
鄱阳湖湿地豆雁(Anser fabalis)越冬种群的春季迁徙路线及其活动特征*
2020-03-27 01:33曹开强李言阔周学红钟毅峰缪泸君
湖泊科学 2020年2期
曹开强,李言阔**,王 强,周学红,钟毅峰,缪泸君
(1:江西师范大学生命科学学院,南昌 330022)(2:中国科学院东北地理与农业生态研究所,中国科学院湿地生态与环境重点实验室,长春 130102)(3:东北林业大学野生动物资源学院,哈尔滨 150040)(4:江西省林业科学院,南昌 330032)
豆雁(Anserfabalis)属雁形目(Anseriformes)鸭科(Anatidae)雁属,是一种大型迁徙水鸟[1]. 长江中下游是豆雁的重要越冬地,近年来鄱阳湖豆雁越冬种群显著增加,2004年冬季记录到5200只个体[2],2007年该区越冬种群数量达13000余只[3],2009年统计数量达28000余只[4]. 鄱阳湖湿地分布有较大的豆雁越冬种群,但有关鄱阳湖湿地豆雁的研究报道极少,主要涉及其种群数量和分布,种群生态学基础研究相对缺乏.
候鸟迁徙行为是鸟类生态学中一个重要的研究领域,国内外学者开展了大量的研究[5-8]. 目前有关豆雁迁徙行为的研究鲜见报道,研究技术落后是造成这种局面的关键原因之一. 近年来,由于卫星跟踪技术具有跟踪范围广、定位精度高、工作时间长等优点,在研究鸟类迁徙方面得到了广泛推广,尤其是国产卫星跟踪设备的成功研发和推广,降低了该技术的使用成本,为研究鸟类迁徙行为提供了有力的技术支撑. 为了揭示鄱阳湖湿地越冬豆雁的迁徙路线和活动特征,我们在鄱阳湖湿地采用国产卫星跟踪技术对豆雁迁徙路线及其活动规律进行了研究,主要分析了:(1)豆雁的迁徙路线;(2)豆雁迁徙前期和迁徙过程中活动量的变化;(3)豆雁迁徙过程中的日活动距离;(4)迁徙期间昼夜活动距离的变化,以及温度对豆雁活动的影响.
1 研究方法
1.1 研究区域
鄱阳湖(28°11′~29°51′N,115°49′~116°46′E)位于江西省北部,是中国第一大淡水湖,上承赣江、抚河、信江、饶河、修水五大河流,下与长江相通. 本研究跟踪的20只豆雁分别来自鄱阳湖南矶湿地国家级自然保护区和都昌候鸟省级保护区(图1). 鄱阳湖南矶湿地国家级自然保护区位于鄱阳湖主湖区南部,是赣江中支和南支汇入鄱阳湖开放区域形成的内陆河湖型三角洲湿地. 都昌候鸟省级自然保护区位于江西省北部,长江南岸,鄱阳湖北岸,地貌以丘陵和滨湖平原为主,且水域宽阔. 两保护区均处于东亚-澳大利亚水鸟迁飞线路,是重要的水鸟越冬地和中继站.
1.2 方法
本研究利用国产卫星跟踪设备对20只豆雁健康个体进行了卫星跟踪定位,豆雁主要来自都昌候鸟省级自然保护区和鄱阳湖南矶湿地国家级自然保护区,卫星跟踪器分为背负式和颈环式(表1). 卫星跟踪器安装时间分别为2019年2月26日、3月5日和3月12日. 跟踪器为湖南信士科技有限公司生产,背负式重量为24 g,颈环式为35 g,均在豆雁体重(平均3.0 kg)的4%以下[9]. 卫星跟踪器采用GPS定位,精度分为A(±5 m)、B(±10 m)、C(±20 m)、D(±100 m)和无效(无法定位)5类定位等级,对卫星数据分析时,A~D级数据可用于豆雁的迁徙研究的数据统计. 数据内容包括活动量、温度、经度、纬度、速度、采集时间等10个字段,各个字段的数据一一对应,其中活动量为设备在一个采集周期内(1小时)所运动的次数. 温度为设备在采集时间记录的温度值(℃). 佩戴在豆雁颈部的颈环式跟踪器由于没有羽毛覆盖,记录到的温度值与环境温度值一致. 本文在探究温度对活动距离的影响时,对相邻两个采集时间点上的温度值取算术平均数,作为这段时间内的迁徙温度. 根据在线(http://richurimo.51240.com)查询的2-5月天亮及天黑的时间,选取每个月天亮及天黑时间的中值作为该月日间和夜间的划分依据[10]. 根据迁徙状态,将豆雁自释放开始至迁飞出江西省定义为迁徙前期,北迁离开江西省为迁徙期. 采用迁徙的日期变化分析法来确定迁徙状态改变的“日期阈值”[11],即某一时刻经度、纬度及活动距离发生明显变化,为迁徙状态变化的临界点. 由于迁徙期间经纬度变化较大,根据迁徙状态将迁徙期划分为飞行状态和停歇状态.
