
福建省两种林分类型物种结构特征
2020-03-25 03:09:52叶宝鉴陈永滨曾钦朦游水生陈世品
江西农业大学学报 2020年1期
叶宝鉴,陈永滨,马 良,曾钦朦,游水生,陈世品 *,陈 辉
(1.福建农林大学 林学院,福建 福州 350002;2.福建农林大学 森林与环境学报,福建 福州 350002;3.福建生物工程职业技术学院,福建 福州 350002;4.福建农林大学 园林学院,福建 福州 350002)
【研究意义】常绿阔叶林具有良好的生态效益,其评估价值是人工纯林的10倍,是草地的20~30倍[1],主要表现在保水保肥效益[2-3]、光合作用高效[4]、物种组成复杂[5]等。良好自然生态环境可为人类提供包括水在内的各种不可或缺的生态战略资源,是人类生存和可持续发展的重要保障。据我国第8次森林资源清查结果表明,我国现有森林面积达2.08亿hm2,其中生态公益林占56%[6-7],但许多生态公益林为人工纯林,树种结构单一,抵御病虫害、飓风、泥石流等自然灾害的能力差[8],影响了其群落稳定及生态功能的发挥。保护意识的薄弱,人工林的快速发展,导致部分常绿阔叶林林分类型的改变,遭受到严重的破坏,森林生态效益被极大限制[9-10]。加之近年大面积的马尾松(Pinus massoniana)、桉树(Eucalyptussp.)人工林遭受毁灭性病害[11-12]启发了林业经营者新的森林经营理念,林分的稳定优于短期的经济利益。常绿阔叶林是亚热带地带性植被的稳定林分,如何将现有功能单一的人工林向具有多功能的常绿阔叶林转变日益为人所关注[13-14]。【前人研究进展】功能单一的人工林向功能多样的常绿阔叶林改造的技术理论尚处于萌芽阶段,该研究已成为恢复生态学研究热点之一[15]。已有研究表明,森林多功能的发挥在很大程度上取决于其物种结构是否合理[16],抚育间伐、封山育林、混交林营造等[3,17-18]是改变林分群落结构的主要措施,但由于种源匮乏、土壤退化、恢复时间过长、经营技术低下等原因,导致恢复效果仍然不甚理想[19-20]。【本研究切入点】仿生栽培是指模仿生物自然规律栽培植物的方法,该方法已在农作物[21]、中草药[22]、食用菌[23]等领域大量推广。近自然林业经营理念近年也为人们所推崇[24],但对于人工林如何依据地带性常绿阔叶林物种结构进行人工林改造的研究还比较匮乏。【拟解决的关键问题】本文拟通过米槠常绿阔叶林(Castanopsis carlessiforest,CCF)及其周边的杉木林群落(Cunninghamia lanceolataforest,CLF)结构特征的比较,理清二林分之间的差异,掌握待恢复林分中需要增加的物种,进而为后期的苗木储备、林分改造提供依据。
