
日本鲐对马群系资源丰度预测研究
2020-04-16 05:34薛艳会陈新军汪金涛
海洋技术学报 2020年1期
薛艳会,陈新军,2,3,4,5,汪金涛,2,3,4,5*
(1. 上海海洋大学 海洋科学学院,上海 201306; 2. 农业部大洋渔业开发重点实验室,上海 201306; 3. 国家远洋渔业工程技术研究中心,上海 201306; 4. 大洋渔业资源可持续开发教育部重点实验室,上海 201306; 5. 农业部大洋渔业资源环境科学观测实验站,上海 201306)
日本鲐属于硬骨鱼纲,鲈形目,分布于我国的渤海、黄海、东海、南海以及西北太平洋沿岸海域,主要由我国(包括台湾省)、日本和朝鲜等国捕捞利用[1]。近几年来,它已成为重要的中上层鱼类资源,有研究[2]表明其性成熟年龄降低,约1 龄以上开始性成熟,对马群系的寿命多为6 龄[3]。日本鲐对马群系是包括东海、黄海群在内的对马暖流群系[4],其中东海西部群在春、夏季向东海、黄海近海洄游产卵,随后在其附近索饵,秋冬返回越冬场;黄海群主要来自东海中南部至钓鱼岛北部和日本九州西部外海两个越冬场;日本海西部群体的洄游路线尚不明了。
前人有不少关于日本鲐的研究,主要是针对近海海域。李纲等[5]对东海鲐鱼资源和渔场时空分布特征进行研究,发现鲐鱼主要作业渔场在经度上分布无差异,而在纬度上分布差异明显。李日嵩等[6]关于水温对东海鲐鱼补充量的研究表明,水温会影响到鲐鱼生长初期鱼卵孵化、仔鱼变态的时间及仔幼鱼的生长发育,进而影响其资源丰度。Akihiko Yatsu 等[7]的研究表明,太平洋年际振荡指数(Pacific Decadal Oscillation,PDO)对鲐鱼补充量有间接影响。PDO通过海气相互作用,可直接影响太平洋及周边地区气候的年际变化,PDO指数为正值是暖位相,为负值是冷位相,即当其正负值不同时,对鱼类资源量变动的影响不同。也有研究[8]表明,Nino3.4 海表温距平值(Sea Surface Temperature Anomaly,SSTA)对鲐鱼资源变化有明显影响。
灰色系统理论是研究渔业资源的一种极有效的分析手段和工具,原因是渔业资源具有的不确定性和变动性较其他资源大[6]。灰色预测模型在渔业研究中也得到广泛应用,对于渔业资源的管理起到了很好的指导作用。例如,段丁毓[9]、朱文涛[10]运用灰色系统,分别构建了秘鲁鳀资源量预测模型、西北太平洋秋刀鱼的资源丰度预测模型,都得到了比较好的预测结果。本文所研究的区域为包括日本海的整个对马群系,研究采用灰色关联分析和相关系数分析等方法探讨日本鲐对马群系资源丰度与其产卵场和索饵场的海表温度(Sea Surface Temperature,SST)、SSTA和PDO的关系,并基于选出的关键环境和气候因子建立多种灰色预测模型,为其今后的资源开发和可持续利用提供技术支撑。
1 材料和方法
1.1 材料来源
