
生物分子液-液相分离的物理化学机制
2020-04-02 02:52张长胜来鲁华
物理化学学报 2020年1期
张长胜,来鲁华
1北京大学化学与分子工程学院,物理化学研究所,分子动态与稳态结构国家重点实验室,北京分子科学国家研究中心,北京 100871
2北京大学定量生物学中心,北京大学-清华大学生命科学联合中心,北京 100871
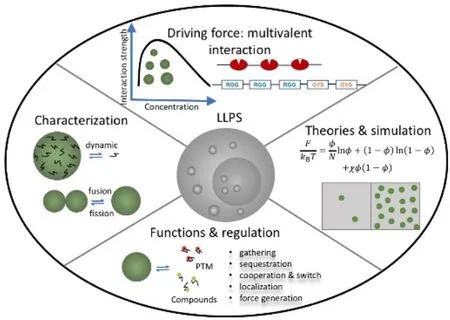
1 引言
2009年Hyman、Brangwynne等人用PGL-1(Guanyl specific ribonuclease 1)等分子参与的相分离的理论解释了线虫受精卵细胞极化和不对称分裂机制这个困扰该领域至少25年的难题1,2。该理论简单自然,实验证据丰富,启发了生命科学其它领域的研究者认识到相分离可能是一个普遍的生物分子组织定位、功能调控的机制。随后细胞的相分离机制在发育1,2、环境压力应激3、基因转录调控4,5、DNA修复6、RNA剪接加工与转运7,8、信号转导9,10、固有免疫11、神经信号传递12等大量的生命活动过程中被发现。生命体通过生物分子相分离产生具有各种功能的高度动态的无膜细胞器的概念被广泛接受13,相分离机制在生命活动中至关重要的作用开始逐渐清晰14。近几年来对生物体系相分离的研究更是呈现井喷式的增长(图1展示了PUBMED文献数据库https://www.ncbi.nlm.nih.gov/pubmed/收录的关于生物体系液-液相分离文献在近20年中的变化情况)。
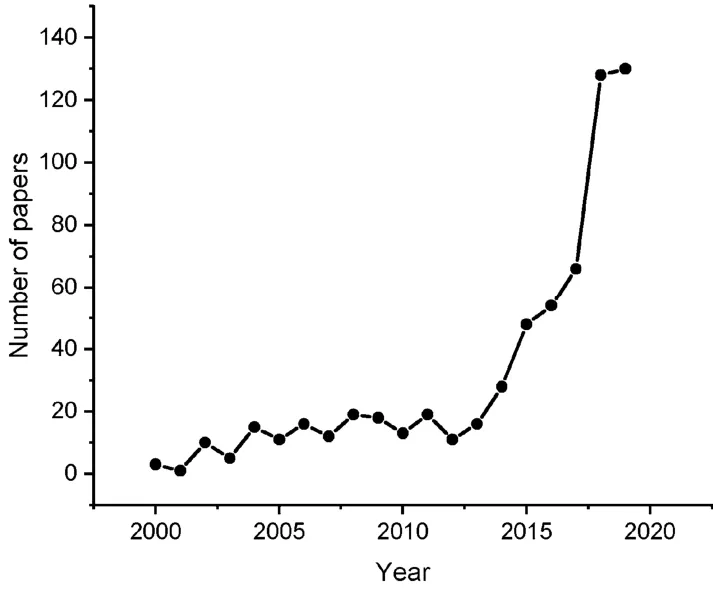
图1 PUBMED数据库收录的2000-2019年间液-液相分离文献逐年快速增长情况Fig.1 Rapid growth of literatures on liquid-liquid phase separation from 2000 to 2019 in PUBMED.
多组分体系相转变是物理化学领域研究的基本问题之一。由于生物分子序列与结构复杂多样,研究生物分子的相分离机制对于物理化学学科来说也是一个新的重要挑战。需要发展新的实验、模拟和理论研究方法,从而深入认识相分离的物理化学机制,包括其热力学、动力学、聚集体微观结构、序列与相分离条件和聚集体性质关系、相分离的驱动力和调控方式等。
本文将从相分离聚集体的基本性质、相图、微观结构、统计热力学、实验和分子模拟研究方法、相分离性质与生物功能关系、调控分子设计这七个方面简介基本认识、阐述当前主要的研究进展、总结梳理主要的研究结果,为进一步的生物分子相分离物理化学机制的研究提供借鉴。
2 液-液相分离液滴基本性质及表征方法
2.1 形貌
生物分子在体外形成的液滴一般为微米级大小,在溶液中自由状态下呈球形,具有液-液两相界面张力所导致的光滑表面。微分干涉差显微镜(differential interference contrast microscopy)、宽场荧光显微镜(wide-field fluorescence microscopy)是观察聚集体状态的主要手段,软X射线光谱15可用于研究其三维结构。溶液中微米级的相分离液滴可以引起可见光的散射,散射的强度,即浊度(turbidity)主要跟液滴的尺寸和数量相关。通过浊度的测量可以很便捷的获得体系相分离的临界条件14,16。体外实验中观察到的溶液中生长的液滴一般比细胞内的相分离液滴大10-1000倍17,Shayegan等人用凸透镜诱导约束(convex lens-induced confinement)技术,模拟细胞内的受限环境,限制体外液滴的大小,并用荧光显微镜观察RNA polyU和RNA结合蛋白Dhh1形成的液滴,大小受限后液滴由布朗运动变为跳跃式运动18。
2.2 融合与浸润
相分离过程首先是在过饱和溶液中,聚集体分子快速成核并生长形成初始液滴(生长growth),液滴在溶液中自由移动,小液滴之间逐渐碰撞凝聚融合成大液滴(粗化coarsening),剧烈振动下大液滴可以分裂为小液滴,与固体表面接触有浸润现象。小液滴融合成为大液滴的速度可以用融合液滴的纵横比的变化来粗略表征,也可以通过双光阱光镊(dual-trap optical tweezer)实现对两个液滴的控制和融合速度的测量19,20。理论上融合速率与液滴粘度和表面张力的比值呈负相关21,22。Berry等人23研究了核仁蛋白FIB在体外相分离过程中液滴生长、粗化的动力学,实现结果表明粗化过程中液滴的平均半径与时间的1/3次幂成正比,他们还发现含RNA的液滴粗化更快,而溶解过程速度更慢。对于成分不同,不能互溶的液滴相互碰撞,依据表面张力的不同,形成分散或分层融合状态24;当两液滴之间的界面张力小于它们与溶剂的界面张力时形成分层融合结构,例如在核仁中有从内到外FC-DFC-GC三层液态结构25。