1 研究区概况
福州北山位于福州北郊,纬度26°03′~26°40′N,经度119°18′~119°44′E。全年冬短夏长,温暖湿润,无霜期达326 d,年均日照数为1 848 h,太阳辐射104.6 kcal/cm2;年均降水量为1 740.2 mm,总雨日115~175 d;年均气温为17℃,最冷月为1月,平均气温5.9℃;最热月为7月,平均气温为24℃。极端气温最高42.3℃,最低-4℃。年相对湿度约77%。福州北山属海洋性亚热带季风气候,夏季盛行东南风和南风,冬季多为西北风和北风,7—9月是台风活动期,每年直接登陆市境的台风平均约2次。植被有常绿阔叶林、人工林等林分类型,本文选取米槠常绿阔叶林(米槠林)、杉木人工林(杉木林)为研究对象,其中杉木人工林于1990年造林,现已划为省级生态公益林,保护较好。
2 研究方法
2.1 样地建立与调查
根据最小面积理论,福建省中亚热带常绿阔叶林最小面积为1 200 m2[25],因而本文在鼓山涌泉寺米槠林、宦溪林场杉木林各设面积1 600 m2永久性固定样地,并划为64个5 m×5 m样方,对林分进行样方调查,以全面地反映群落特征。两种 分样地坡向均为西南坡,其中米槠林样地坡度为15°,杉木林坡度为20°。统计物种组成、Jaccard相似系数、木本植物高阶分布,群落多样性等群落特征。
2.2 数据分析方法
Jaccard群落相似系数:

式中a为在群落1出现的物种数,b为在群落2出现的物种数,j为群落1和群落2共有的物种数。
Margalef物种丰富度指数:

Shannon-Wiener多样性指:

Simpson生态优势度:

Pielou均匀度指数:

以上公式中,S为物种数,ni为第i个物种的个体数,N为样方物种总数,pi为第i个物种的个体数占物种总个体数的比例。
根据植株高度划分高阶,共划分为5个高阶,即A级:0~2 m;B级:2~5 m;C级:5~10 m、D级:10~15 m;E级:≥15 m,其中0~5 m为灌木,5 m以上为乔木。采用Microsoft Excel 2007等软件进行数据处理、分析及绘图。
3 研究结果
3.1 物种组成
