
合江方竹根际土壤丛枝菌根真菌多样性1)
2020-04-10 05:55:16姚莉梅尚昆朱青青江龙
东北林业大学学报 2020年3期
姚莉梅 尚昆 朱青青 江龙
(贵州大学,贵阳,550025)
合江方竹(ChimonobambusahejiangensisC.D.Chu et C.S.Chao)是禾本科竹亚科寒竹属植物,是中国的特有种,主要的分布区包括四川合江、贵州赤水和贵州习水等地,生长海拔为800~1200m。因其出笋期在出笋淡季,且自身营养比较丰富,通过加工成鲜笋或干笋的形式进行销售,深受市场欢迎,开发价值很高[1]。丛枝菌根(AM)真菌是球囊菌门(Glomeromycota)的一类专性活体营养共生菌[2],80%以上的植物都能被AM真菌侵染形成共生体[3],通过共生体AM真菌能促进植物对N、P、K等矿质元素及微量元素的吸收和利用,增强植物的抗旱、抗寒和盐胁迫等抗逆性,促进植物生长,加快幼苗成活,促进农林业发展。研究发现,竹类与AM真菌也存在共生关系,可以在土壤中形成共生体以促进竹类植物的生长[4],但在方竹方面相关研究较少,仅见叶文兰等[5]报道了金佛山方竹(Chimonobabusautilis)根围AM真菌种类,关于合江方竹共生AM真菌资源研究未见报道。本研究以合江方竹根际土壤为研究对象,对合江方竹根系进行侵染调查研究,并从根际土壤中分离AM真菌孢子,通过形态学特征对合江方竹AM真菌进行种属鉴定,探明与合江方竹共生的AM真菌种类及多样性状况,为AM真菌在合江方竹上的应用,特别是菌根苗生产中的菌种筛选提供理论依据。
1 材料与方法
土样采集:采样地位于贵州省遵义市赤水市大同镇合江方竹天然林分布区,海拔831~1 050 m。选取长势良好的合江方竹,采集根系和根际土壤。采样时,挖除土壤表面约2 cm的土壤及杂质,采集距地表5~30 cm土层中合江方竹根系和根际土壤,3株重复,混合后约1 kg,放入采集袋中,待根际土壤自然风干后,放入冰箱中4 ℃进行保存,备用。用自来水将根系表面土壤杂质洗净后,放进广口瓶,用FAA固定液固定备用[6]。
根系侵染状况与侵染率:采用碱解离-酸性品红染色法观察根系菌根侵染状况[7]。从FAA固定液中取出根系,清水冲洗2~3次后将根系裁剪为1 cm的小段,用质量分数10% NaOH溶液在90 ℃水浴锅中解离1 min,水洗3~5次,用30% H2O2在90 ℃水浴锅中酸化0.5~1.0 min,水洗3~5次。根段用酸性品红乳酸甘油染色2 d,取出进行制片,并置于Olympus BX53光学显微镜下观察,记录每条根段各种菌根结构(菌丝、泡囊和根内孢子等)的侵染情况,根据每条被侵染菌根的根段长度所占根段总长度的百分比,按加权法[8]计算根段侵染率,共统计50根,3次重复。
侵染率=∑((0×未侵染的根段数)+(10%×侵染率为10%的根段数)+(20%×侵染率为20%的根段数)+…+(100%×侵染率为100%的根段数))/观察的总根段数。
