
大金山岛常绿阔叶林和落叶阔叶林中小型土壤动物群落特征
2020-04-03 07:44罗鼎晖骆蓓菁许洺山阎恩荣由文辉
生态与农村环境学报 2020年3期
罗鼎晖,李 翔,骆蓓菁,许洺山,妥 彬,阎恩荣,由文辉
(浙江普陀山森林生态系统定位观测研究站/ 华东师范大学生态与环境科学学院,上海 200241)
土壤动物一般指生活阶段大部分或全部在土体内或土表凋落物层度过的动物[1]。目前的研究一般根据体宽将土壤动物分成小型(平均体宽小于0.1或0.2 mm,例如线虫和轮虫等)、中型〔平均体宽在0.1(或0.2)~2 mm之间,例如螨虫和跳虫等〕和大型(平均体宽在2 mm以上,例如蚯蚓和马陆等)[2]。相较于大型土壤动物,中小型土壤动物由于体积较小,挖掘能力较弱,其适应生存环境水热条件改变的能力较差,因此其群落特征对于季节变化的响应较为明显[3]。此外,中小型土壤动物对土壤物理结构的改变较小,因此主要通过取食作用影响土壤物质和能量循环,例如:土壤线虫可以通过取食微生物显著改变土壤碳、氮周转[4]。“金山三岛海洋生态自然保护区”是上海市所辖范围内第一个自然保护区[5],大金山岛作为其中最大的岛屿,其上自然植被保护良好,甚少有人类活动,土壤动物的生存环境近似自然水平。目前国外大部分研究主要集中于海岛土壤动物分类学相关内容的探索[6],鲜有涉及海岛土壤动物群落多样性的研究[7]。国内对于海岛土壤动物群落多样性研究同样较为匮乏[8]。基于此,笔者分秋、冬、春和夏4季调查分析了大金山岛常绿阔叶林和落叶阔叶林中小型土壤动物密度、类群数、垂直分布及群落多样性指标,解析土壤理化特性与中小型土壤动物群落特征参数的关系,旨在探究岛屿森林植被对中小型土壤动物群落特征的影响规律及其机制,为大金山岛森林生态系统的保护提供基础理论依据。
1 研究区域与研究方法
1.1 研究区概况
大金山岛(30°41′42″ N,121°24′25″ E)地处上海西南角的杭州湾,位于中亚热带北缘,受东南季风及太平洋暖湿气团的影响,属于海洋性气候区,温和湿润,四季分明。金山区气象资料显示,大金山岛年均温度为16.9 ℃,平均最高温度为37.2 ℃,平均最低温度为-5.1 ℃,年均总降水量约为1 000 mm,年日照时间为2 000 h[9]。大金山岛植被在外貌水平上可划分为草甸、灌草丛、灌木林、常绿阔叶林和落叶阔叶林5个类型[10]。通过前期调查,典型和代表性的大金山岛森林植物群落包括常绿阔叶林中青冈、天竺桂和红楠群落以及落叶阔叶林中麻栎、野桐、椿叶花椒、朴树和丝绵木群落8种类型。
1.2 样品采集及处理
在8种森林植物群落类型样地选址(图1)完成后,对每个群落的基本群落学特征进行调查,调查内容包括群落内乔木、灌木和草本层植物名称,建群种平均胸径以及凋落物层厚度,结果见表1。由于季节变化与土壤动物群落特征参数间存在密切关系[11],因此,于2017年秋季(10月)和冬季(12月)以及2018年春季(3月)和夏季(6月)分别对大金山岛12个样地(8个植物群落)进行4次采样;除麻栎群落(样地面积为0.07 hm2)和天竺桂群落(样地面积为0.02 hm2)外,其余植物群落样地面积均为0.04 hm2。在每个样地设置3个样点,样点间隔距离均在5 m以上,样地分布见图1。