通过样方调查,结果表明米槠林有维管束植物51科89属115种,杉木林有32科48属54种(表1),两种林分类型均以双子叶植物为主体,裸子植物种类较少。杉木林物种数约为米槠林一半,其中米槠林蕨类植物科属种数均低于杉木林;米槠林裸子植物科数低于杉木林,属数、种数相等;米槠林双子叶植物、单子叶植物科属种数均大于杉木林。
3.2 Jaccard相似系数分析
Jaccard相似系数分析表明两种林分单子叶植物的科相似度最高(图1),为0.75。蕨类植物、裸子植物相似度值都为0.5,双子叶植物科相似度为0.39。属相似度统计表明蕨类植物科属种Jaccard相似系数分别为0.50、0.36、0.25;裸子植物科属种相似系数分别为0.50、0.33、0.33;双子叶植物科属种相似系数较低,分别为0.39、0.23、0.16;单子叶植物科属种数相似系数分别为0.75、0.22、0.23。双子叶植物是构成群落乔木、灌木的主要成分,而其在2种群落中的物种相似系数最低,决定了2群落生态功能的发挥。
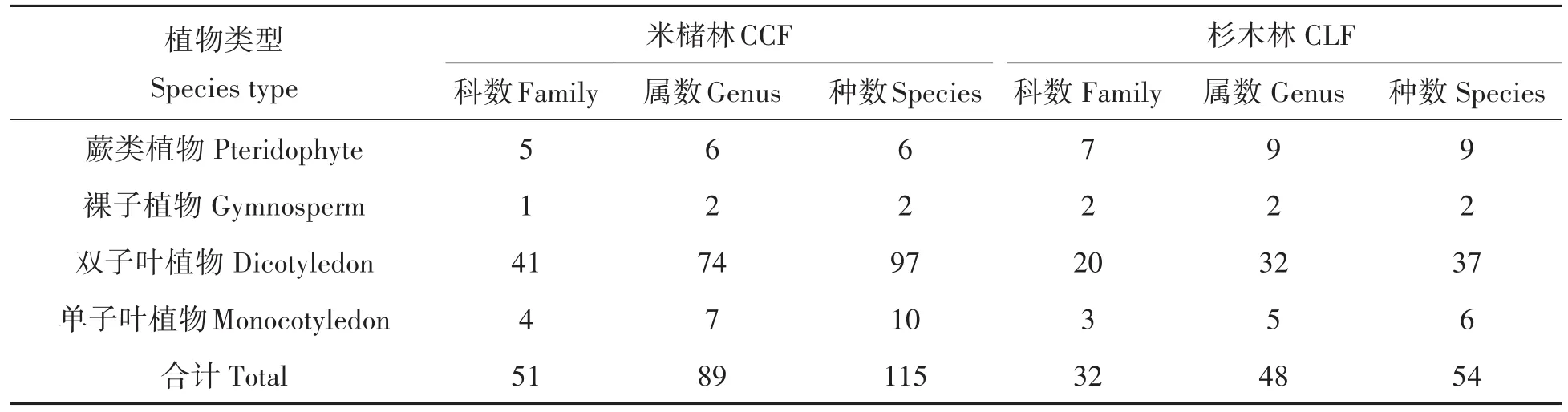
表1 米槠林与杉木林物种组成比较Tab.1 Species composition in CCF and CLF
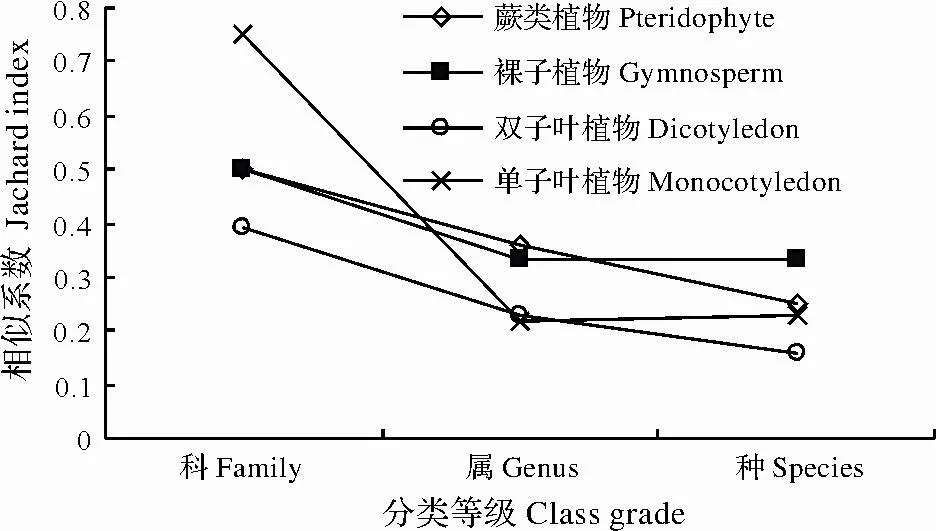
图1 两种林分科属种Jaccard相似度Fig.1 Jaccard analysis in the scale of family,genus,species in two forests
3.3 木本植物高度径阶分布
木本物种高度径阶分布表明天然林木本植物共5 511株,杉木林1 625株,天然林木本植物株数是杉木林的3倍以上,呈倒J型分布(图2)。天然林在A、B径阶有分别有4 394株、839株,大于杉木林株数,其中A径阶约为杉木林的7倍,C、D、E径阶杉木林株数大于天然林,天然林灌木数量庞大,能够很好地保证林分的可持续发展。杉木林灌木数量虽然大于乔木,但比例不够合理,存在许多丛生杉木,随着苗木的生长将加剧种内竞争,导致该径阶苗木大量死亡。杉木林乔木数量高于天然林数量,主要原因是人工林密植导致的。
3.4 木本植物物种径阶分布
木本植物物种径阶表明(图3),米槠林、杉木林内物种随着高度径阶的升高而降低。米槠林在A径阶物种数量最大,为74种,B径阶有52种,C径阶29种,D、E径阶分别有11种、4种,样地灌木层有77种,乔木层31种。杉木林A径阶有23种,B径阶13种,C、D、E径阶均为3种,样方内灌木层有24种,乔木层有4种。米槠林各径阶物种数及灌木株数均高于杉木林,表明米槠林物种库较为丰富,结构合理。