利用ArcGIS 10.2软件中Tracking Analyst Tools中查询定义停歇状态(“speed”≤10),生成停歇地和迁徙路线图,并筛选出迁徙前后日间和夜间所有的点. 在SPSS 20.2软件中,利用独立样本T检验的方法,对迁徙前期活动量和迁徙期活动量差异的显著性进行检验,并使用Pearson相关性来了解温度与豆雁迁徙期活动距离之间的相关性.

图1 佩戴跟踪器的豆雁释放地点Fig.1 The release sites of Bean goose tagged with transmitters
2 结果
2.1 卫星跟踪数据概况
目标豆雁迁徙最早的个体在3月12日开始北迁,较晚的个体4月25日开始北迁,截至6月20日仍有部分个体没有北迁(表2);已迁徙的豆雁中有10只在3月中下旬迁徙,6只在4月初迁徙,而被救助的2只直到4月下旬才迁徙. 追踪距离为1473.3±1035.0 km(n=19),追踪时间为60.5±15.6天(n=19),迁徙距离为1349.5±1004.9 km(n=18),迁徙时间为34.7±16.1天(n=18),停歇地的数量为6±3(n=18). 除了MTT072个体仍停留在鄱阳湖尚未迁徙,MTT046、MTT094和MTT098在迁徙途中部分数据出现异常,MTT029在释放两天后异常未获得有效数据. 豆雁迁徙开始的时间大部分在上午. 迁徙距离占追踪距离的比为77.2%±21.5%,n=18),迁徙时间占追踪时间的比为56.8%±19.5%,n=18).
2.2 迁徙路线
豆雁在国内的春季迁徙路线如图1所示,即已到达东北三省和内蒙古一带的豆雁共有9只,分别为JXNU11、JXNU13、JXNU14、JXNU15、JXNU17、JXNU18、MTT011、MTT013、MTT098. 春季迁徙路线可以分为东线和中线两部分,东线经过我国安徽、江苏、山东、辽宁、吉林、黑龙江,进入俄罗斯境内;中线经过我国湖北、安徽、河南、河北、内蒙古,进入蒙古境内. 对比两条迁徙路线的地形,沿着中线迁徙的豆雁相较于东线的海拔跨度更大. 中线从鄱阳湖平原出发,经江淮平原、黄淮平原,沿着太行山脉以东、海河平原西部,到达中国第二级地势阶梯内蒙古高原的锡林郭勒草原、呼伦贝尔草原. 东线则沿着海河平原东部,由渤海湾进入辽河平原、松嫩平原和三江平原,均在第三级地势阶梯内.

表1 鄱阳湖佩戴跟踪器的豆雁信息

表2 豆雁卫星跟踪数据概况
“-”代表数据缺失.

图2 鄱阳湖越冬豆雁北迁路线Fig.2 The migration routes of Bean goose of Lake Poyang
2.3 迁徙前期与迁徙期活动量
迁徙前期共获得7736个定位点,迁徙期共10034个定位点. 豆雁迁徙前期活动量为1081.1±857.8次/h(n=7736),迁徙期活动量为916.7±828.4次/h(n=10034),迁徙前后的活动量存在差异显著(t=-12.910,df=17768,P=0.000). 豆雁迁徙前期日间活动量为1245.2±877.3次/h(n=4575),迁徙前期夜间活动量为843.6±769.3次/h(n=3161),迁徙前期昼夜活动量存在差异显著(t=20.798,df=7734,P=0.000). 迁徙期日间活动量为1154.5±855.6次/h(n=5874),迁徙期夜间活动量581.0±655.0次/h(n=4160),迁徙期昼夜活动量差异显著(t=-36.246,df=10032,P=0.000). 迁徙前期日间活动量与迁徙期日间活动量差异显著(t=5.313,df=10447,P=0.013),迁徙前期夜间活动量与迁徙期夜间活动量差异显著(t=15.750,df=7319,P=0.000),豆雁的活动具有明显的昼夜变化. 迁徙前期日间在8:00-9:00和18:00-19:00这两个时段为活动高峰,其中18:00-19:00时段活动量最大,为1358.9±864.5次/h(n=319),其次是8:00-9:00时段,为1243.7±872.1次/h(n=312);迁徙期日间在7:00-8:00和18:00-19:00这两个时段活动高峰,其中18:00-19:00时段活动量最大,为1431.0±918.1次/h(n=421),其次是7:00-8:00 时段,为1232.8±919.4次/h(n=420)(图3).