中小型土壤动物按生活习性分为湿生(线虫、线蚓和原生生物等)和干生(螨虫和跳虫等)类型[12]。湿生和干生土壤动物分别采用直径3.6和5 cm的不锈钢采样器按0~5、>5~10、>10~15和>15~20 cm分层采集土样,用塑封袋分袋后带回实验室。干生土壤动物采用干漏斗法(Tullgren法)分离,所得土壤动物用φ=75%的乙醇固定;湿生土壤动物采用湿漏斗法(Baremann法)分离,进行活体镜检。所有土壤动物样本参照《中国土壤动物检索图鉴》[13]和《昆虫分类图谱》[14]进行鉴定。
在土壤动物样点旁采集0~20 cm土壤,供测定土壤理化性质。土壤容重、含水量采用环刀法[15]测定,pH采用pH计法(NY/T 1377—2007)测定,全氮含量采用凯氏定氮法(LY/T 1228—2015)测定,全磷含量采用碱熔法(LY/T 1232—2015)测定,全钾含量采用酸消解法-ICP-AES(EPA3050B)测定。

A为麻栎(Quercus acutissima)群落,B为野桐(Mallotus japonicus)群落,C为青冈(Cyclobalanopsis glauca)群落,D为椿叶花椒(Zanthoxylum ailanthoides)群落,E为红楠(Machilus thunbergii)群落,F为朴树(Celtis sinensis)群落,G为丝绵木(Euonymus maackii)群落,H为天竺桂(Cinnamomum japonicum)群落。
表1 研究样地植物群落及群落组成
Table 1 Plant community and community commposition of study plots

群落类型乔木层灌木层草本层 麻栎群落建群种为麻栎,落叶乔木,白檀(Symplocos paniculata)金樱子(Rosa laevigata) 平均胸径为73.48 cm柃木(Eurya japonica)酢浆草(Oxalis corniculata) 红盖鳞毛蕨(Dryopteris erythrosora) 凋落物层厚度为0.02 m 野桐群落建群种为野桐,落叶乔木,野桐金樱子 平均胸径为15.76 cm白檀凋落物层厚度为0.04 m 丝绵木群落建群种为丝绵木,落叶乔木,白檀竹子(Bambusoideae) 平均胸径为31.11 cm构树(Broussonetia papyifera)络石(Trachelospermum jasminoides) 小果蔷薇(Rosa cymosa) 凋落物层厚度为0.06 m 椿叶花椒群落建群种为椿叶花椒,落叶乔木,白檀红盖鳞毛蕨 平均胸径为28.81 cm柃木天南星(Arisaema heterophyllum) 桑树(Morus alba)凋落物层厚度为0.06 m 野桐海州常山(Clerodendrum trichotomum)朴树群落建群种为朴树,落叶乔木,白檀红盖鳞毛蕨平均胸径为22.33 cm构树金樱子海州常山凋落物层厚度为0.04 m 青冈群落建群种为青冈,常绿乔木,白檀海金沙(Lygodium japonicum) 平均胸径为69.92 cm野桐红盖鳞毛蕨 天竺桂(Cinnamomum japonicum)蔷薇(Rosa multiflora) 凋落物层厚度为0.03 m 天竺桂群落建群种为天竺桂,常绿乔木,白檀无 平均胸径为15.81 cm柃木凋落物层较薄 红楠群落建群种为红楠,常绿乔木,柃木红盖鳞毛蕨 平均胸径为46.33 cm野葡萄(Vitis heyneana) 凋落物层厚度为0.12 m
1.3 数据处理
将各季节捕获到的土壤动物个体数换算为密度(ind·m-2),并根据以下公式分别计算中小型土壤动物群落Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(E)、Simpson优势度指数(C)和Margalef丰富度指数(D)。