2.3 液滴内分子的运动性
液滴内各组分呈自由运动状态,液滴内外分子可动态交换,常用荧光光漂白恢复(fluorescence recovery after photobleaching)技术定量研究液滴组分的运动性。荧光相关光谱(Fluorescence correlation spectroscopy)技术通过定量测量液滴微区内的荧光涨落,可获得第二维里系数、分子扩散系数、黏度、以及浓度等数据。Wei等人26用此技术描绘了LAF-1蛋白体系的包含高浓度分支的完整的相分离相图,并研究了体系形成的液滴中LAF-1分子的运动性质,他们发现LAF-1与长链RNA分子polyrA3k共同作用产生的液滴中LAF-1蛋白的浓度及密度比纯LAF-1液滴的值显著降低,而液滴粘度却显著升高。脉冲梯度场核磁共振(Pulsed-field gradient NMR)也用于研究液滴内分子的扩散系数。Brady等人27应用此技术测定的Ddx4在凝聚相中和溶液相中的扩散系数分别为7.5 × 10-9和8.8 ×10-7cm2·s-1。他们对液滴中LAF-1分子主链15N的R2弛豫速率的测量结果是16.4 s-1,表明蛋白质主链仍具有显著的运动性。Reichheld等人28应用核磁共振技术研究了弹性蛋白ELP3液滴内分子的运动性质和结构,得到类似的结果。
2.4 渗透性
非专一性渗透的分子通过液滴“孔隙”渗透到液滴内部的能力跟分子大小有关,可用不同粒径的葡聚糖(Dextran)研究液滴的渗透性26,29。如蛋白质LAF-1液滴可以透过10kDa,直径约3纳米的葡聚糖分子26。专一性的渗透一般由进入液滴的客体分子和基架分子专一性的相互作用介导。例如转录因子雌激素受体(ER)含有配体结合结构域(LBP),只有当雌激素配体结合于LBP之后,ER转换至与相分离基架蛋白MED1 (LXXLL区)结合状态,ER才会被招募至MED1液滴中5。
3 影响生物大分子液液相分离发生的条件及相图
3.1 分子浓度
一定条件下,当生物分子浓度较稀时,生物分子均一地溶于溶剂中,当超过临界饱和浓度cS时,生物分子由于相互吸引作用而凝聚,体系发生相分离,从溶液中析出生成新的相,聚集相液滴具有浓度cD,当生物分子浓度超过cD时,体系又变为单一相。一般通过不同浓度下的观察相分离是否发生,来确定体系的临界饱和浓度cS,临界饱和浓度越低说明体系越易于发生相分离。对于两种生物分子相互作用产生的相分离,一般用二维相图研究相分离发生的浓度条件。
3.2 温度
温度对相分离的影响体现了相分离过程的熵效应。多数的生物分子相分离过程是体系熵减少的过程所以温度的升高不利于相分离的进行。在温度-浓度二维相图上存在最高临界共溶温度(upper critical solution temperature),例如静电区块相互作用驱动的Ddx4蛋白27。对于相分离的溶剂熵或离子熵效应显著的体系,相分离使体系熵增加,温度的升高有利于相分离的进行,在温度-浓度二维相图上存在最低临界共溶温度(lower critical solution temperature)。例如依靠疏水作用聚集的弹性蛋白30,和依靠Lys/Gly rich区域相分离的Tau蛋白31。
3.3 离子强度(盐浓度)
对于一价金属离子(Na+,K+),静电作用对于多数的生物分子的相分离具有关键作用,提高盐浓度减弱静电作用,所以一般增加盐浓度不利于相分离的发生,使临界饱和浓度cS增加,聚集体粘度也会下降27。然而如果体系中疏水作用在蛋白分子间相互作用中起主要作用,那么增加盐浓度将有利于该蛋白的相分离,例如RNA结合蛋白FUS的N端低复杂度(LC)结构域32。对于二价金属(Mg2+,Zn2+)提供多价相互作用促进相分离的体系,其浓度超过临界浓度时相分离才可发生11,33。
3.4 pH值
pH值影响氨基酸的质子化状态。增强分子间相互作用的pH值条件促进相分离的发生。例如酵母细胞在受到环境压力后细胞质pH值下降,将促进天然无序蛋白SUP35的相分离3。
3.5 拥挤效应