AM真菌的分离与鉴定:采用湿筛倾注-蔗糖离心法[9]对土壤AM真菌孢子进行分离。将分离后的孢子置于体式显微镜下挑选成熟孢子,用无菌枪头吸取孢子于载玻片上,以水为载浮剂,置于生物显微镜(Olympus BX53)和摄影系统(Olympus DP70)下,对孢子大小、形状、颜色、连孢菌丝、孢子壁及发芽壁的层数和厚度、孢子压破后的内含物及在Melzer’s试剂中的染色情况、孢子表面有无纹饰或附着物等特征进行详细观察、记录,并照相[10]。依据国际丛枝菌根真菌保藏中心(INVAM,http://invam.caf.wvu.edu)、http://www.amf-phylogeny.com和http://www.zor.zut.edu.pl/Glomeromycota等网站提供的种属描述及其相应的图片,结合贵州大学AM真菌保藏资源进行种属鉴定,中文学名参见王幼珊等[11]球囊菌门丛枝菌根真菌最新分类系统菌种名录。
物种多样性分析:按上述孢子分离方法,每个土样重复3次,测定以下指标,孢子密度(SD)为20 g风干土样中孢子的总数;相对多度(RA)为该采样点某种孢子数占该采样点总孢子数的比例;相对频度(RF)=(某个种的频度/所有种的频度总和)×100%;重要值I=(RF+RA)/2,即频度和相对多度的平均值。
2 结果与分析
2.1 合江方竹根系侵染状况
镜检观察合江方竹根系的AM真菌侵染情况(图1),其菌根侵染率为23%,其中菌丝和泡囊是AM真菌侵染根系的主要形式(图1A、B),此外也可观察到根系表面存在根内孢子(图1C),说明AM真菌可侵染合江方竹根系,能与之建立起良好的共生关系。

V.泡囊;H.菌丝;S.根内孢子。
2.2 合江方竹根际AM真菌的鉴定
经过孢子形态学的鉴定,从贵州省赤水市合江方竹根际土壤中共分离、鉴定出AM真菌8属24种,其中无梗囊霉属(Acaulospora)10种,多样孢囊霉属(Diversispora)1种,盾巨孢囊霉属(Scutellospora)1种,巨孢囊霉属(Gigaspora)1种,球囊霉属(Glomus)6种,隔球囊霉属(Septoglomus)2种,硬囊霉属(Sclerocystis)1种,双型囊霉属(Ambispora)2种,详见图2。
2.3 AM真菌多样性
由表1可知,球囊霉属和无梗囊霉属为合江方竹根系AM真菌优势属,柯氏无梗囊霉(A.koskei)、小果球囊霉(G.microcarpum)、长孢球囊霉(G.dolichosporum)为优势种。
3 结论与讨论
笔者在染色的合江方竹根系中观察到根内孢子、菌丝和泡囊等结构,说明AM真菌能够侵染合江方竹根系,其侵染率为23%,表明AM真菌能与合江方竹建立较好的共生关系。该研究结果与叶文兰等[7]发表的金佛山方竹侵染率相差不大,但是与其他植物如甘蔗(Saccharumofficinarum)[29]、玉米(Zeamays)[30]、栌橘木(Noueliainsignis)[31]、胡杨(Populuseuphratica)和骆驼刺(Alhagisparsifolia)[32]相比,侵染率并不高。不同植物之间侵染率有一定的差异性,研究表明,土壤类型[33]、质地、土壤中的养分情况[34]、土壤pH[35]、温度[36]及湿度[37]都会对菌根侵染率造成影响,除此之外植物与AM真菌的亲和程度也造成侵染率差异[38]。合江方竹侵染率低可能是因为采样时间为冬季,温度低,且生长环境潮湿,也可能和调查地土壤中养分情况,尤其是含磷水平有关,具体原因有待进一步研究。虽然合江方竹自然状态下侵染率不高,但是可以通过人为接种AM真菌菌种来提高其侵染率,如邢红爽等[39]通过接种摩西斗管囊霉(Funneliformismosseae)和变形球囊霉(Glomusversiforme)提高了百合(Liliumbrowniivar.viridulum)的侵染率。因此,通过接种AM真菌在提高合江方竹产量、品质、移栽苗成活率等应用上仍然具有广阔的前景。


