
广玉兰新品种‘碧翠’的抗寒性评价及抗寒性指标筛选
2020-04-10 12:32:12孙凌霄金晓玲胡希军谭殷殷罗峰张哲陈燕
湖南生态科学学报 2020年1期
孙凌霄, 金晓玲,2*, 胡希军,2, 谭殷殷, 罗峰, 张哲, 陈燕
(1.中南林业科技大学 风景园林学院,湖南 长沙 410004;2.湖南省环境资源植物开发与利用工程技术研究中心,湖南 长沙 410004)
广玉兰(Magnoliagrandiflora)又名荷花玉兰、洋玉兰,为木兰科(Magnoliaceae)木兰属(Magnolia)常绿乔木,树高7 m~13 m,在原产地最高可达30 m,胸径20 cm~39 cm;树皮深灰色,嫩枝和芽均密被锈色短绒毛,具有很大的向北方地区引种的潜力[1].
木兰科植物的抗寒性研究一直以来都是一个比较热门的研究方向,大量试验研究都认为广玉兰是木兰科中抗寒性较强的植物[2].刘艳萍等[3]通过测定几种广玉兰品种的相对电导率计算出半致死温度,并进行比较.相对电导率随着温度的降低,先缓慢上升,然后急剧升高,最后趋于稳定,每个品种的变化趋势均一致.开始降温时相对电导率缓慢增加,说明在此温度范围内能够适应,但温度下降到-5 ℃~-10 ℃时,相对电导率开始急剧升高,而且温度进一步降低到-20 ℃~-25 ℃时,广玉兰叶片的相对电导率趋于稳定,说明此时细胞已受到严重伤害.几个广玉兰品种的半致死温度最高为-18.86 ℃,最低可达-22.56 ℃.李杰等[4]研究了低温胁迫对广玉兰生理特性的影响,在人工低温胁迫条件下,探讨广玉兰细胞质膜相对透性[5]、SOD活性[6]和可溶性糖含量[7]动态变化规律得出结论:广玉兰细胞质膜相对透性随温度下降而呈“S”型曲线变化,与温度成负相关,不同温度处理下SOD活性变化呈双峰曲线,且与温度呈正相关;可溶性糖含量呈先升高后下降的变化趋势.
对广玉兰的杂交后代和变种的研究较少,本研究以4年生新品种‘碧翠’和4年生‘埃克斯茅斯’(园林绿化中普遍使用的广玉兰品种)为试验材料,研究在人工低温胁迫过程中植株的抗寒性,旨在了解该品种的抗寒性强弱,便于进一步推广,同时筛选广玉兰的抗寒性评价指标,为广玉兰在园林中引种驯化和推广提供依据.
1 材料与方法
1.1 试验材料
试验材料为种植于中南林业科技大学校园内的四年生‘埃克斯茅斯’广玉兰(Magnoliagrandiflora‘Exmouth’)和四年生‘碧翠’广玉兰(Magnoliagrandiflora‘Bicui’),选择长势和管理良好的植株进行试验.
‘埃克斯茅斯’广玉兰叶片深绿色,叶形较窄,叶背面有棕色绒毛.‘碧翠’广玉兰叶片颜色翠绿,叶形较宽,叶背面无毛.
1.2 试验方法
在2017年2月23日进行采样.从两植株上随机采集位于中上部的当年生侧枝上的第3枚~5枚完整无破损大小相近的成熟叶片,每株采8片(2片备用),采摘后立即放入密封塑料袋带回实验室.将叶片用蒸馏水洗净再用去离子水清洗,用滤纸擦干净后分别取两棵植株的6片叶片,放入6个洁净的密封袋中,共12片叶片.把装袋的叶片放置低温恒温槽中,试验设6个温度梯度,分别为:5 ℃(常温)、0 ℃、-5 ℃、-10 ℃、-15 ℃、-20 ℃,每个梯度均处理2 h,之后将材料取出放入冰箱解冻,次日测定各材料的生理生化指标,将每组平均分成3份,重复3次,测得结果求平均值.
采用电导仪测定法测定相对电导率,仪器为雷磁DJS-1D电导仪;采用酸性茚三酮法进行游离脯氨酸含量测定;采用硫代巴比妥酸比色法进行丙二醛含量测定;采用考马斯亮蓝G-250染色法进行可溶性蛋白含量测定;采用氮蓝四唑(NBT)光化还原法测定SOD活性;采用愈创木酚法测定POD活性.