H′=-∑ni/Nln (ni/N),
(1)
E=H/lnS,
(2)
C=∑(ni/N)2,
(3)
D=(S-1)/lnN。
(4)
式(1)~(4)中,ni为第i个类群个体数;N为群落中所有类群个体总数;S为类群数。采用SPSS 23.0软件进行单因素方差分析(one-way ANOVA),多重比较采用最小显著性检验法(LSD),并对各样地中小型土壤动物密度、主要优势类群密度、类群数、群落多样性指标与土壤理化因子进行Pearson相关性分析。
2 结果与分析
2.1 群落结构
如表2所示,8个植物群落4个季度共捕获中小型土壤动物8 506只,隶属于23个类群,优势类群为线虫纲、蜱螨亚纲和弹尾纲,分别占总密度的66.06%、17.95%和11.15%,其余为稀有类群。
图2显示,按密度比较,常绿阔叶林中小型土壤动物4季平均总密度高于落叶阔叶林,常绿阔叶林春季中小型土壤动物密度显著高于落叶阔叶林(P<0.05);2种植被夏季中小型土壤动物密度均显著高于其他3季(P<0.05)。按类群数比较,常绿阔叶林4季平均类群数显著高于落叶阔叶林(P<0.05),常绿阔叶林春、夏、冬季中小型土壤动物类群数显著高于落叶阔叶林(P<0.05);落叶阔叶林秋季中小型土壤动物类群数显著高于其他3季(P<0.05),而常绿阔叶林秋季中小型土壤动物类群数显著高于春、冬两季(P<0.05)。
表2 大金山岛不同群落类型中小型土壤动物群落组成
Table 2 Composition of soil meso- and micro-fauna in different community types in Dajinshan Island
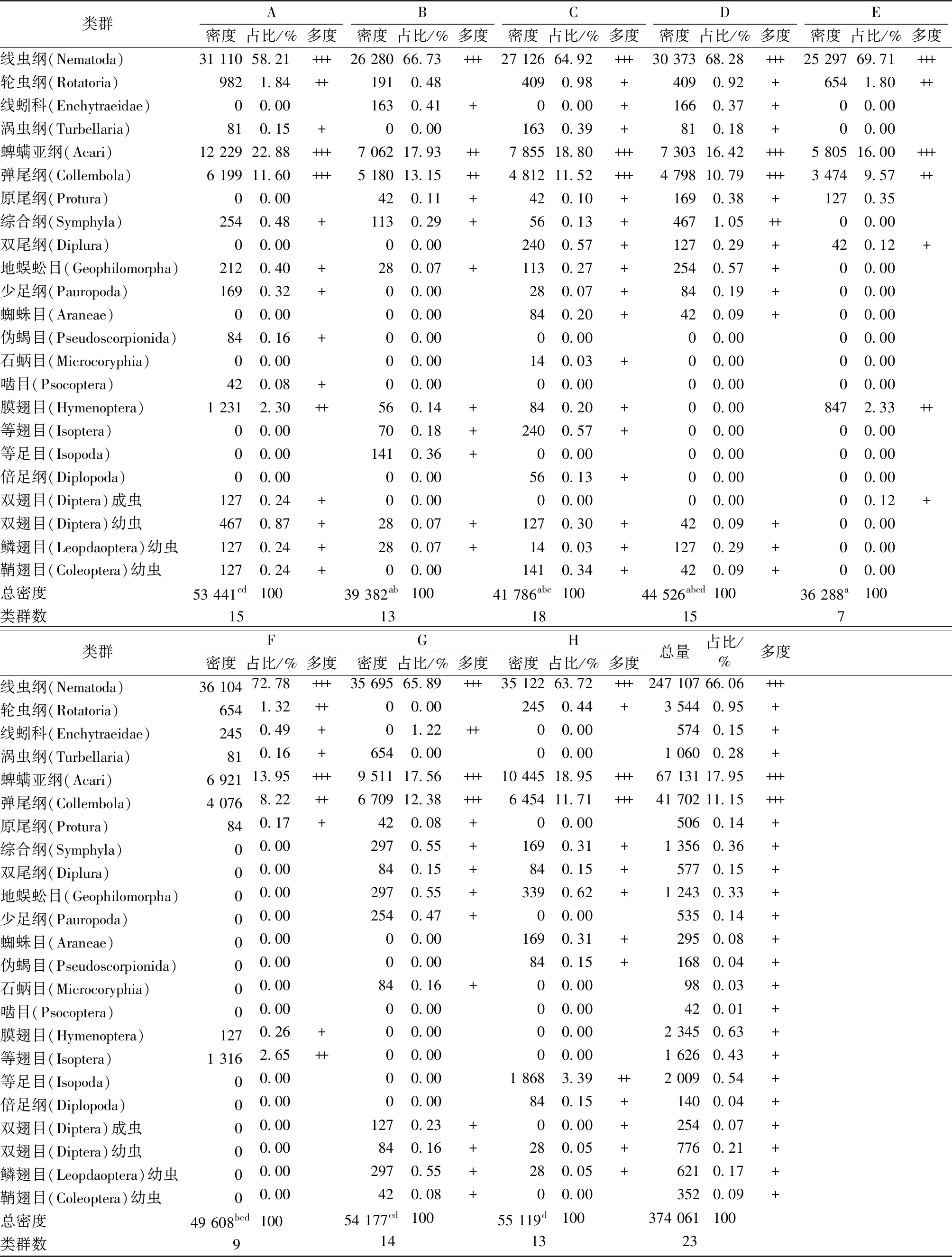
类群ABCDE密度占比/%多度密度占比/%多度密度占比/%多度密度占比/%多度密度占比/%多度 线虫纲(Nematoda)31 11058.21+++26 28066.73+++27 12664.92+++30 37368.28+++25 29769.71+++ 轮虫纲(Rotatoria)9821.84++1910.484090.98+4090.92+6541.80++ 线蚓科(Enchytraeidae)00.001630.41+00.00+1660.37+00.00 涡虫纲(Turbellaria)810.15+00.001630.39+810.18+00.00 蜱螨亚纲(Acari)12 22922.88+++7 06217.93++7 85518.80+++7 30316.42+++5 80516.00+++ 弹尾纲(Collembola)6 19911.60+++5 18013.15++4 81211.52+++4 79810.79+++3 4749.57++ 原尾纲(Protura)00.00420.11+420.10+1690.38+1270.35 综合纲(Symphyla)2540.48+1130.29+560.13+4671.05++00.00 双尾纲(Diplura)00.0000.002400.57+1270.29+420.12+地蜈蚣目(Geophilomorpha)2120.40+280.07+1130.27+2540.57+00.00 少足纲(Pauropoda)1690.32+00.00280.07+840.19+00.00 