GW1、GW2、GW1-L1、GW1-L2、GW2-L1、GW2-L2.发芽壁壁层;L1、L2、L3.孢子壁壁层;1-2.细凹无梗囊霉(Acaulosporascrobiculata)[12];3-4.蜜色无梗囊霉(A.mellea)[13];5-6.柯氏无梗囊霉(A.koskei)[9];7-8.皱襞无梗囊霉(A.rugose)[14]19;9-10.波状无梗囊霉(A.undulata)[15-16];11-12.光壁无梗囊霉(A.laevis)[14]14;13-14.孔窝无梗囊霉(A.foveata)[17];15-16.凹坑无梗囊霉(A.excavate)[14]11;17-18.疣状无梗囊霉(A.tuberculata)[14]23-24;19-20.大型无梗囊霉(A.colossica)[14]7-8;21-22.光亮多样孢囊霉(Diversisporaclara)[18];23-24.美丽盾巨孢囊霉(Scutellosporacalospora)[19];25-26.球状巨孢囊霉(Gigasporamargarita)[20];27-28.长孢球囊霉(Glomusdolichosporum)[21];29-30.小果球囊霉(G.microcarpum)[14]70-71;31-32.刺球囊霉(G.spinosum)[22];33-34.黄孢球囊霉(G.flavisporum)[23];35-36.卷曲球囊霉(G.convolutum)[24];37-38.大果球囊霉(G.macrocarpum)[23];39-40.叉状隔球囊霉(Septoglomusfurcatum)[25-26];41-42.缩隔球囊霉(S.constrictum)[14]44-45;43-44.悬钩子硬囊霉(Sclerocystisrubiforme)[27];45-46.厚皮双型囊霉(Ambisporacallosa)[28];47-48.网纹双型囊霉(A.reticulata)[16]。
图2合江方竹根系AM真菌孢子形态
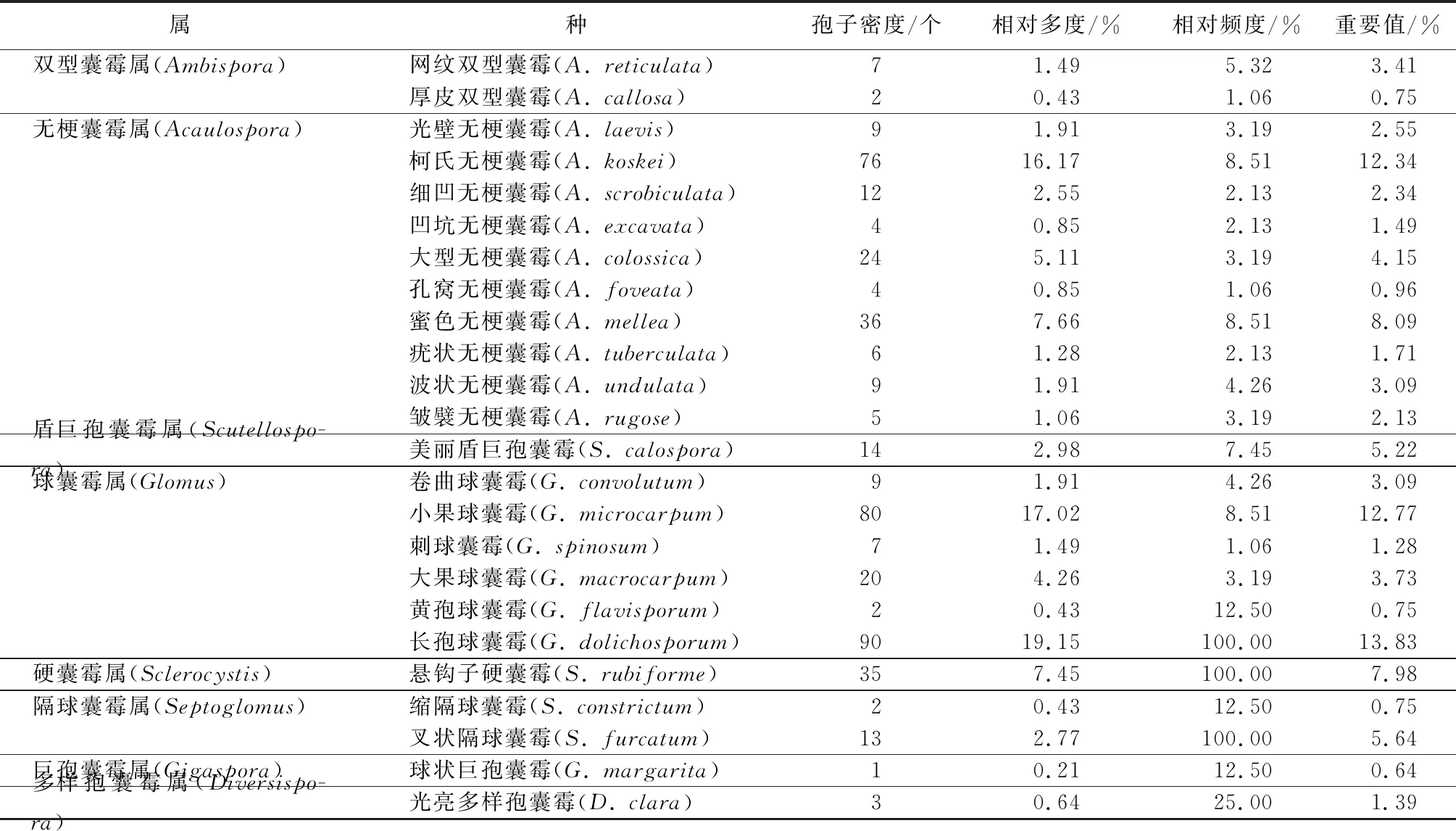
表1 合江方竹根系AM真菌多样性指标
合江方竹天然林分布地贵州省遵义市赤水市大同镇方竹根际土壤中AM真菌孢子经形态学鉴定确定其有8属24种,其中无梗囊霉属(Acaulospora)10种,多样孢囊霉属(Diversispora)1种,盾巨孢囊霉属(Scutellospora)1种,巨孢囊霉属(Gigaspora)1种,球囊霉属(Glomus)6种,隔球囊霉属(Septoglomus)2种,硬囊霉属(Sclerocystis)1种,双型囊霉属(Ambispora)2种。可见,在自然环境中,合江方竹根系AM真菌种质资源丰富。