1.3 数据处理
数据采用Excel 2010及SPSS 18.0软件处理,对各项指标进行方差分析和相关性分析,采用Logistic拟合求得半致死温度,应用模糊隶属函数对‘碧翠’广玉兰和‘埃克斯茅斯’广玉兰进行抗寒性综合评价.将每个品种各阶段各指标的隶属函数值相加求平均值,隶属函数均值越大的品种抗寒性越强.
2 结果与分析
2.1 低温胁迫对相对电导率的影响
叶片受低温胁迫,由于细胞膜受到破坏致使胞内物质外渗,温度的降低会使外渗物质增多,相对电导率(REC)便会逐渐增大,随着人工低温胁迫的加强,两个广玉兰品种的离体叶片REC不断增大,与低温胁迫程度呈负相关.在不同的温度段,REC增加的程度不同,从两个品种的REC变化曲线可以看出,当温度不断降低达到临界值时,REC的变化趋势由缓慢上升变为急剧上升,不同品种的这个临界值会有所不同,由图1可知,‘碧翠’的临界值在0 ℃~-5 ℃之间,‘埃克斯茅斯’广玉兰的临界值在-5 ℃~-10 ℃之间,‘埃克斯茅斯’广玉兰细胞膜通透性发生大幅度变化的临界温度低于‘碧翠’广玉兰.

图1 低温胁迫对相对电导率的影响Fig.1 Effect of low temperature stress on REC
两个广玉兰品种的的REC拟合的Logistic方程,均达到极显著水平.得到‘埃克斯茅斯’广玉兰的半致死温度为-4.28 ℃,‘碧翠’广玉兰的半致死温度为-3.03 ℃(表1).

表1 相对电导率的Logistic方程参数值及半致死
e自然对数的底数约等于2.71828
2.2 低温胁迫对MDA含量影响
由图2可知,低温胁迫下,两个品种MDA含量变化趋势相同,均随温度下降而增加,与低温胁迫呈极显著负相关.‘埃克斯茅斯’广玉兰的MDA含量高于‘碧翠’广玉兰.两者在-10 ℃~-15 ℃之间增加较多,在-20 ℃时MDA含量达到最大值,‘埃克斯茅斯’广玉兰为24.63 umol/g FW,‘碧翠’广玉兰为22.83 umol/g FW.
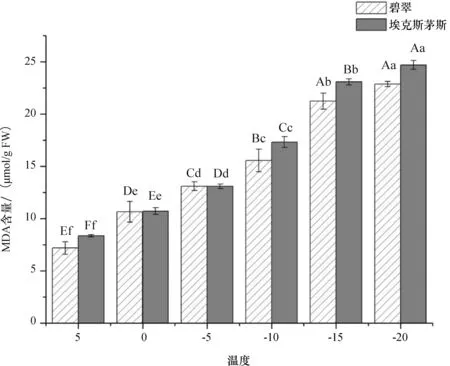
图2 低温胁迫对MDA含量的影响Fig.2 Effect of low temperature stress on MDA content
2.3 低温胁迫对游离脯氨酸含量影响
通常植物体内游离脯氨酸(Pro)含量不高,当植物受到胁迫时,Pro含量会迅速增加,该指标可以反映植物受低温伤害程度及对低温的承受能力.从图3可以看出,两个广玉兰品种的Pro含量随着温度的降低呈先升后降的趋势,但积累的量与积累速率有差异.不同温度下两个广玉兰品种的Pro含量存在明显差异,‘埃克斯茅斯’广玉兰明显高于‘碧翠’广玉兰.两种广玉兰的Pro含量均在-5 ℃时达到最大值,‘埃克斯茅斯’广玉兰为152.90 ug/g FW,‘碧翠’广玉兰为150.29 ug/g FW.
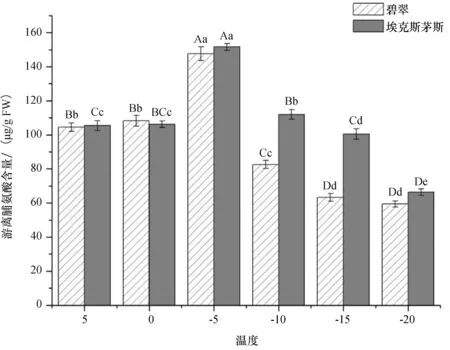
图3 低温胁迫对游离脯氨酸含量的影响Fig.3 Effect of low temperature stress on free proline content
2.4 低温胁迫对可溶性蛋白含量影响
可溶性蛋白(SP)具有较强的亲水性,能够增强细胞的保水能力,从而提高植物的抗寒性.由图4可知,随着温度的降低,两个广玉兰品种的SP含量均呈现先升后降的趋势,SP含量整体与低温胁迫温度呈极显著正相关,均在-5 ℃达到最大值,且在温度低于-5 ℃时两个品种的SP含量差异显著,‘碧翠’广玉兰的下降幅度和速率大于‘埃克斯茅斯’广玉兰.因此,在低温环境下‘埃克斯茅斯’广玉兰的SP含量高于‘碧翠’广玉兰,其保水能力及抗寒性更强.