体外实验中常用加入PEG(聚乙二醇)分子的方法引入拥挤效应,以模拟细胞中的拥挤环境。拥挤效应提高了生物分子的有效浓度,会促进相分离的发生34。
3.6 核酸分子对蛋白质相分离的影响
核酸分子对蛋白质相分离的影响主要取决于核酸分子是否增强了体系的多价相互作用。若核酸分子与蛋白质分子有较强专一性的多价相互作用,那么核酸分子将促进蛋白质相分离的发生;若核酸分子的强带电性主要起到的是类似盐离子的静电屏蔽作用,削弱蛋白质之间的相互作用,那么核酸分子的加入会抑制相分离的发生;若同时存在核酸-蛋白作用和静电屏蔽两种效应,但是都较弱,那么一般是在核酸浓度较低时促进相分离,核酸浓度较高时抑制相分离,例如FUS蛋白LC结构域32以及Banerjee等人构建的模式体系35。Zhou实验室36用表面带3个或2个相互作用区块的小球模拟蛋白质和核酸分子的多价和静电作用,用Gibbs系综蒙特卡洛模拟的方法37,38研究了核酸/蛋白质体系在不同的蛋白质-核酸作用强度下的两相平衡(即两相共存)浓度,得到相图。
3.7 相分离数据库
Zhang实验室建立的生物体系液-液相分离数据库(LLPSDB)39较全面的收集了生物分子相分离发生条件的实验数据,为相分离的物理化学机制研究提供了基础。
4 驱动液-液相分离的生物分子结构特征
相分离的主要驱动因素是多价相互作用,同时液-液相分离需要结构上保证相互作用的动态性。本文将提供多价相互作用的结构总结为三大类,并提供部分实例,更多的例子可以参考近期的其他综述文章13,40,41。
4.1 由多个重复结构域特异性作用介导的相分离
第一大类是利用特异性的蛋白质相互作用,其中重复的结构域提供多价作用,重复结构域之间的柔性提供相互作用的动态性。主要有两类体系:
(1)多个相同/相似结构域的线性串联(图2a所示),例如NCK等蛋白质的SH2,SH3结构域重复,N-WASP蛋白的PRM结构域重复42,多泛素化43等。
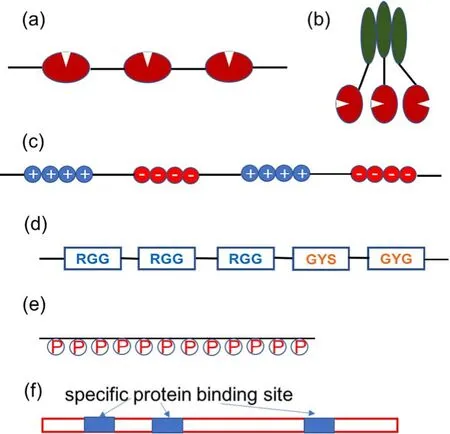
图2 相分离体系提供多价相互作用的类型示例Fig.2 Illustration of multivalent force types in biomolecular phase separation systems.
(2)蛋白质的寡聚(图2b所示),如突触后致密物PSD中的PSD-95是底物结合诱导的二聚体,SynGAP蛋白通过Coiled结构域形成三聚体44,核仁蛋白NPM1通过N端结构域形成五聚体25等。
4.2 由蛋白质无序区域介导的相分离
第二大类是由蛋白质天然无序区(intrinsically disordered proteins/domains)或称低复杂度结构域(low complicity domains)驱动形成的相分离45。大量的重复结构片段可以提供多价作用,固有的无序结构提供相互作用的动态性46。这类蛋白相比于第一大类可以提供更多价、更灵活多样的相互作用方式,因此驱动生物体系大多数液态聚集体形成的基架分子为这类蛋白。有以下典型的例子:
(1)多个带相同电荷残基集中区域(图2c所示)。例如在Ddx4的N端结构域(1-236)包含多段正电荷残基(Arg/Lys)或负电荷残基(Glu/Asp)集中分布的区域47,48,提供多价的静电相互作用。
(2)含多个RGG以及(G/S)Y(G/S)单元(图2d所示),例如FUS,LAF1等核蛋白,精氨酸的胍基与酪氨酸的芳香环有cation-π以及类似π-π的相互作用,酪氨酸之间也可以形成π-π相互作用,精氨酸的胍基与核酸的磷酸基团有强静电作用,以及与碱基有cation-π以及类似π-π的较强相互作用。Vernon等人49详细研究了π-π或类似π-π相互作用在蛋白质相分离中的重要作用。Wang等人19研究了22个FUS家族的蛋白质的低复杂度结构域相分离的临界饱和浓度,发现与Arg、Tyr残基数目的乘积成反比,将FUS中的精氨酸突变为赖氨酸,或将酪氨酸突变为苯丙氨酸都会明显降低蛋白的相分离能力。他们的研究也发现Gly对于增加液滴内蛋白的运动性,维持聚集体的流动性很重要。
(3)富含Gln/Asn序列。这是多数朊蛋白样结构域的特征。天冬酰胺、谷酰胺残基侧链之间以及侧链与主链之间氢键50、以及其它偶极作用是这类蛋白聚集的关键驱动力51。例如亨廷顿蛋白(HTT exon1)的聚集主要是其polyQ序列提供的多价作用驱动的,并且其中谷酰胺(Q)的数目决定了聚集体的形态,过长的polyQ序列会导致不可逆的纤维样聚集体的形成,与疾病直接相关52。Fiumara等人分析了富含Gln/Asn以及polyQ序列的卷曲螺旋结构,他们认为卷曲螺旋结构对于促进这类蛋白的聚集具有重要作用53。