日本鲐资源量数据来源于2017 年6 月日本西海水产研究所对日本鲐对马群系资源量的评估报告[11]。时间为1973-2014 年,数据包括资源量和渔获量数据(图1)。

图1 1973-2013 年日本鲐对马群系资源量、渔获量统计
海洋环境数据包括产卵场和索饵场的SST,PDO,SSTA。SST下载自网站(http://iridl.ldeo.columbia.edu),空间分辨率为1°×1°,时间分辨率为月。PDO和SSTA来自网站(http://www.cpc.ncep.noaa.gov)。
PDO的冷暖位相不同,即冷暖年对渔业资源影响不同,所以在分析PDO这一气候因子对日本鲐资源量变动的影响时,采用冷暖年份分开的方式。在本研究中,PDO数据是1995-2014 年的,其中1995年、1998 年、2002 年、2006 年和2014 年为暖位相,1999 年、2001 年、2007 年、2013 年为冷位相。
根据前人研究[11],推测日本鲐有两个主要的产卵场和一个索饵场(图2),其空间范围分别为26°~31° N,122°~127° E(SG1);30°~35° N,128°~131° E(SG2);35°~38° N,127°~138° E(FG)。这3 块区域月平均SST分别表示为SSTSG1,SSTSG2,SSTFG。
1.2 关键环境因子选取
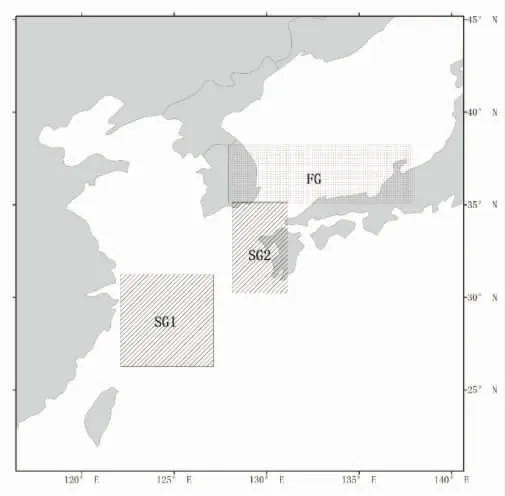
图2 日本鲐对马群系主要产卵场和索饵场分布图
计算2000-2014 年资源量与产卵场、索饵场月平均温度的相关系数;以2000-2014 年的资源量为母序列,SSTSG1、SSTSG2和SSTFG为子序列,计算灰色关联度,具体计算过程参见文献[6],综合考虑相关分析和灰色关联分析选择影响日本鲐对马群系资源量的关键环境因子。同理,计算PDO与SSTA月平均值,然后按滞后年份为0 a,1 a,2 a 来分别计算其与资源量的关联度。另外,计算PDO的年平均值,分别将PDO为正位相和为负位相时,它们与对应年份的资源量和渔获量的变化趋势图采用均值化处理。用同样的方法画出厄尔尼诺年和拉尼娜年的SSTA值与对应年份资源量和渔获量的变化趋势图。根据其计算结果来确定是否取PDO和SSTA作为建立模型的因子。
1.3 模型建立
根据选取的关键环境与气候因子依次建立GM(1,2),GM(1,3),GM(1,4)模型。GM(1,N)模型的建模过程可参照参考文献[6]。
2 研究结果
2.1 海表面温度
灰色关联分析(表1)表明:产卵场2 的4 月和9 月海表面温度(SSTSG2-04、SSTSG2-09)与资源量的灰色关联度最大,均为0.77(表1);索饵场4 月的海表面温度(SSTFG-04)与资源量的灰色关联度最大,为0.73。产卵场2 的4 月海表面温度与资源量的相关系数最大(R=0.45,P<0.05,表2);索饵场4 月的海表面温度与资源量的相关系数最大(R= 0.46,P<0.05)。因此,选择SSTSG2-04,SSTSG2-09和SSTFG-04作为影响日本鲐对马群系资源丰度的关键环境因子及其基础数据(表3)。