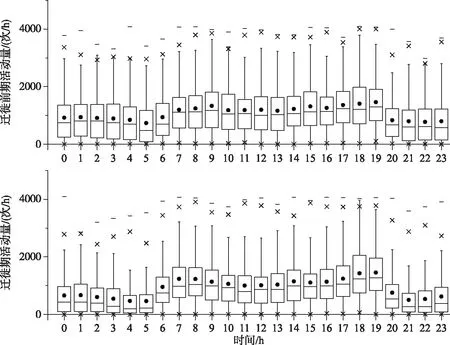
图3 豆雁迁徙前期和迁徙期的日活动量Fig.3 Daily activity of pre-migration and migration period of Bean goose
2.4 停歇地日活动距离
具有完整国内春季迁徙路线的9只豆雁纬度跨度在28°6′~50°7′N之间,我们制作了纬度和时间关系图,水平线表示停留,垂直线表示向北迁飞(图4). 成功迁徙的豆雁从越冬地鄱阳湖(28°87′~29°19′N)迁出后,在30°~40°N之间较少做长时间的停留,利用时间为1.6±1.1天(n=9),而在40°~50°N之间利用时间为35.6±13.7天(n=9),两个区间利用时长存在显著差异(F=10.969,df=16,P=0.000). 在40°~50°N之间平均每只豆雁会做2.7±1.1次停歇(n=9),每次停歇长达12.7±7.2天(n=24),在30°~40°N间平均每只豆雁会做3.0±1.7次停歇(n=9),两个区间豆雁停歇次数差异不显著(F=1.306,df=16,P=0.634).
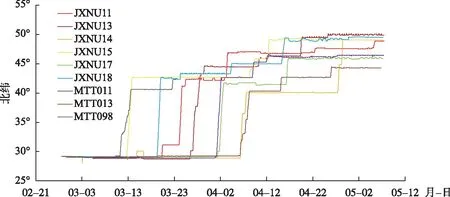
图4 豆雁春季迁徙的时间和纬度Fig.4 Time-latitude analysis of spring migration of Bean goose
筛选出越冬地和40°~50°N之间的中途停歇地的点,计算日活动距离. 共确定除越冬地鄱阳湖外5个省市17处中途停歇地(表3),豆雁中线迁徙种群共有5处停歇地,1处位于海河平原,1处位于内蒙古高原南缘,1处位于锡林郭勒草原,2处位于呼伦贝尔草原;东线迁徙的种群共有12处停歇地,1处位于滦河三角洲,1处位于渤海湾,1处位于辽河平原,4处位于松嫩平原,4处位于科尔沁草原,1处位于三江平原. 停留时间最长的停歇地是在黑龙江,为25.7天,日活动距离最长的停歇地是在额尔古纳河,为98.18 km,卧龙湖和洋沙泡是豆雁利用最多的中途停歇地,数量均为3只. 17处中途停歇地豆雁日间活动距离为28.3±18.7 km(n=17),夜间活动距离为11.0±10.7 km(n=17),豆雁的昼夜活动距离存在显著差异(F=5.650,df=32,P=0.002).