(4)在弹性蛋白(Elastin)中,VPG等序列重复出现,其疏水作用提供相分离驱动力30。
4.3 核酸分子介导的相分离
第三大类是核酸分子(DNA或RNA),单链核酸的柔性为相互作用提供动态性。在模式体系(表1g)中的研究发现单链核酸与蛋白质易于聚集成液滴,而双链核酸较为刚性,与蛋白质易于形成凝胶状聚集体54。
核酸分子提供的多价作用有三种类型。(1)核酸分子的主链磷酸基团可以提供多价的静电作用(图2e所示)。这种多价作用类型没有序列特异性。(2)核酸序列的重复扩展也为其依靠碱基对的多价性发生核酸分子自身的相分离(如表1a的(CAG)或(CUG) repeat RNA可发生不依赖蛋白质的相分离55)。(3) RNA序列中存在重复多个特异结合某个蛋白质的区块,这些区块为其与该蛋白质的相分离提供较专一的多价作用(图2f所示)。相同的蛋白质分子与不同多价结构的RNA分子相分离,会导致形成不同性质、不同功能的聚集体。Longdon等人的研究发现33,56,在霉菌Ashbya gossypii细胞中,三种mRNA BNI1、SPA2与CLN3都与Whi3蛋白作用发生相分离,但是BNI1与SPA2的液滴可以共定位于细胞的尖端,而CLN3液滴具有更大的密度与前两者不能融合,定位于细胞核周围。他们的研究揭示发现这种差异性来源于BNI1/SPA2二级折叠结构中的Whi3结合位点的分布不同。它们的序列中都含有5个Whi3结合位点,但是在CLN3中位点间隔显著小于BNI1/SPA2,导致CLN3与Whi3形成的液滴相比于BNI1/SPA2液滴具有更大的粘度,更大的密度和内部分子更差的运动性56。另外CLN3二级折叠结构中与BNI1或SPA2互补的序列被埋藏,导致CLN3不能进入BNI1/SPA2液滴,不能发生CLN3与BNI1/SPA2液滴的融合。CLN3重折叠改变二级结构,暴露互补序列后就可以被BNI1/SPA2液滴招募33。

表1 研究生物分子相分离物理化学机制的模式实验体系Table 1 The model systems for experimental studying the physiochemical mechanisms of biomolecular phase separation.
5 影响生物大分子液液相分离发生的条件及相图
5.1 Flory-Huggins理论
从热力学角度看,相分离的发生降低了体系整体的自由能57。体系的自由能可分解为焓变和熵变的贡献,焓变包含生物分子与溶剂之间、生物分子之间以及溶剂分子之间势能的变化,熵变衡量体系自由度的变化。Flory58和Huggins59在上世纪40年代提出了高分子溶液的“似晶格”模型,用高分子的体积分数φ作为参数,推导出了理想高分子溶液混合熵变的计算公式,以及用平均作用场假设,引入参数χ描述相互作用能的变化,得到了混合焓变的理论计算公式,混合自由能变F的计算公式即为公式(1),
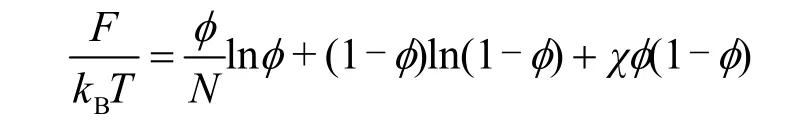

式中kB为玻尔兹曼常数,T为温度,N为高分子链的长度,z为晶格的配位数,ups、upp、uss分别为高分子的一个链段与一单位溶剂之间、一对链段之间、一对溶剂之间的结合能。Flory和Huggins的理论以十分简洁、粗略的形式解释了包含生物大分子在内的高分子相分离过程的热力学机制。
Flory和Huggins的理论中体系由于混合熵变总为负,当χ取较大的正值时,混合自由能会取到正值,也就是体系趋向于两相,而不是混合,这时混合自由能对浓度(体积分数)的函数存在两个谷值点,即为相分离的临界浓度值60。
5.2 其它统计热力学理论
在Flory和Huggins的焓变公示推导过程中只考虑了近程的相互作用,Overbeek和Voorn61在他们的工作基础上考虑了长程静电相互作用,应用于带Z个正电和Z个负电而整体不带电的聚电解质高分子溶液。他们假设这些电荷随机的散布于溶液中,与盐溶液类似可以用Debye-Hückel理论假设计算体系中的电荷间相互作用的能量。之后,随机相位近似(random phase approximation,RPA)方法被应用于的聚电解质高分子溶液静电作用的计算,该方法可用于任意电荷分布样式的聚电解质高分子溶液体系62,63。Lin等人48,64将该方法应用于蛋白质分子相分离体系的计算,解释了同种电荷在序列上集中分布对于相分离的驱动作用,以及离子强度对于静电作用为主的相分离体系的影响。场理论模拟(Field theory simulation)方法是更细致处理聚电解质高分子溶液体系的方法,通过对场理论中的共轭变量进行模拟采样,来获得体系的相图。McCarty等人65用离散的链模型表示不同Lys/Glu肽链,用场理论模拟方法获得了它们的盐浓度-蛋白浓度相图,得到跟RPA理论相近的结果。