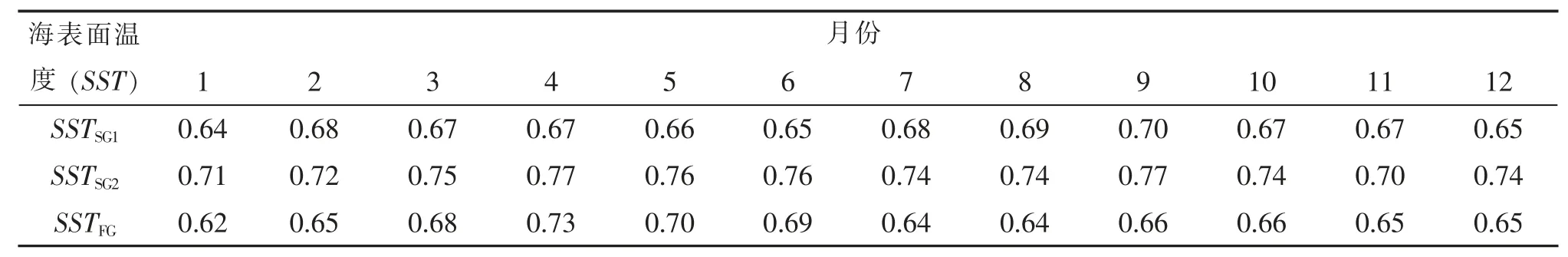
表1 产卵场与索饵场各月海表温度与资源量的关联度分析

表2 产卵场与索饵场各月海表温度与资源量的相关性分析
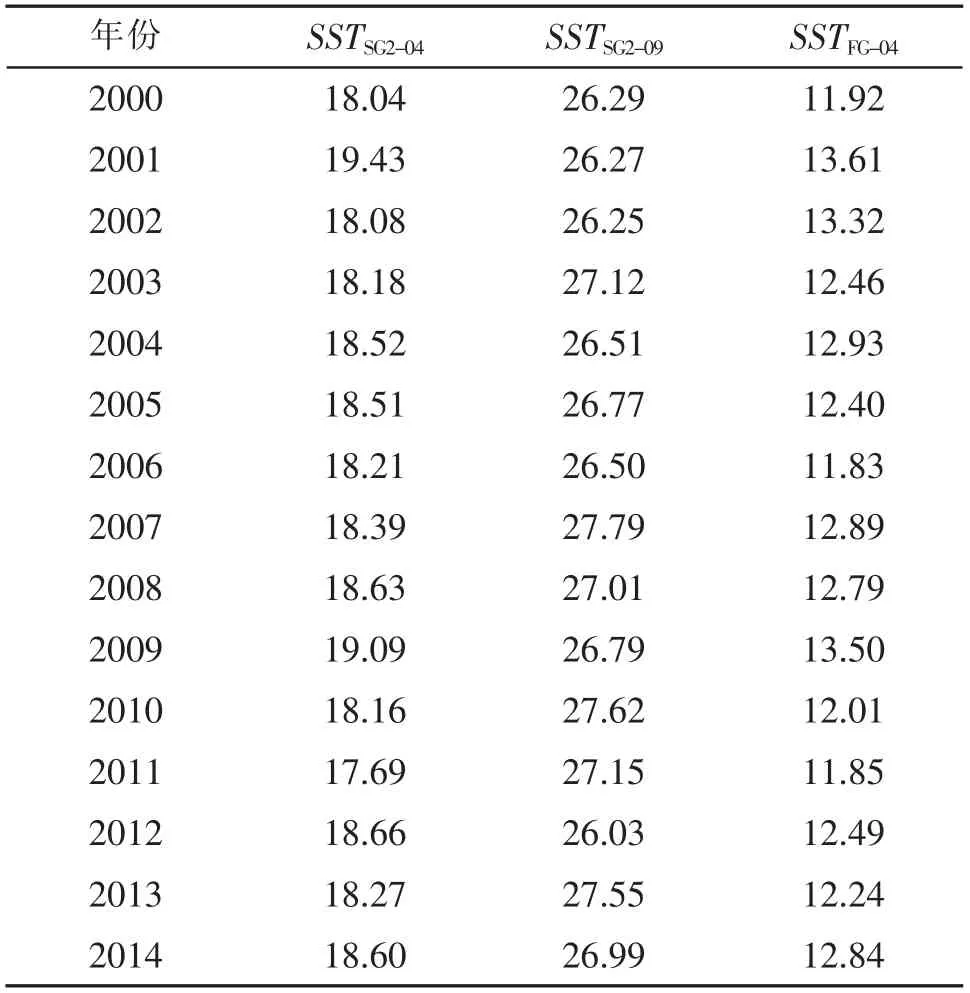
表3 2000-2014 年间SST SG2-04,SST SG2-09 和SST FG-04 的基础数据
2.2 关键气候因子
1973-1987 年海表温距平值波动较大,其中1987 年的距平值为1973 年的2.5 倍,1976 年变化趋势为0.1,资源量变化趋势在1 附近波动,变化不明显;1990 年资源量变化趋势为0.13,距平值变化趋势为0.61;1991-1994 年距平值在0.5~1.5 倍波动,资源量则不断增加;2002-2006 年距平值变化趋势在0~1 间波动,资源量则基本保持在0.5(图3~图4)。1995-1997 年PDO扩大2 倍,资源量下降至原来的0.6;1997 年之后,PDO变化趋势为0.1~2.23,资源量则基本保持在0.5(图5~图6)。综合这些年份的数据发现,PDO和SSTA值的变化对鲐鱼资源量变动的影响不明显。
根据PDO和SSTA的滞后计算灰色关联分析结果可得:L(PDO滞后2 a 的2 月)=0.511 7;L(PDO滞后2 a 的3 月)=0.513 73;L(SSTA滞后1 a的12 月)=0.510 25;L(SSTA当年的1 月)=0.510 94;L(SSTSG2-04)=0.77;L(SSTSG2-09)=0.77;L(SSTFG-04)=0.73。因此,根据文献[11]选择L>0.6 的产卵场2 的4月、9 月海表面温度、索饵场4 月的海表面温度用于资源预测建模。

图3 1973-2014 年间厄尔尼诺年资源量、渔获量与SSTA 的变化趋势图
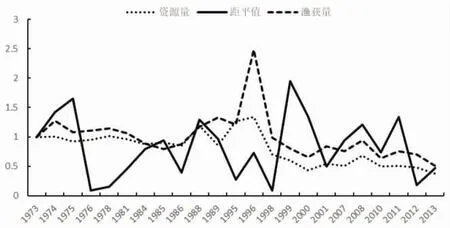
图4 1973-2014 年间拉尼娜年资源量、渔获量与SSTA 的变化趋势图
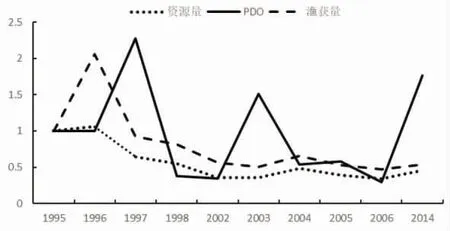
图5 1995-2014 年间暖年资源量、渔获量与PDO 的变化趋势图
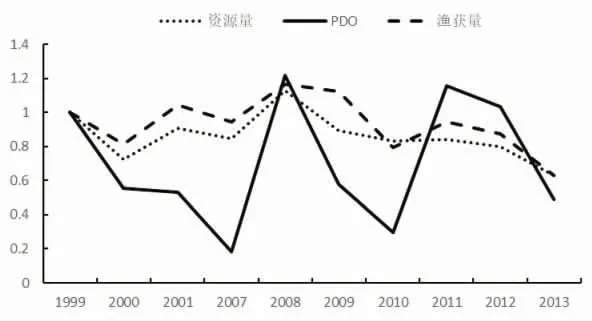
图6 1995-2014 年间冷年资源量、渔获量与PDO 的变化趋势图
2.3 灰色模型预测资源量
根据2.2 节所选出的因子来建立模型(表4)。

表4 根据环境因子所建立的模型