蜘蛛目(Araneae)00.0000.00840.20+420.09+00.00伪蝎目(Pseudoscorpionida)840.16+00.0000.0000.0000.00 石蛃目(Microcoryphia)00.0000.00140.03+00.0000.00 啮目(Psocoptera)420.08+00.0000.0000.0000.00 膜翅目(Hymenoptera)1 2312.30++560.14+840.20+00.008472.33++ 等翅目(Isoptera)00.00700.18+2400.57+00.0000.00 等足目(Isopoda)00.001410.36+00.0000.0000.00 倍足纲(Diplopoda)00.0000.00560.13+00.0000.00 双翅目(Diptera)成虫1270.24+00.0000.0000.0000.12+ 双翅目(Diptera)幼虫4670.87+280.07+1270.30+420.09+00.00 鳞翅目(Leopdaoptera)幼虫1270.24+280.07+140.03+1270.29+00.00 鞘翅目(Coleoptera)幼虫1270.24+00.001410.34+420.09+00.00 总密度53 441cd10039 382ab10041 786abc10044 526abcd10036 288a100 类群数 15 13 18 15 7 类群FGH密度占比/%多度密度占比/%多度密度占比/%多度总量占比/%多度 线虫纲(Nematoda)36 10472.78+++35 69565.89+++35 12263.72+++247 10766.06+++ 轮虫纲(Rotatoria)6541.32++00.002450.44+3 5440.95+ 线蚓科(Enchytraeidae)2450.49+01.22++00.005740.15+ 涡虫纲(Turbellaria)810.16+6540.0000.001 0600.28+ 蜱螨亚纲(Acari)6 92113.95+++9 51117.56+++10 44518.95+++67 13117.95+++ 弹尾纲(Collembola)4 0768.22++6 70912.38+++6 45411.71+++41 70211.15+++ 原尾纲(Protura)840.17+420.08+00.005060.14+ 综合纲(Symphyla)00.002970.55+1690.31+1 3560.36+ 双尾纲(Diplura)00.00840.15+840.15+5770.15+ 地蜈蚣目(Geophilomorpha)00.002970.55+3390.62+1 2430.33+ 少足纲(Pauropoda)00.002540.47+00.005350.14+ 蜘蛛目(Araneae)00.0000.001690.31+2950.08+ 伪蝎目(Pseudoscorpionida)00.0000.00840.15+1680.04+ 石蛃目(Microcoryphia)00.00840.16+00.00980.03+ 啮目(Psocoptera)00.0000.0000.00420.01+ 膜翅目(Hymenoptera)1270.26+00.0000.002 3450.63+ 等翅目(Isoptera)1 3162.65++00.0000.001 