合速率和水分利用率加速生长。
光是植物必需的资源之一,对植物的生长、发育和演化具有极其重要的作用,同时又是影响植物形态和生理功能、调控植物生长的重要环境因子[21,25]。红树林是滨海蓝碳的重要贡献者,其固碳效率显著高于热带雨林[18]。因此,对于红树林下木层植被碳库而言,虽然郁闭的红树林冠层下老鼠簕仅贡献了5%~10%的植被碳库[8],但在40%~80%的中等光照强度下老鼠簕对群落总碳库的贡献将达到最大值。可见,在深圳福田红树林中,老鼠簕灌丛的固碳率不容忽视,是该区域红树林蓝碳的重要组成。从滨海湿地生态修复的角度而言,物种配置对于红树林植被碳库极为重要[11]。因此,在我国红树林造林中,通过在林下种植老鼠簕灌丛可提高植被碳储量,也能提升红树林的固碳效率,起到更好的减排效果。从固碳效率提升的角度看,还可对冠层植物群落的郁闭度进行适当控制,使其光照强度维持在40%~80%之间,以提高老鼠簕的生物量固碳。
4 结论
深圳福田红树林区的林窗生境下,自然生长的老鼠簕灌丛光合能力、光合固碳速率和地上生物量累积均高于林下、林缘和光滩生境,而达到最大;林下生境最不利于老鼠簕植株固碳;光滩的老鼠簕植株具有较高的地下生物量累积,但由于其蒸腾速率高、水分利用率低下,其植株固碳和生物量低下。光照强度是决定5 种生境的老鼠簕植株固碳的关键因子。
猜你喜欢
西南林业大学学报(2022年5期)2022-10-17 09:56:38
儿童故事画报·自然探秘(2022年3期)2022-04-27 00:17:10
绿色科技(2020年20期)2020-11-20 01:56:34
农业机械学报(2020年1期)2020-03-13 00:27:38
疯狂英语·初中天地(2018年6期)2018-11-24 02:39:24
歌海(2018年4期)2018-05-14 12:46:15
西部林业科学(2018年4期)2018-02-14 22:43:50
江苏农业科学(2017年6期)2017-05-11 14:35:37
林业勘查设计(2017年1期)2017-03-11 16:49:57
西南林业大学学报(2014年6期)2014-07-24 19:02:11