6 研究生物分子液-液相分离物理化学机制的手段
天然发生相分离的生物大分子序列复杂导致其构象复杂,分子间相互作用多样,从实验上细致研究分子序列、结构与相分离条件、聚集体性质之间的关系比较困难。当前的研究试图从两个方面解决这个问题,一是寻找并研究简化的相分离模式实验体系,用简单重复序列研究电荷性质、残基种类、序列长度等参数对相分离的影响;另一方面是利用分子模拟的方法,利用与序列相关的力场势能函数,研究预测体系相分离的性质和规律。
6.1 利用模式体系实验研究生物分子相分离
表1收集了不同类型的蛋白质/核酸相分离的模式体系研究对象及主要结果。包括单核酸(a)、柔性连接(b、c)、天然无序蛋白质(d)、蛋白质+核酸(e、f、g)、蛋白质+蛋白质(h、i)等体系。
6.2 生物分子液-液相分离过程的分子模拟
生物分子相分离是众多分子通过随机的多价作用形成,因此相分离的模拟体系需要包含足够数目的分子,参与相分离的蛋白质或核酸又往往包含几十到几百的残基,因此相分离体系的模拟难以依靠原子级精细的力场模拟完成,一般需要粗粒化的模型来完成。最近,Dignon等人70对天然无序蛋白质的液-液相分离模拟方法作了简要的综述。本文将着重介绍Mittal实验室71开发的用于长条形盒子动力学模拟(Slab simulation)的蛋白质粗粒化方法以及Pappu实验室72开发的格点模型下模特卡罗模拟的粗粒化方法。Chan实验室73,74在生物分子相分离的模拟分子模拟方法方面也有较深入的研究。
6.2.1 长条形盒子动力学模拟(Slab simulation)的粗粒化方法
该模拟方法将所有分子放在长条形的周期性盒子(三个方向边长:x=y<<z)中,在NPT系综的隐式溶剂分子动力学模拟中,做只沿z轴方向平衡的各向异性压力耦合(pressure coupling)。这样分子模拟中相分离完成后蛋白分子聚集于凝聚相,在z轴的很窄的区域分布。Mittal实验室71开发了用于这一模拟的蛋白质粗粒化方法。蛋白质链的每个残基看作一个粒子,相邻粒子间用弹簧相连。不相邻粒子间包含两种作用势函数:Debye-Hückel静电屏蔽的静电作用和短程的成对势。短程的成对势从Lenard-Jones势改造而来,Mittal实验室建议了两种模型,即基于残基对疏水性调整LJ势的模型(HPS模型)和Kim-Hummer发展的原用于蛋白-蛋白相互作用研究的修正LJ势势阱深度参数的模型(KH模型)。这两个模型的未定参数都通过拟合天然无序蛋白质的回转半径(来自小角X射线散射实验)来得到。随后他们用这个粗粒化动力学模拟方法计算了FUS和LAF-1这两个相分离体系的温度-浓度相图。他们的计算表明FUS的模拟磷酸化突变使最高临界共溶温度降低(即不利于相分离),将FUS序列拷贝延长将增加最高临界共溶温度,LAF全长的最高临界共溶温度比只有IDR时的值高。这些结果都与实验结果相吻合。该实验室还用这个模型计算的临界温度值,同天然无序蛋白质单分子的性质,即天然无序线团到蜷缩球转变温度(Tθ)高度相关,因此他们认为天然无序蛋白自身的性质可以用于预估其相分离发生的条件75。他们也与Fawzi实验室合作,用此模拟方法研究了hnRNPA2低复杂度结构域的单点突变对其相分离条件的影响。他们发现D290V突变体比野生型具有更低的临界共溶温度,相同温度下,此突变体的临界浓度比野生型高8。
6.2.2 格点模型下蒙特卡洛模拟