6260.43+ 等足目(Isopoda)00.0000.001 8683.39++2 0090.54+ 倍足纲(Diplopoda)00.0000.00840.15+1400.04+ 双翅目(Diptera)成虫00.001270.23+00.00+2540.07+ 双翅目(Diptera)幼虫00.00840.16+280.05+7760.21+ 鳞翅目(Leopdaoptera)幼虫00.002970.55+280.05+6210.17+ 鞘翅目(Coleoptera)幼虫00.00420.08+00.003520.09+ 总密度49 608bcd10054 177cd10055 119d100374 061100 类群数 9 14 13 23
+++优势类群(>10%);++常见类群(1%~10%);+稀有类群(<1%)。A为椿叶花椒群落,B为野桐群落,C为青冈群落,D为朴树群落,E为麻栎群落,F为丝绵木群落,G为天竺桂群落,H为红楠群落;同一行数据后英文小写字母不同表示不同植被类型间中小型土壤动物总密度差异显著(P<0.05)。空白处表示无数据。
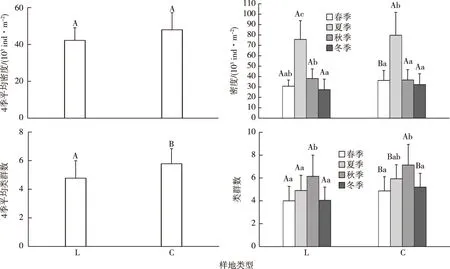
L为落叶阔叶林,C为常绿阔叶林。各分图中直方柱上方英文小写字母不同表示相同植被类型不同季节间某指标差异显著(P<0.05),英文大写字母不同表示相同季节不同植被类型间某指标差异显著(P<0.05)。
2.2 垂直分布
图3显示,大金山岛森林植被中小型土壤动物表现出明显的表聚性,0~5 cm土层中小型土壤动物密度占0~20 cm土层总密度的67.81%;常绿阔叶林和落叶阔叶林0~5 cm土层中小型土壤动物密度分别占0~20 cm土层总密度的68.68%和66.94%;4个土层中小型土壤动物密度呈明显递减趋势,且湿生土壤动物密度均高于干生土壤动物。

右分图中,L为落叶阔叶林,C为常绿阔叶林,英文大写字母后的1、2、3和4分别表示土层深度为0~5、>5~10、>10~15和>15~20 cm。
2.3 多样性指数的季节变化
如图4所示,常绿阔叶林秋季中小型土壤动物群落Shannon-Wiener多样性指数和Margalef丰富度指数均显著高于其他3季(P<0.05);落叶阔叶林秋季中小型土壤动物群落Margalef丰富度指数均显著高于其他3季(P<0.05)。常绿阔叶林夏季中小型土壤动物群落Shannon-Wiener多样性指数显著高于落叶阔叶林(P<0.05),而常绿阔叶林春、夏、冬中小型土壤动物群落Margalef丰富度指数显著高于落叶阔叶林(P<0.05)。
2.4 环境因子与中小型土壤动物的相关性
由表3可知,常绿阔叶林和落叶阔叶林土壤含水量和pH无显著差异(P>0.05),落叶阔叶林土壤容重显著高于常绿阔叶林(P<0.05);常绿阔叶林土壤有机质、全N、全P和全K含量显著高于落叶阔叶林(P<0.05)。结果表明相较于落叶阔叶林,常绿阔叶林土壤较为疏松,养分含量较为丰富。

L为落叶阔叶林,C为常绿阔叶林。各分图中直方柱上方英文小写字母不同表示相同植被类型不同季节间某指标差异显著(P<0.05),英文大写字母不同表示相同季节不同植被类型间某指标差异显著(P<0.05)。
表3 大金山岛常绿阔叶林和落叶阔叶林土壤理化性质
Table 3 Soil physicochemical properties in evergreen broad-leaved forest and deciduous broad-leaved forest in Dajinshan Island