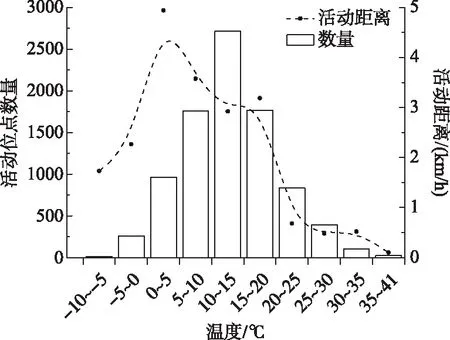
图5 温度分布与活动距离及活动位点数量Fig.5 Temperature distribution and active distance, active site numbers
2.5 温度与活动距离
具有国内完整迁徙路线的6个佩戴颈环的个体共收集8827个定位点,这些活动点的温度表现出正态分布(图5),温度在10~15℃的位点占总位点数的30.7%,15~20℃的位点占比为20.0%,5~10℃的位点占比为20.0%,0~5℃占比为10.9%,20~25℃占比为9.5%,25~30℃占比为4.4%,-5~0℃占比为2.9%,30~35℃占比为1.1%,35~41℃占比为0.3%,-10~-5℃占比为0.2%. 由于各温度区间的活动距离方差不齐,所以利用非参数Kruskal-Wallis 检验的方法,结果各温度区间的活动距离存在显著差异(n=8827,df=9,P=0.000),在0~5℃时的平均活动距离最大,为4.9±14.4 km/h;35~41℃时平均活动距离最小,为0.1±0.2 km/h;-10~-5℃ 的平均活动距离为1.7±4.5 km/h(图5). 温度与活动距离呈显著相关(r= -0.061,n=8827,P=0.000),当温度在35~41℃时,温度与活动距离呈显著负相关(r=-0.405,n=23,P=0.05).
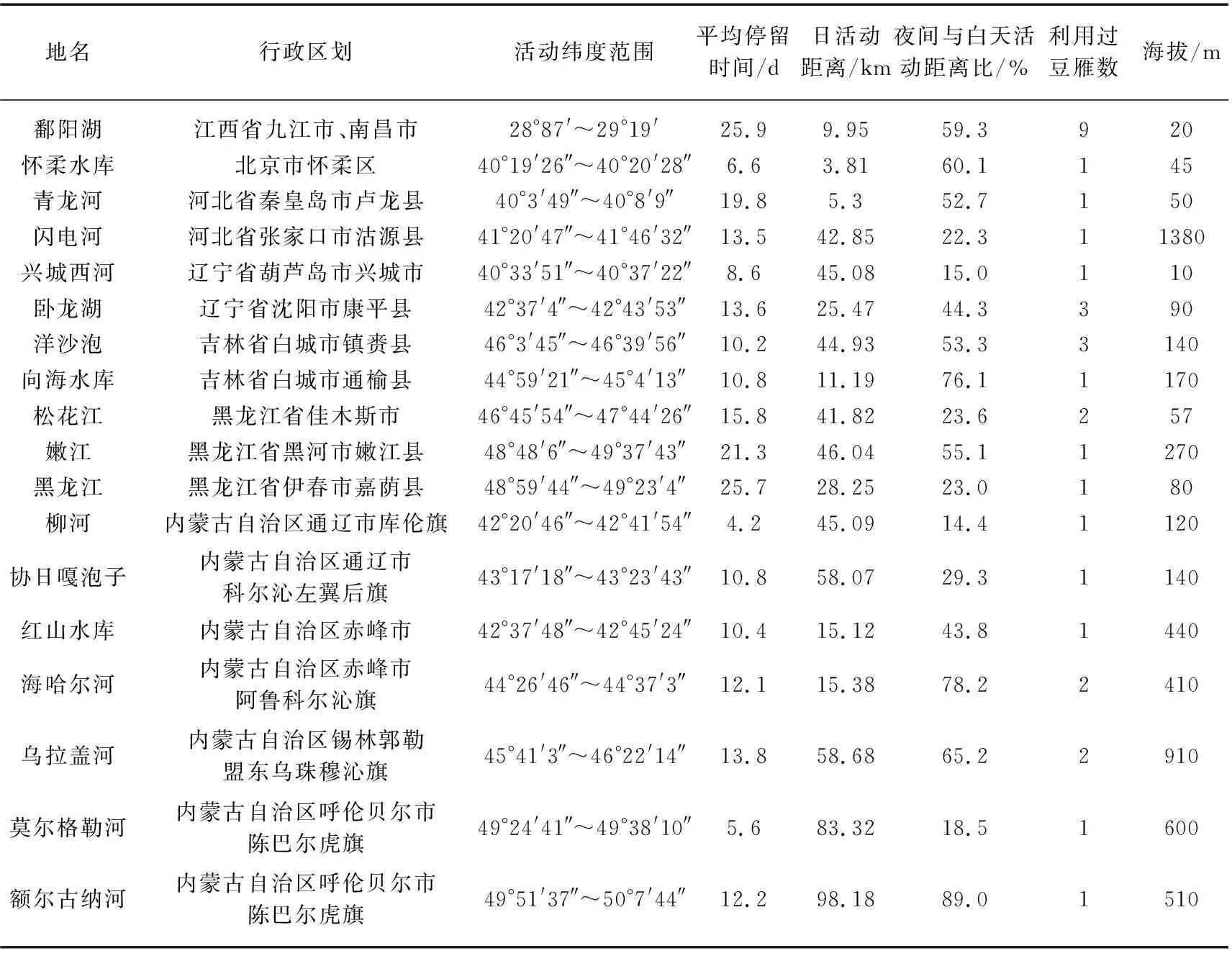
表3 豆雁国内迁徙期间的停歇地
3 讨论与结论
3.1 卫星跟踪对豆雁的影响
对比两种类型卫星跟踪器的效果来看,颈环式跟踪器具有明显的优势,7只佩戴颈环的豆雁均成功迁徙,并且跟踪器在持续工作无异常,而背负式跟踪器13只豆雁中有4只个体出现异常,6只个体成功迁徙,2只个体在迁徙途中停歇地做长时间的停留,1只个体未迁徙仍停留在鄱阳湖. 从佩戴方式上考虑,背负式跟踪器由于绑在豆雁背部并且绳子需要绕过双翅可能在一定程度上限制了豆雁的飞行;而且背负式相较于颈环式更加的繁琐,需要更长的时间,这样对其造成伤害的可能性也会增加. 所以在利用卫星跟踪技术研究雁鸭类时,颈环式跟踪器是更为合适的选择.