各模型的2000-2014 年的资源量预测结果如表5 所示:模型1 相对残差Q检验结果最小为0.131,模型5 相对残差Q检验结果最大为0.177。包含一个环境因子的GM(1,2)模型,其相对残差Q检验结果均在0.13~0.14 之间。包含产卵场9 月海表面温度的GM(1,N) 模型的相对残差Q检验结果较大。2000-2014 年间SSTSG2-04变化与资源量实际值、预测值的关系详见图7。

表5 各模型的预测结果
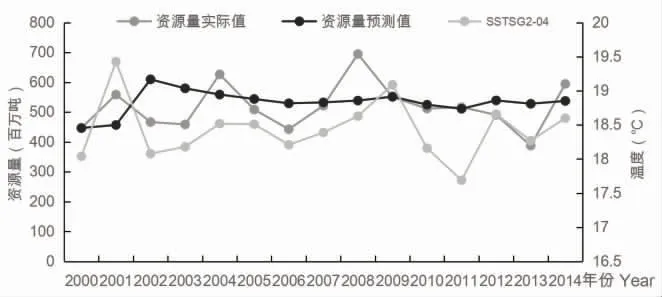
图7 2000-2014 年间SST SG2-04 变化与资源量实际值、预测值关系图
3 讨论与分析
3.1 日本鲐对马群系历年资源量和渔获量变化
对马海域为日本鲐的主要产区,中国、韩国和日本的渔获量占绝对优势。日本鲐对马群系的资源量和渔获量在1987-2001 年间波动较大(图1),1996 年均达到峰值,其后资源量波动变化但总体上低于1996 年以前的水平,渔获量则稍低于达到峰波动前的水平,趋于稳定。从1996 年开始,资源量的总体下降趋势与亲鱼数量的减少有关,自1900年之后,捕捞日本鲐总数中0 岁和1 岁占比例较大,2 岁以上占比例较小[2],但是鲐鱼1 岁之后才开始产卵,因此降低了种群繁殖能力。1990 年资源量骤减与当时机轮围网渔业规模较大、过度利用资源有关[12]。从1999 年开始,灯光围网渔业的大力发展以及在近海对幼鱼资源的过早捕捞,使其资源水平出现二次衰退的迹象[12]。由于鲐鱼的生长周期不长,并具有生长迅速的特点,所以如保持一定的产卵群体,还是可以保持其产量的[13]。
3.2 灰色预测模型分析
灰色系统模型的检验方法有很多种,常见的有后验差检验、关联度检验、残差检验等,其中残差检验是一种较为客观的检验方法,所以本文所建模型采用残差检验方法。在灰色预测模型中,残差检验的结果小于0.35 即为精度较好的模型。从各个模型的相对残差Q检验结果均小于0.35 来看,GM(1,N)模型是预测其资源量的有效方法,但是这也与建立GM(1,N)模型时所选择的因子有很重要的关系。从预测结果来看,加入SSTSG2-09因子后的模型的相对残差Q检验值比没有加入SSTSG2-09因子的模型的相对残差Q检验值较大,但是只加入SSTSG2-09因子的GM(1,2)模型的相对残差Q检验值较小,原因可能是产卵洄游对于环境的要求更为严格[14]。日本鲐鱼在位于南部的产卵场的产卵时间是1-4 月,9 月南下洄游越冬群体中成鱼较多,温度对成鱼的影响与幼鱼不同[15-17],所以SSTSG2-09对鲐鱼资源丰度的影响低于SSTSG2-04。只加入SSTSG2-04的GM(1,2)模型的残差Q检验结果最小,模型预测数据也最最接近实际情况(图6),原因是SSTSG2-04与资源量有较高的相关性,所以更能反映其资源丰度。此外,产卵场1 的4月海表面温度与资源量的关联度较低于产卵场2(表1),SSTSG2各月的值与资源量的关联度均在0.7~0.77 之间,SSTSG1各月的值与资源量的关联度均在0.6~0.7 之间,原因可能是日本对日本鲐对马群系的资源量评估较低[3],主要侧重于日本海海域,而产卵场1 属于近海的产卵场。3.3 各因子影响分析