样地含水量w/%容重/(g·cm-3)pHw(全N)/(g·kg-1)w(全P)/(g·kg-1)w(有机质)/(g·kg-1)w(全K)/(mg·kg-1) 落叶阔叶林27.12±4.58a0.92±0.20b4.86±0.95a4.82±1.09a1.28±0.41a13.91±4.28a595.16±89.15a 常绿阔叶林27.29±5.52a0.75±0.22a4.56±0.83a5.94±1.44b1.70±0.44b17.16±2.88b680.13±112.79b
同一列数据后英文小写字母不同表示不同植被类型间某指标差异显著(P<0.05)。
对不同植被类型土壤理化性质及中小型土壤动物总密度、主要优势类群密度、类群数和群落多样性指标作Pearson相关系分析。由表4可知,土壤全N含量与土壤容重呈极显著负相关(P<0.01),土壤全P含量与土壤pH呈显著负相关(P<0.05),土壤全K含量与土壤全P含量呈极显著正相关(P<0.01),其余各土壤理化因子间无显著相关性;中小型土壤动物类群数及Margalef丰富度指数与土壤全K呈显著负相关(P<0.05),其余指标与土壤理化因子无显著相关性。由表5可知,弹尾纲密度与土壤有机质含量呈显著正相关(P<0.05),蜱螨亚纲密度与土壤pH值呈显著负相关(P<0.05),其余土壤理化因子与主要优势类群密度无显著相关性。
3 讨论
3.1 大金山岛森林植被土壤动物群落的影响因素
大金山岛由于其明显的海域隔离和较少的人为干扰,植物和动物的生活环境近似自然。笔者研究发现大金山岛干生土壤动物优势类群主要为蜱螨目和弹尾目,与靳亚丽等[8]在大金山岛所调查的有关干生土壤动物的结果一致。此外,笔者研究额外捕获到石蛃目(表2),但并未捕获到靳亚丽等[8]发现的腹足纲动物。相比同纬度大陆城市森林生态系统和自然森林生态系统,大金山岛中小型土壤动物密度及类群数均高于城市森林生态系统,但低于自然森林生态系统[12]。相比同纬度自然森林,海岛生态系统易受损且难恢复[16],这可能是大金山岛中小型土壤动物密度和类群数较少的原因。而相比邻近大陆的城市森林生态系统,人为干扰是限制土壤动物繁殖生存的主要因素[17],大金山岛相对自然的生存环境较其更为适合土壤动物生存。
表4 中小型土壤动物群落总密度、类群数、群落多样性指数与土壤理化性质相关性分析
Table 4 Correlation analysis of total density, group number, community diversity and soil physicochemical properties of soil meso- and micro-fauna

指标WdvpHOCKNPSTDH′CE dv-0.230 1 pH-0.2820.2841 OC-0.2120.097-0.0351 K-0.156-0.067-0.0420.2591 N0.278-0.654∗∗-0.3160.0700.1461 P0.069-0.202-0.357∗0.2400.504∗∗0.3281 S0.263-0.207-0.280-0.239-0.393∗0.266-0.0241 TD0.1140.096-0.3180.255-0.0760.1870.1370.629∗∗1 H′0.097-0.121-0.312-0.160-0.0950.177-0.1860.715∗∗0.499∗∗1 C-0.2460.1620.0230.1200.298-0.207-0.201-0.732∗∗-0.433∗-0.1571 E-0.0070.0490.2410.072-0.104-0.1340.169-0.366∗-0.290-0.856∗∗-0.2111 D0.261-0.189-0.253-0.256-0.408∗0.225-0.0410.995∗∗0.616∗∗0.703∗∗-0.738∗∗-0.350∗