本研究中追踪的20只豆雁,有16只在2月份安装并释放,11只已成功迁徙到内蒙古和东北三省的边境地区,而3月份安装的4只个体,只有1只成功迁徙,3只出现异常. 从豆雁迁徙开始时间来看,3月中下旬和4月初为正常的迁徙时间,而在3月12日释放的3只豆雁(JXNU16、MTT035和MTT090),由于受伤被救助的时间较晚短时间内来不及完成能量储备直至4月下旬才开始迁徙,过晚的迁徙在没有合适迁徙种群的情况下迁徙成功率可能较低. 安装卫星跟踪器应选择3月份之前,这样可以减小对豆雁迁徙活动的影响,从而提高卫星跟踪的成功率.
3.2 迁徙路线与停歇地
季节性迁徙可以理解为一种维持繁殖地资源同时避免该地资源季节性枯竭的长期生存策略[12-14]. 本研究发现豆雁在国内的迁徙路线可分为东线和中线两部分,在30°~40°N之间东线和中线均从鄱阳湖平原出发,经江淮平原、黄淮平原和海河平原,中线最西侧沿着太行山脉以东,东线东侧为山东丘陵和渤海湾. 中线和东线在40°N左右开始区分,中线迁徙种群进入内蒙古高原东部,东线迁徙种群进入东北平原地区. 大兴安岭山脉是这中、东两线的天然屏障. 东线经过我国安徽、江苏、山东、辽宁、吉林、黑龙江,进入俄罗斯,与郭家良和Si等报道的迁徙路线基本一致[15-16]. 中线经过我国湖北、安徽、河南、河北、内蒙古,进入蒙古,与Wang得出的迁徙路线有部分重叠[17]. 张孚允等分析了我国鸟类环志资料后将我国分3个主要候鸟迁飞区:东部候鸟迁徙区,包括东北地区、华北东部平原、丘陵和滨海地区;中部候鸟迁徙区,包括内蒙古东部、中部草原,华北西部地区以及陕西地区;西部候鸟迁徙区,包括在内蒙古西部干旱草原,甘肃、青海、宁夏等地的干旱或荒漠、半荒漠草原地带和高原草甸草原地区[18]. 本研究中豆雁的迁徙路线在一定程度上支持这一结论.