Pappu实验室专注于该方法的研究,他们开发了LASSI (Lattice simulation of Sticker and Spcacer Interactions)具体实现了格点模型的聚集模拟72。格点模型是更为简化的模型,一般将生物分子的结构域(功能单元)整体作为一个粒子,粒子只能处于离散立方网格的格点中,不同粒子所在格点不能重合,一般假设只有相邻格点粒子有相互作用并且粒子对的相互作用能参数化为一个确定值,体系用蒙特卡洛方法演化采样。Pappu实验室用此模拟方法研究了核仁FIB1-NPM1-RNA三元体系中结构域相互作用模式影响形成分层液滴的机制,其中用一个分枝的结构模拟NPM1分子五聚体的结构25。他们也用此模拟方法做了(SH3)m+ (PRM)n体系中无序连接链对相分离的影响的研究76。模拟结果显示连接链的有效溶剂体积影响该体系聚集态的性质,连接链有效溶剂体积大、主要为较刚性的伸展构象时体系倾向于聚集成凝胶,而相反,连接链以压缩状态为主时体系倾向于聚集成液滴。
7 液液相分离的物理化学性质决定了其在细胞中的功能
7.1 生物分子相分离的生物学功能
生物分子相分离在生命体活动中扮演着重要而多样的功能。表2列举了生物分子相分离在发育(a、b)、神经活动(c、d)、固有免疫(e)、环境压力应激(f、g)、染色质结构调控和DNA修复(h、i)、DNA转录调控(j、k、l)、RNA剪接加工与转运(m、n、o)、核糖体合成(p)、细胞分裂(q、r)、信号转导(s、t)、蛋白质自噬降解(u)、细胞膜内吞(v)等细胞活动的各方面所起的关键作用。相分离的这些生物功能可以总结为以下主要的五个方面,对生物分子相分离生物功能的其它分类方式可见综述14,24,77。
(1)富集。将同种功能分子富集在一起以提高局部浓度,或是将不同种类的分子富集在一起来完成同一件任务,是生物分子相分离的首要功能。通过提高酶及其反应底物的浓度,可以显著提高的催化效率。Strulson等人78发现不依赖蛋白的RNA相分离聚集体中的核酶(Ribozyme),HHL和HHS,催化裂解反应的效率比溶液状态下增加了近70倍。DNA转录调控93、RNA剪接加工7、核糖体合成(核仁中)25等过程需要众多分子组件组装成分子大机器才可以完成,相分离形成的液滴富集、组装、修饰加工这些分子组件,为分子大机器提供了工作场所。另外液态聚集体提供了信号转导过程中需要的蛋白质相互作用网络足够的稳定性,使信号输出需要的相互作用维持足够的时间,来达到对输入信号放大输出的作用。在Nephrin +Nck + N-WASP相分离激发肌动蛋白组装的研究中,发现变化Nck/Nephrin的比例,将改变N-WASP稳定停留的时间,进而影响信号输出,即肌动蛋白的组装79。
(2)屏蔽。在细胞受到环境压力时,为避免细胞内重要生物分子受到破坏损伤,这些分子通过躲避至这些临时的聚集体中,同时一般抑制其自身活性,等到压力解除后重新恢复3,80。mRNA通过相分离液滴载运过程中mRNA的翻译暂时被沉默,也是屏蔽保护的功能8。DNA损伤后,损伤部位的相分离对受损DNA也起到屏蔽保护作用6,81。HP1α相分离形成的液态聚集体专一性的招募异染色质,屏蔽RNA聚合酶和转录因子等蛋白质,沉默异染色质的基因表达92。
(3)开关。细胞需要迅速对外界信号做出反应,在不同状态间进行切换。信号转导10、增强子基因转录4,5、神经细胞突触的兴奋信号传递12都是需要一个判断输入信号,达到阈值后快速高强度激活的过程。相分离的协同和高度动态的性质保证了这些生物开关的实现。Hnisz等人82对增强子体系的相分离过程的模型研究表明参与转录调控的蛋白的数目决定着基因转录状态转变过程的希尔系数,增强子体系通过相分离增加了希尔系数,也就是状态转变的协同性。
(4)定位。液滴通过招募锚定在特定细胞位置的蛋白质来实现液滴的定位,从而实现液滴成d分在细胞内的精确定位。P granule向细胞极性一端迁移,使得生殖细胞所需的RNA和蛋白质向极性一端富集定位2。T细胞信号通路的激活引发下游蛋白向细胞膜的胞内一侧富集定位10。
(5)生力。液滴将表面张力转化为机械力的功能。例如由常染色质的低密度区液滴的表面张力转化的机械力改变了染色质的三维组织结构和应力91。细胞膜胞内一侧的液滴表面张力转化的机械力为细胞膜内吞提供动力99。
认识生命体中某一特定体系的相分离过程一般需要如表2所示研究三类相关的分子。首先是确定推动相分离发生的关键成分分子,这些分子的凝聚为招募功能分子搭起基架,称为基架分子(scaffold);然后是被招募的完成生物功能的功能客体分子(client);第三是聚集体的生成和消失受哪些分子调控,一般是执行转录后修饰的分子。

表2 细胞活动过程中生物分子相分离的重要作用与机制Table 2 Important cases of biomolecular phase separation in cell activities.