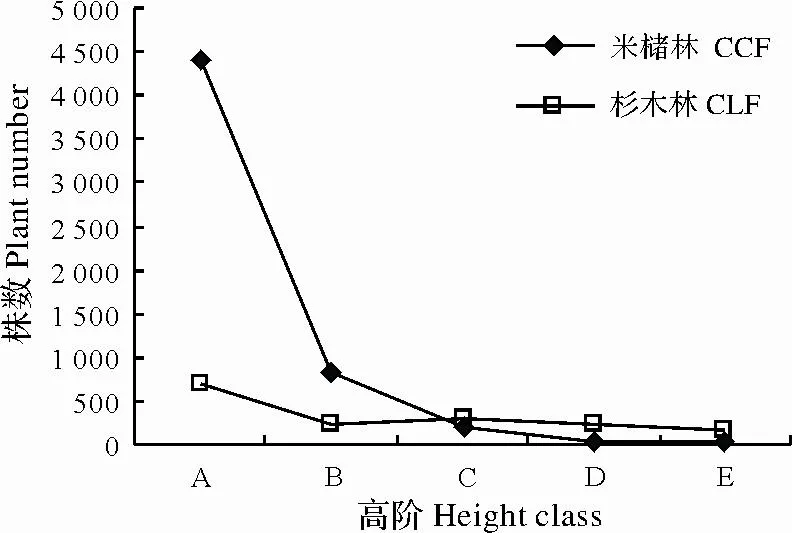
图2 两种林分不同径阶株数统计Fig.2 Static of plant number at different classed in C.castanopisis and plantation forest
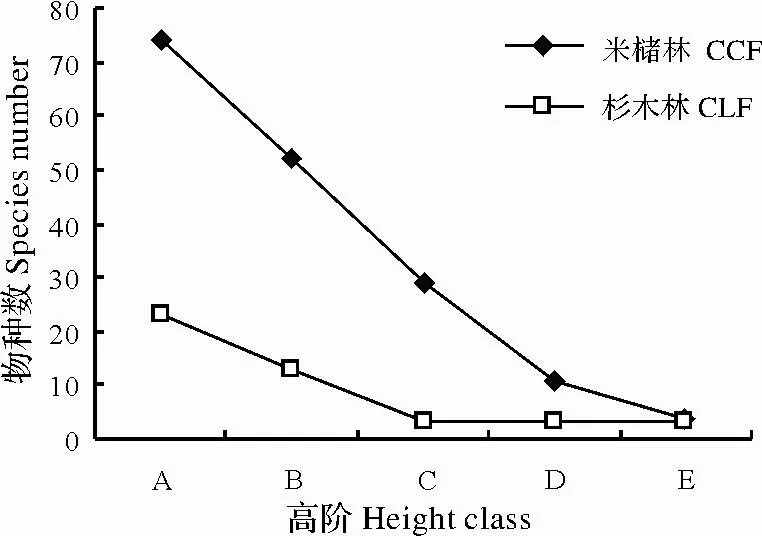
图3 两种林分不同径阶物种数统计Fig.3 Static of species number at different classes in C.castanopisis and plantation forest
3.5 群落多样性
群落生物多样性计测结果表明,米槠林与杉木林群落Shannon-Wiener指数H、Simpson指数D、Pielou指数J差异不大,杉木林Margalef指数与米槠林有较大差异(表2)。杉木林乔木层物种单一,主要以杉木为主,因而乔木层物种R、H、J都较小,而D则显著大于天然林。米槠林灌木层R、H、J均大于杉木林,D小于杉木林,表明米槠林灌木层多样性优于杉木林。米槠林层间层R、H、D、J与杉木林大体一致。天然林草本层R与杉木林一致;但H、J小于杉木林,D大于杉木林,表明米槠林下草本植物物种与杉木林相当,但是有部分物种占优势地位,如华山姜(Alpinia chinensis)分布,而杉木林成林时间较短,尚处于演替前期,阳性植物占优势,阴性植物也已进入林分,开始展开激烈竞争,因而物种分布较为均匀。米槠林乔木层、灌木层物种多样性几个测度表明,米槠林乔木层、灌木层物种多样性较杉木林丰富,杉木林生态优势度大于米槠林。
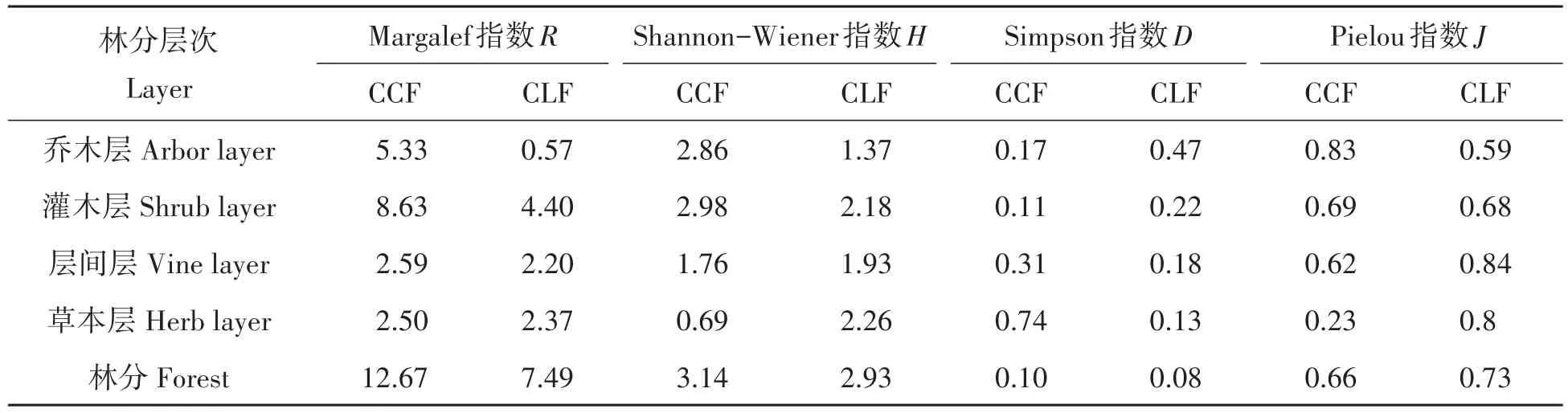
表2 米槠林、杉木林群落多样性Tab.2 Community biodiversity in C.castanopisis and plantation forest
4 讨 论