迁徙性动物依靠迁徙期间成百上千个运动决策实现迁徙以及对时机的把握[19]. 迁徙路线和迁徙策略具有很强的适应性,并且中途停歇地在繁殖地和越冬地之间提供重要的连接[20]. 本研究发现迁徙豆雁对30°~40°N和40°~50°N两个纬度区间的利用时间存在显著差异,但是停歇次数差异不显著. 这些停歇地可以分为三类:第一类包括湖北、安徽、江苏和山东等长江中下游平原地区与华北平原地区,如龙感湖、升金湖、吉利河或者人工库塘等湿地,是具有随机选择性的临时停歇地,供候鸟临时休息或供候鸟在恶劣气候下进行临时停歇,停歇时间较短[21];第二类包括北京、天津和河北等华北地区向东北地区和内蒙古地区的过渡地带,如怀柔水库、青龙河等湿地,是飞越高山、海洋等生态屏障前的停歇地. 候鸟在飞越生态屏障前,常需要在这些地区的高质量停歇地储备大量的能量[22];第三类包括被大兴安岭分隔开的内蒙古东部地区和东北地区,如锡林郭勒草原、呼伦贝尔草原、辽河平原、松嫩平原和三江平原等地区湿地,是候鸟传统的补给地,候鸟对这些停歇地的忠诚度较高,在此停留时间较长,能够积蓄能量为鸟类继续飞行和繁殖提供保障[23]. 豆雁对临时停歇地选择的随机性,以及对传统补给地高忠诚度形成了迁徙路线前期的分散和后期在重要停歇地的高重复率[24].
3.3 迁徙前后的活动量变化
活动量测度是个体在一个采集周期内(一般为1 h)活动的次数,活动量大表明个体可能与迁飞有关,也可能是个体在停歇地开展大量的觅食或其它活动. 豆雁迁徙前期和迁徙期的活动量都具有明显的昼夜变化,白天的活动水平高于夜间,这表明豆雁是昼行性动物,这一习性与其它大多数雁鸭类物种一致[25]. 迁徙前期日间活动高峰为8:00-9:00和17:00-18:00,而迁徙期活动高峰为7:00-8:00和17:00-18:00,迁徙期活动高峰较迁徙前期提前一个小时,可能是因为北半球春分日到夏至日昼渐长夜渐短,纬度越高的地区天亮的时间越早,光照强度达到一定水平豆雁即开始活动[26-28]. 迁徙前期和迁徙期白天活动量都反映出早上和下午较晚时分较高,中午较低,呈现出一个双峰值曲线,这与刘静[29]观察到的日觅食曲线规律基本一致. 刘静[29]发现豆雁越冬后期的觅食行为比例为64%,其中3月份的觅食比例上升到74%,越冬期间尤其是越冬后期觅食行为是豆雁日间活动的主要行为,而迁徙前期为了满足北迁的能量需要豆雁日平均觅食水平明显增加. 在这种情况下,活动量可以有效地反映出豆雁日间觅食行为的变化. 由于自身条件决定了雁群不可能在夜间觅食,同时3-4月份的东北、内蒙地区夜间比鄱阳湖地区夜间更加寒冷,为了减少能量消耗雁群在迁徙期夜间活动量较迁徙前期显著减少.
3.4 迁徙期间温度分布与活动距离
成功迁徙的豆雁中有6只为颈环式跟踪器. 置于豆雁颈部的跟踪器,在没有羽毛覆盖的情况下,设备采集到的温度值应与环境温度值一致. 迁徙期间豆雁所处环境极端温度(-10~-5℃和35~41℃)只占0.5%,而中性温度5~20℃占70.7%,各温度区间的活动距离差异显著,这可以理解为迁徙性鸟类对温度的一种适应策略,即通过选择温度适宜的时间、避开不利温度的空间共同作用的方式. 豆雁作为恒温动物新陈代谢旺盛,羽毛致密且皮肤无汗腺. 当环境温度过高时,温度与豆雁的活动距离存在显著的负相关,以豆雁喜冷怕热的习性,而且飞行时新陈代谢加快需要消耗大量能量的同时产生大量热量,所以在35~41℃的外界高温情况下,豆雁的平均活动距离只有0.1 km/h,在几乎不移动的情况下通过腿部与环境(水)散热[30]. 而在0~5℃的平均活动距离最大达4.9 km/h,说明这区间的温度最适合雁类迁徙飞行. 低温环境下(-10~-5℃)大部分水域处于冰冻状态,不利于豆雁栖息而选择飞行寻找合适的栖息地点.
猜你喜欢
水利水电快报(2022年7期)2022-07-18
太阳能(2022年3期)2022-03-29
中国畜牧杂志(2021年8期)2021-08-15
太阳能(2020年3期)2020-04-08
现代出版(2019年6期)2020-01-14
保健与生活(2019年15期)2019-09-12
当代工人·精品C(2019年2期)2019-05-10
神州·上旬刊(2017年11期)2017-12-04
计算机应用与软件(2017年7期)2017-08-12
考试周刊(2016年66期)2016-09-22