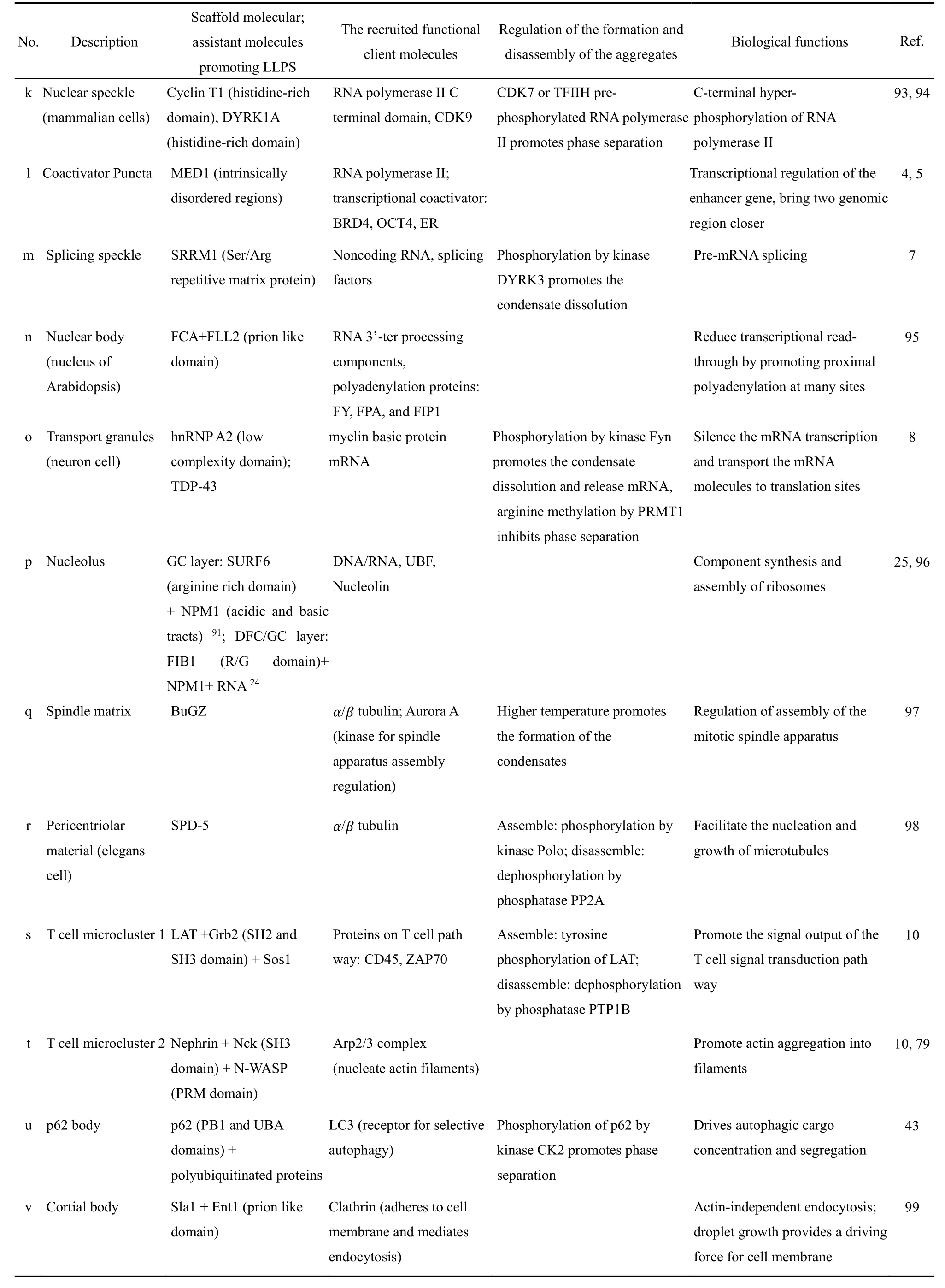
continued Table 2
7.2 生物分子相分离的物理化学性质与生物学功能的关系
图3列举了相分离生物学功能所依赖的物理化学性质。(1)聚集体对酶/底物分子的的专一性招募富集,显著提高了局部聚集体对酶/底物分子的浓度,提升了催化效率。(2)若已经定位的蛋白质可以被液滴招募,那么整个液滴由该蛋白的所在位置而定位。(3)多组分的生物分子在同一时空聚集,同时聚集体是高度动态可逆的,这保证了这些组分完成功能的协同性,起到生物开关的功能。(4)液滴对小分子量分子可以渗透,对非专一性的大分子不可渗透,达到了对液滴外分子屏蔽,对液滴内分子保护的功能。(5)液滴提供拥挤的环境,高粘度低扩散系数稳定了信号转导网络中的蛋白质相互作用。液-液相分离聚集体所提供的拥挤环境在有些体系中会对酶的构象产生影响,从而影响酶的活性,在cGAS聚集体中cGAS被激活;而在进入应激颗粒(stress granules)中mTORC1的活性被抑制。(6)静电作用是多数细胞内液液相分离发生的关键驱动力,因此改变带电状态的翻译后修饰可以灵活高效地调控相分离的发生,这一特点将在下一节中更详细阐述。(7)液滴表面张力驱动机械力的产生。
8 生物分子相分离的调控机制和调控分子设计

图3 生物分子液液相分离的物理化学性质决定了其在细胞内发挥着各种重要的生物学功能Fig.3 The physicochemical properties of biomolecular liquid-liquid phase separation determine that they play various important biological functions in cells.