其中无梗囊霉属和球囊霉属的孢子种类和数量较多,与金佛山方竹[5]、紫背天葵(Begoniafimbristipula)[40]、半夏(Pinelliaternata)[41]、滑桃木(Trewianudiflora)[42]、广西木薯(Manihotesculenta)[43]、广西柳江生姜(Zingiberofficinale)[44]、牡丹(Paeoniasuffruticosa)主栽培根际根围[45]和贵州茶树(Camelliasinensis)[46]的AM真菌组成具有一定的相似性,说明无梗囊霉属与球囊霉属能适应更多的环境,具有适应性强、分布较广泛的特点。本研究中的AM真菌物种多度与金佛山方竹相似,但属的多样性高于金佛山方竹,可能是合江方竹生存的环境更适合与AM真菌不同属建立共生关系,也可能是合江方竹与AM真菌具有更好的亲和性,具体原因有待进一步研究。
AM真菌的多样性受到环境因素影响,其对环境的适应也具有选择性,适应生境的菌种大量繁殖并随之成为优势种,相反则会被慢慢淘汰至消失在该类生境中[47]。本研究中的柯氏无梗囊霉(A.koskei)、小果球囊霉(G.microcarpum)、长孢球囊霉(G.dolichosporum)可能自身生物学的特性比较适应该生境,在试验观察过程中出现的频度较高、重要值高,初步确定为该生境优势种,与金佛山方竹[5]、灌木铁线莲(Clematisfruticosa)[48]及梵净山菝葜(Smilaxchina)、箭竹(Fargesiaspathacea)和银叶杜鹃(Rhododendronargyrophyllum)等植物根际土壤丛枝菌根的研究结果[19]基本一致。
完善植物丛枝菌根真菌的种质资源调查,对筛选高效菌种、菌根真菌资源在农林业上的开发和应用具有重要的意义,如英国洛桑农业研究中心将生产的AM菌剂推广到法国、丹麦和日本等国家,将其广泛运用在果树、蔬菜及花卉上,提高了经济效益[49]。在生物技术上,通过接种丛枝菌根真菌,育成菌根化苗木可促进幼苗生长[50-52]、提高植物的抗旱性[49]、提高植物耐受盐碱胁迫能力[53]、增强对N、P等元素的吸收[54]能力。本研究在国内首次对合江方竹根际中AM真菌的侵染状况进行了调查,并对根际AM真菌种类进行了鉴定,确定了合江方竹部分AM真菌种质资源,完善了贵州部分丛枝菌根真菌(AMF)菌种资源,为丛枝菌根的研究与应用提供了资源;在生产上,可为菌剂筛选提供基础数据,为合江方竹菌根化育苗、栽培和丰产提供依据。
猜你喜欢
农村实用技术(2022年2期)2022-11-22 07:27:43
岭南音乐(2022年6期)2022-02-04 13:50:24
现代园艺(2020年13期)2020-12-21 05:44:32
防护林科技(2020年10期)2020-12-20 23:26:27
园林科技(2020年2期)2020-01-18 03:28:26
——我可爱的家乡
黄河之声(2017年20期)2017-12-27 15:51:59
岷峨诗稿(2017年4期)2017-04-20 06:26:36
小学教学研究·新小读者(2014年12期)2014-12-03 13:14:06
环境与可持续发展(2013年6期)2013-03-11 16:21:51
植物营养与肥料学报(2011年6期)2011-10-24 05:53:04