日本鲐对马群系产卵季节在1-6 月,产卵场的纬度不同,产卵时间有所不同[18]。本次研究选取了两个产卵场,分为纬度较低的产卵场1(26°~31° N,122°~127° E)与纬度较高的产卵场2(30°~35° N,128°~131° E)。有研究[18]称,日本鲐主要选择短距离洄游,东海、黄海海域与日本海海域都有其对应的产卵场和索饵场。李日嵩等[5]发现,在鲐鱼产卵后的15~40 d(4 月)这个时间段的水温等物理环境是影响其最终补充量的关键动力学因素,该期间仔幼鱼所处的平均水温往往低于最适合水温,也是此阶段的仔幼鱼生存的最佳温度。吕为群等[19]研究发现,鱼类发育时期的温度会影响其成年后的生长及繁殖状况,低温发育会使繁殖力下降,高温发育会使其生长速率降低。产卵场2 的4 月的温度接近仔幼鱼生长最适温度20 ℃,而且日本鲐在这里的产卵时间是1-4 月,用其作为建立GM(1,2)模型的因子结果最优,所以模型因子选择SSTSG2符合实际情况。
4 结论
(1)对日本鲐对马群系的资源量预测结果与事实情况较为接近,这是因为产卵场的选择符合实际情况,而产卵场温度这一环境因子又是直接影响其补充群体正常发育生长的重要环境因素。渔业资源的变动受到资源本身、捕捞行为、环境和气候等多方面的影响[20],日本鲐对马群系在日本岛海域和东海、黄海都有产卵场分布,属于洄游性鱼类,常集群进行繁殖、索饵和越冬洄游[21]。由于日本鲐不同群体进行产卵和索饵的地理位置不同,研究其各个产卵场和索饵场温度变化对于其资源量变动的较大影响,对于其资源量预测有重要意义。
(2)灰色理论和建立的GM(1,N)模型适用于渔业研究。在本文中,将依据灰色关联度和相关分析选取的SSTSG-04,SSTSG-09和SSTFG-04作为建立日本鲐对马群系资源量预测模型的因子。根据模型分析结果得出,基于SSTSG-04所建立的GM(1,2)模型是最优模型。
(3)本文虽然分析了SSTA 和PDO 与鲐鱼资源量的关系,但在建模过程中并没有涉及,主要是因为冷、暖年和不同强度的厄尔尼诺、拉尼娜事件对鲐鱼资源的影响不同[22-24]。在以后的研究中,应分别研究其与资源量的具体对应关系,利用更多的多元统计法[25]建立更精确的预测模型,为日本鲐鱼的资源预测和科学管理提供更好的理论依据。
猜你喜欢
当代水产(2022年8期)2022-09-20
成都信息工程大学学报(2022年2期)2022-06-14
网络安全与数据管理(2022年3期)2022-05-23
矿产勘查(2020年11期)2020-12-25
矿产勘查(2020年7期)2020-12-25
北京航空航天大学学报(2020年10期)2020-11-14
青年文学家(2020年16期)2020-07-13
北京航空航天大学学报(2019年9期)2019-10-26
文学港(2019年5期)2019-05-24