8.1 生物分子相分离的体内调控机制
生命体体内的生物分子相分离过程受到精细的调控,以适应环境改变、生命活动周期性的变化。体内相分离过程的调控主要包括以下几种可能方式:
(1)通过翻译后修饰改变参与相分离的生物分子的带电性质。最主要的是通过激酶催化的丝氨酸、酪氨酸、苏氨酸的磷酸化,及磷酸酶催化的去磷酸化改变蛋白质的带电状态,调控其聚集或溶解。精氨酸甲基转移酶(PRMTs)将一个或多个S-腺苷甲硫氨酸(AdoMet)上的甲基转移到精氨酸的胍基上,如PRMT调控hnRNPA2或Ddx4的RGG结构域的精氨酸甲基化,实验表明不对称的甲基化显著削弱胍基与芳香环的相互作用从而影响相分离的发生8,47。乙酰化酶/去乙酰化酶催化赖氨酸末端乙酰化或去乙酰化从而改变蛋白的带电状态,如去乙酰化酶HDAC6催化stress granule中DDX3XN端天然无序结构域的乙酰化赖氨酸去乙酰化,从而促进相分离液滴的形成100。多泛素化修饰通过形成与泛素结合蛋白的多价作用,实现凝聚。
(2)通过改变参与相分离的生物分子的浓度。细胞通过激活/抑制基因的表达、改变mRNA可变剪接的方式,以及调整降解速率等改变总体浓度,或是通过定向输运等方式改变局部浓度。
(3)通过调控蛋白质的寡聚状态。对于通过寡聚实现多价作用的体系,可以通过加强或干扰寡聚状态的方式调控其相分离的能力。
(4)对于局部溶液环境敏感的相分离体系,可以通过调整这些溶液环境值影响相分离,如pH值,金属离子浓度,ATP浓度等。例如酵母细胞在饥饿压力下,pH值降低使stress granule生成。ATP作为细胞内的高浓度、高电荷分子,在细胞内充当重要助溶作用,实验表明ATP可以显著地减少FUS的聚集,促进其聚集体的溶解101。
8.2 设计小分子调控生物分子相分离
设计小分子干预调控生物分子相分离过程可以依靠以上调控方式实现,其中最主要的是通过设计相关翻译后修饰酶的调控分子来实现对相分离过程的调控。例如,Wippich等人88通过基于成像的筛选方法得到的抑制stress granule溶解的抑制剂大都是激酶的抑制剂,它们通过调控激酶的活性,来调控相分离基架蛋白的磷酸化状态,从而影响相分离。Aumiller和Keating设计了体外通过激酶/磷酸酶的磷酸化和去磷酸控制多肽可逆地聚集、溶解的体系,他们通过控制ATP的量控制激酶活性,通过控制Mn2+的浓度控制磷酸酶的活性102。相分离调控分子的设计的另外的思路是设计分子直接结合调控蛋白构象、或者影响天然无序蛋白质的构象系综以及多价作用的能力,干扰相分离的发生。1,6-己二醇分子是目前应用广泛的生物分子液-液相分离的非专一性抑制剂103,可能的机制就是影响了天然无序蛋白的构象系综。多聚物poly(ADP-ribose) (PAR)的性质与核酸类似,在细胞内参与调控与核酸相关的相分离过程。Altmeyer等人发现在DNA损伤部位PAR水平显著提高,并促进FUS等含RGG结构域的蛋白质相分离形成聚集体81。McGurk等人发现多聚物PAR结合于TDP-43的PAR结合结构域之后可以促进TDP-43的相分离,在果蝇体内促进TDP-43富集进入stress granule,从而保护该蛋白不发生疾病相关的磷酸化修饰104。Jin等人105揭示了天然无序蛋白质与配体结合的动态特征,Yu等人106发展了针对天然无序蛋白质构象系综进行配体分子设计的方法,为相分离调控分子的设计提供了重要参考。此外设计小分子化合物调控相分离的重要功能客体分子进入聚集体也有重要的价值。例如Fang等人107筛选发现了小分子化合物可以在细胞内阻止TDP-43进入应激颗粒,从而抑制肌萎缩性侧索硬化与额颞部痴呆(ALS/FTD)病变中TDP-43蛋白在神经细胞中的累积。
9 总结与展望
生物分子相分离在生命活动中扮演着富集、屏蔽、开关、定位、生力等重要功能,这些功能与生物分子相分离的协同性、液滴的动态性、专一的招募渗透性、较高密度和粘度等物理化学性质紧密联系的。这些物理化学性质的微观基础是相分离各组分之间的弱而多价的相互作用,以及天然无序蛋白质、无序连接链、以及单链核酸的构象柔性等。序列突变、核磁共振等微观结构测量实验、以及分子模拟等研究手段已经开始揭示生物分子序列、结构与相分离条件和聚集体性质之间的关系,为深入理解生命体怎样通过相分离实现复杂功能提供了定量或定性的数据支持。实现对生物分子相分离过程的精准调控、干预相分离相关的疾病过程24是相分离研究的重要目的,设计翻译后修饰酶的调控分子或是设计构象调控分子是调控生物分子相分离的主要手段。
生物分子相分离过程所展现的普遍性表明当前所发现的生命体内的相分离分子可能只是冰山一角,未来需要从基因组及蛋白质组的层次系统地发掘生命过程中的相分离事件和相关分子,整理出生命系统虚(溶液分散相)-实(聚集相)转换的条件和调控途径。更深入地理解生物分子序列、结构与相分离条件和聚集体性质之间的关系,可以从构建新的模式体系、采用更精确定量的实验方法、以及更准确的分子模拟方法这些方面入手。以可极化力场为代表的新一代生物分子力场108,109提供了更为可靠的静电作用、偶极作用、π-π作用110等这些在相分离体系中最关键的分子间相互作用力的计算方法,有可能为生物分子聚集现象展现出不同于经典力场的图景,比如更为显著的分子协同性。另外利用更准确的力场研究天然无序蛋白质的构象系综,并设计分子改变其构象系综111,从而改变天然无序蛋白质的相分离行为,可能是专一性地直接调控相分离的有效方法。
猜你喜欢
节能技术(2022年4期)2022-11-08
湖北农业科学(2022年11期)2022-07-18
中国慈善家(2022年3期)2022-06-14
现代苏州(2022年9期)2022-05-26
河北农业大学学报(2022年2期)2022-04-26
快乐语文(2021年34期)2022-01-18
实用肿瘤学杂志(2020年4期)2020-12-08
中国(俄文)(2020年8期)2020-11-23
物理化学学报(2020年9期)2020-09-28
生物工程学报(2020年7期)2020-07-29