W为土壤含水量,dv为土壤容重,OC为土壤有机质含量,K为全钾含量,N为全氮含量,P为全磷含量,S为土壤动物类群数,TD为土壤动物总密度,H′为Shannon-Wiener多样性指数,C为Simpson优势度指数,E为Pielou均匀度指数,D为Margalef丰富度指数。*和**分别表示相关显著(P<0.05)和相关极显著(P<0.01)。
表5 中小型土壤动物主要优势类群密度与土壤理化性质相关性分析
Table 5 Correlation analysis of main dominant group density and soil physicochemical properties of soil meso- and micro-fauna

指标WdvpHOCKNP线虫纲蜱螨亚纲 dv-0.230 1 pH-0.2820.2841 OC-0.2120.097-0.0351 K-0.156-0.067-0.0420.2591 N0.278-0.654∗∗-0.3160.0700.1461 P0.069-0.202-0.357∗0.2400.504∗∗0.3281 线虫纲-0.0190.132-0.1080.333-0.0960.0560.2811 蜱螨亚纲0.060-0.057-0.387∗0.1140.0020.2690.0040.406∗1 弹尾纲-0.0540.142-0.1650.360∗0.2860.0830.2540.430∗0.497∗∗
W为土壤含水量,dv为土壤容重,OC为土壤有机质含量,K为全钾含量,N为全氮含量,P为全磷含量,线虫纲、蜱螨亚纲和弹尾纲为此3种优势类群密度。*和**分别表示相关显著(P<0.05)和相关极显著(P<0.01)。
植被类型对土壤动物的影响一直是学者们的研究热点[18]。不同植被类型凋落物层的组成、厚度和质量等是影响土壤动物密度的重要因素[19]。笔者研究中,红楠群落可能由于其成林时间较早,较其他群落类型累积了更多的凋落物,对其下的土壤动物起到了一定的促进作用,导致其土壤动物总密度和线虫纲密度均位于前列(表2)。不同植被类型的植物群落组成复杂程度也会对土壤动物密度和类群数产生影响[20]。通常,植物群落组成越复杂,其能给土壤动物提供的小生境就越多,类群数也会有所提高。笔者研究中,青冈群落、椿叶花椒群落和朴树群落的组成较为复杂,灌木层和草本层植物种类繁多,这3种群落类型土壤动物类群数相较于其他群落类型都要位于前列(表2),表明群落组成复杂程度对土壤动物群落具有一定的积极影响。