本文通过两种林分类型物种结构特征的比较表明,米槠林物种多样性优势主要表现在双子叶植物层次,米槠林、杉木林内蕨类植物、裸子植物、单子叶植物在种的数量上没有显著差异。杉木林在长期的经营过程中,由于蕨类植物、单子叶植物对已经郁闭的林分生长不形成阻碍,是其能够在杉木林内存在的主要原因,其特殊的繁殖方式也是其存在林分的一大因素。非目的树种在森林经营中被大量去除,现存在林分中的也多呈灌木状,从而表现出杉木林中等径阶物种及株数的缺失。米槠林物种种类数量与《福建植被》中亚热带常绿阔叶林物种种类描述基本吻合[26]。杉木林物种数量较前人研究略有增加,这可能是由于亚热带优越的水热条件与周围较为丰富的种质资源,导致杉木林内物种演替的发生[27]。
树木是森林的主体,是森林发挥生态效益的主要载体,分析林分木本植物组成能够更好的解释林分生态效益差异的原因。结果表明,物种径阶分布表明二林分灌木、低径阶乔木差异显著,米槠林木本数量较多,尤其是灌木,许多灌木都是发展成为乔木的储备,因而是稳定的林分;杉木林灌木数量少,物种分布不合理,难以长期存在。物种径阶分布表明,米槠林物种径阶分布合理,A、B径阶物种丰富,C、D、E径阶物种分布均匀,杉木林A、B物种较少,C、D、E径阶物种缺乏变化,即杉木、湿地松(Pinus elliottii)及木荷(Schima superba)或樟树(Cinnamomum camphora)。二林分D、E径阶都较少,表明林分是少数种占优势的林分类型。天然林物种逐级降低,表明天然林有很好的多级结构,层次分明。刘万德等[28]对森林群落特征的研究中也支持许多低径阶物种种类较多,数量较大的观点。灌木、低径阶是高径阶乔木的必经阶段,在条件成熟的条件下,部分灌木、低径阶乔木可以往高径阶乔木发展。
物种多样性是森林生态服务功能的指示指标之一,物种多样性高则所在群落的生态功能通常较高[29-30]。群落多样性计测结果表明,米槠林群落多样性显著优于杉木林,尤其在乔木、灌木层次,与何汉杏等人研究结果大抵一致[31]。杉木林草本层H高于天然林,但米槠林内R高于杉木林,其次随着演替的发生,杉木林草本层部分阳性物种将逐渐消失,如野茼蒿(Crassocephalum crepidioides)、三脉紫菀(Aster ageratoides)等,二林分类型在草本层基因库层面差异较小。因而,米槠林的生态服务功能优于杉木林,对杉木林进行林分改造有助于其生态服务功能的提升。
相似度结果表明两种林分科属级别物种相似度较高,种相似较低。考虑属级别物种的生活型有较大程度的相似性,两种林分属级别物种相似度高则林分功能类型相近。双子叶植物种的相似度较低,乔木层、灌木层H差异较大,草本层、层间层H差异较小,表明林分中乔木层、灌木层相似系数较低。乔木层,灌木层生态效益是整个林分中的主体部分[32-33],林分中乔木层、灌木层物种相似度较低,因此认为两种林分生态效益的差异主要取决于木本植物的差异上。