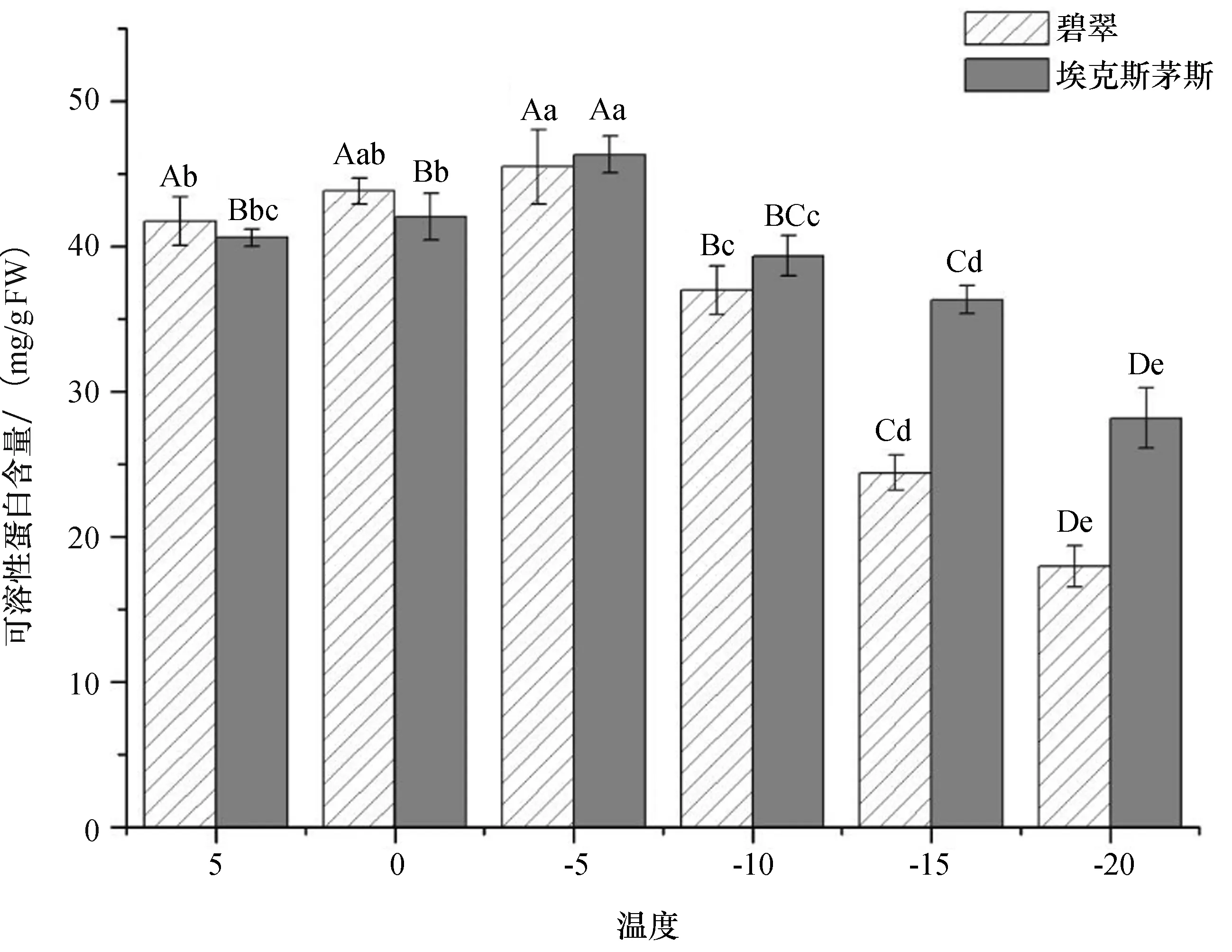
图4 低温胁迫对可溶性蛋白含量的影响Fig.4 Effect of low temperature stress on soluble protein content
2.5 低温胁迫对POD、SOD活性影响
在植物受到低温胁迫时,体内的活性氧和自由基会积累,会导致细胞结构和功能的破坏,而POD和SOD可以有效清除细胞内的这些有害物质,避免植物受到伤害.由图5可知,随着温度降低,两个广玉兰品种的SOD活性差异较大,变化趋势基本相同,但酶活性达到最大值的温度不同.随着温度降低‘埃克斯茅斯’广玉兰的SOD活性逐渐增大,-15 ℃时达到最大值;随着温度降低,‘碧翠’广玉兰的SOD活性逐渐增大,-5 ℃时达到最大值.当温度在一定范围内小幅度降低时,植物的抗逆性使SOD活性增强,以清除细胞内的活性氧和自由基,而过低的温度会使酶活性降低.