不同植被类型由于其建群种、成林时间和各种微地形气候等的不同,导致其群落环境内土壤环境因子有所差异。已有文献表明,通常,常绿阔叶林土壤动物密度和类群数都要高于落叶阔叶林[21]。这可能是由于常绿阔叶林土壤养分含量在一般情况下高于落叶阔叶林[22],笔者研究也支持这一观点,因此养分含量较高的常绿阔叶林较适合土壤动物生存。笔者研究发现中小型土壤动物密度及其优势类群密度均与土壤有机质含量呈正相关,与土壤pH呈负相关(表4~5)。土壤有机质作为土壤养分含量中的重要指标,其对中小型土壤动物个体数一般具有正向促进作用[23]。通常,土壤动物适宜生存在微酸性和中性土壤中[24],笔者研究发现中小型土壤动物密度与土壤pH呈负相关的原因可能在于酸性程度较高的样地具有较高的土壤养分含量,而长期生存也提高了土壤动物对于酸性土壤的耐受能力。笔者研究发现中小型土壤动物类群数和Margalef丰富度指数均与土壤全钾含量呈显著负相关(表4),韩慧莹等[23]研究同样发现中小型土壤动物类群数对土壤全钾含量具有显著负向响应,2项研究结果一致。
3.2 土壤动物群落的时空格局
温度和降雨是影响土壤动物群落的重要因素,而此2种因素通常随着季节更替而变化,因此不同季节间土壤动物密度和类群数通常具有差异性。笔者研究显示,大金山岛森林植被中小型土壤动物秋季(10月)类群数最高,夏季(6月)密度最高(图2),此与王邵军等[25]对武夷山典型植被群落土壤动物群落结构的研究结果相类似。其他研究[26]表明,土壤动物类群数在秋季最高,说明相较于植被类型,气候条件可能是造成土壤动物类群数季节变化更为主要的原因。值得注意的是,笔者研究中线虫纲密度占土壤动物总密度的66.06%(表2),表明线虫纲密度对土壤动物总密度的影响较大。故笔者研究中夏季(6月)中小型土壤动物大量发生主要在于线虫大量繁殖,而线虫大量繁殖可能与大金山岛所处的亚热带海洋性气候有关,夏季降水多、土壤含水量较高可能是其原因,此与李志鹏等[27]对哀牢山常绿阔叶林线虫群落的研究结果一致,雨季线虫纲密度在不同林地均要大于干季。
土壤动物个体数量和类群数的季节变化导致群落多样性指数的季节变化[28],笔者研究中常绿阔叶林秋季中小型土壤动物群落Shannon-Wiener多样性指数和Margalef丰富度指数均显著高于其他3季(P<0.05,图4),落叶阔叶林秋季Margalef丰富度指数显著高于其他3季(P<0.05,图4),结果表明秋季大金山岛森林植被中小型土壤动物群落多样性水平高于其他3季。对比易兰等[29]对天童栲树林土壤动物的研究发现,2项研究中土壤动物群落多样性水平均表现为秋季最高。就笔者研究而言,秋季较高的群落多样性水平主要在于类群数最多,而主要优势类群在4季均有发生,因此稀有类群的季节变化对不同季节群落多样性水平的影响较大。
在自然植被土壤动物垂直分布方面,已有文献大部分表明土壤动物具有明显的表聚性[30],笔者研究也支持这一观点。这可能是由于随着土层加深,土壤养分含量减少,土壤容重增加,紧实、贫瘠的土壤限制了土壤动物的生存。
4 结论
大金山岛森林植被中小型土壤动物密度夏季(6月)最高,类群数秋季(10月)最高。大金山岛森林植被中小型土壤动物表现出明显的表聚性。大金山岛常绿阔叶林中小型土壤动物群落多样性水平高于落叶阔叶林。
致谢:感谢上海交通大学环境科学与工程学院程金平老师课题组冯晶晶博士在大金山岛野外工作中和在提供该文所使用的环境因子数据方面给予的帮助。
猜你喜欢
农业科技与信息(2020年14期)2020-12-18
书城(2020年1期)2020-01-15
绿色中国(2019年18期)2020-01-04
快乐学习报·教研周刊(2020年11期)2020-01-03
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
绿色科技(2018年19期)2018-11-29
农家科技下旬刊(2017年8期)2017-11-13
科技创新导报(2016年21期)2016-12-17