封山育林是一种很好的林分修复方式,只要时间足够长就有可能恢复到顶级群落状态,至少能够恢复到相当水平。人为引入种源并进行合理配置,能够快速恢复到常绿阔叶林水平。常绿阔叶林地处亚热带,生态位物种共存理论能够很好的指导于类天然林的工作[34-35]。因此,人工林改造为常绿阔叶林恢复,应引入具有不同生态位的物种。实现人工林转变,主要应进行乔木层、灌木层树种恢复。通过比较两种林分物种结构,得出待恢复树种层次、种类,为人工选择物种种源、培育苗木提供依据。根据植被调查结果、生态位共存理论,进行种源合理配置,科学地改造现有人工林,不断丰富林内物种,使人工林在物种、结构及功能上不断向地带性植被方向发展,最终发展成常绿阔叶林。
本文仅从物种结构对2种林分特征作出比较,尚未对各物种重要值、空间分布、种群结构特征等群落特征做出分析,还有待进一步研究。
5 结论
天然林是一个地区最适宜的顶级群落,具有相当复杂的群落结构,因而具有很高的多样性。杉木林在林分抚育管理过程中,排除了大量非目的物种,林分进展缺乏动力,最终导致米槠林与杉木林乔木层、灌木层物种存在差异。通过群落学本底调查,分析两种林分物种结构、木本植物数量、物种径阶分布、群落多样性、Jaccard相似度的差异。结果表明林分结构差异显著,米槠林群落多样性优于杉木林,尤其在乔木(H=2.86)、灌木层次(H=2.98),而二林分草本层、层间层差异相对较小。缺乏低径阶乔木、灌木物种及数量是杉木林生态服务功能的限制性因素,现有人工林改造首要恢复的物种应该是乔木层、灌木层木本植物。
猜你喜欢
山西林业(2021年2期)2021-07-21 07:29:28
农村百事通(2020年21期)2020-11-23 02:04:24
安徽农学通报(2020年17期)2020-10-09 10:46:23
绿色中国(2019年18期)2020-01-04 01:57:08
绿色中国(2019年19期)2019-11-26 07:13:16
绿色中国(2019年19期)2019-11-26 07:13:16
农家科技下旬刊(2017年8期)2017-11-13 18:09:22
防护林科技(2016年8期)2016-08-29 01:28:17
贵州科学(2016年2期)2016-06-13 00:58:58
华南农业大学学报(2016年3期)2016-06-01 09:27:59