由图6可知,温度降低对两种广玉兰POD活性影响不一致.温度降低‘埃克斯茅斯’广玉兰POD活性先降低,-5 ℃时达到最小值,然后增大,-15 ℃达到最大值;温度降低‘碧翠’广玉兰POD活性先升高,-5 ℃时达到最大值,两个品种无明显变化规律.
2.6 两个广玉兰品种抗寒性综合评价
植物的抗寒机制十分复杂,只用某个指标判断抗寒性具有片面性,不同的抗寒指标在不同植物上的响应程度也会有一定差异,因此应对多个抗寒指标进行综合分析才能全面地反映抗寒性.采用模糊数学的隶属函数进行转换,将两个品种各梯度的隶属函数值相加求均值,均值越大,品种的抗寒性就越强[8-10].由表2可知,‘埃克斯茅斯’广玉兰各隶属函数均值大于‘碧翠’广玉兰,‘埃克斯茅斯’广玉兰抗寒性大于‘碧翠’广玉兰.

图5 低温胁迫对SOD活性的影响Fig.5 Effect of low temperature stress on SOD activity
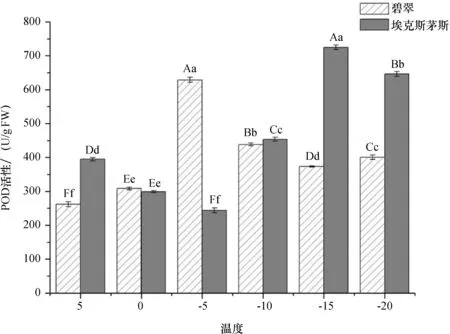
图6 低温胁迫对POD活性的影响Fig.6 Effect of low temperature stress on POD activity

表2 两个广玉兰品种抗寒性评价
3 讨论
由于自然环境中有极端天气等不可控因素,同一物种栽培于不同地理区域时,其低温半致死温度也会有所改变.同时,不同试验条件下测出的低温半致死温度也有一定差别.所以,低温半致死温度应是一个相对的概念,它只能够用于衡量几个种之间的相对抗寒性强弱,而不能直接用来指导植物栽培.
陈洁等[11]对几种含笑属植物抗寒生理指标筛选进行研究,认为REC、MDA和SP等三个指标能够反映出含笑属植物抗寒性.本研究认为广玉兰的抗寒生理指标是REC和SP,相比之下少了MDA指标.含笑属抗寒指标筛选研究发现,低温胁迫下MDA含量与抗寒性呈正相关[11],但本研究认为低温胁迫下两个广玉兰品种的MDA含量的增加无明显差别,因此,MDA不能作为广玉兰品种抗寒性的生理指标.MDA是由于植物细胞受到伤害后应激反应的产物,伤害越大MDA含量越高,MDA含量越高植物抗寒性越弱[8,9].但有些研究结果却相反,MDA含量与其抗寒性强弱呈正比,本研究的结果也是MDA含量高的品种抗寒性更强.MDA含量作为植物抗寒性生理指标以及对木兰科植物的抗寒性作用机理还有待进一步研究.
4 结论
两个广玉兰品种的抗寒能力存在差异,‘埃克斯茅斯’广玉兰半致死温度低于‘碧翠’广玉兰.对6个生理生化指标进行综合评价,REC、SP两个生理生化指标是反应抗寒性强弱的较佳指标.
猜你喜欢
作文与考试·小学高年级版(2023年8期)2023-07-17 09:32:52
真空与低温(2022年6期)2023-01-06 07:33:20
现代塑料加工应用(2021年5期)2021-02-28 08:18:04
中国诗歌(2018年5期)2018-11-14 20:52:30
作文评点报·低幼版(2018年30期)2018-09-27 12:21:24
作文评点报·低幼版(2018年30期)2018-09-27 12:21:24
劳动保护(2018年8期)2018-09-12 01:16:18
新疆农垦科技(2016年2期)2016-08-21 13:50:26
中国果菜(2016年9期)2016-03-01 01:28:40
中国马铃薯(2015年5期)2016-01-09 06:11:11
