
基于微卫星亲权鉴定技术的短蛸性选择研究*
2020-03-25 06:04范婷婷冯艳微刘文芬王卫军杨建敏
渔业科学进展 2020年2期
范婷婷 冯艳微 刘文芬 王卫军 杨建敏
基于微卫星亲权鉴定技术的短蛸性选择研究*
范婷婷1,2冯艳微2①刘文芬3王卫军4杨建敏4①
(1. 上海海洋大学 水产科学国家级实验教学示范中心 上海 201306; 2. 山东省海洋资源与环境研究院 山东省海洋生态资源重点修复实验室 烟台 264006; 3. 中国水产科学研究院长江水产研究所 农业农村部水产品质量安全风险评估实验室 武汉 430070; 4. 鲁东大学农学院 烟台 264025)
为研究短蛸()的性选择行为,本研究利用14个多态性微卫星标记,对3个母本、176个子代及17个候选父本进行亲权鉴定,对3个家系中有子代和无子代雄蛸、子代比例高和子代比例低的雄蛸的各形态参数进行检验,并对子代比例高和子代比例低的父本与母本的遗传相似性与遗传距离进行分析。结果显示,短蛸为多雌多雄的交配模式,为真正意义的雌性混交动物;雄蛸有无子代与其形态参数无相关性;子代比例与父本的形态大小参数无关,而与父母本的遗传相似性和遗传距离有关,父母本遗传相似性越高,遗传距离越小,后代比例越高。本研究为揭示短蛸交配前后的性选择机制提供了重要线索,也为海洋头足类动物的性选择机制研究提供了基础资料。
短蛸;微卫星标记;亲权鉴定;交配模式;性选择
性选择(Sexual selection)是指在有性生殖生物中,同一性别个体之间为获得与异性的交配权而表现出有竞争力特征的选择(Darwin, 1871; Panhuis, 2001)。性选择是自然选择的一种重要形式,对于提高后代繁殖成功率及后代遗传多样性具有决定性作用。动物的性选择既包括交配前对配偶形态特征的选择,也包括交配后精卵之间的选择,如雄性间的精子竞争以及雌性对精子的隐性选择(Birkhead, 2002; Evans, 2003)。性选择机制研究不但有助于了解物种交配模式演化及行为进化趋势,而且也可为经济动物的人工繁育及种质资源保护提供重要的科学依据。
头足类是一类较为高等的海洋经济软体动物,我国近海头足类资源丰富,其在海洋渔业中占有重要地位(齐钟彦, 1998)。头足类在交配期具有复杂的繁殖行为,对其交配模式、繁殖行为和繁殖策略的研究对开展头足类人工养殖和种质资源修复具有重要的指导意义。头足类交配前性选择行为包括争斗与展示、雌性配偶选择、精子移除与替代等(郑小东等, 2009)。在雄性枪乌贼() (Hanlon, 2002; Sifner, 2004)和白斑乌贼()(Corner, 1980)中均存在争斗行为,蛸类的争斗行为比较少见,但有研究发现,2个雄蛸为争夺交配权而相互打斗(Hanlon, 1996)。雌性头足类对配偶具有选择性,在刺断腕蛸()和日本无针乌贼()中均发现此现象(Wada, 2006; Huffard, 2008),这种行为增强了精子竞争的强度,有利于优质基因的选择和种群的稳定。此外,澳大利亚巨乌贼()、金乌贼()和真蛸()中存在精子移除和替代现象(Naud, 2006; Wada, 2005; Wodinsky, 2008)。交配后性选择是雌性混交动物(一雌多雄或多雌多雄)中的一种普遍现象,其选择机制较为复杂。有关交配后性选择机制的假说主要有精子竞争、优中选优、避险策略和遗传不相容(Jennions, 2000; Beebee, 2008)。头足类一雌多雄或多雌多雄的交配模式非常普遍,在长鳍枪乌贼()、枪乌贼、水蛸()、真蛸等物种中都报道过(Buresch, 2001; Shaw, 2004; Larson, 2015; Quinteiro, 2011)。Naud等(2006)对澳大利亚巨乌贼的精子寿命研究中发现,雌性对其体内储存的精子具有选择性,优先利用纳精囊处的精子。此外,在金乌贼、真蛸和南部短尾章鱼()等多种头足类中都发现了精子竞争现象的存在(Wada, 2005; Wodinsky, 2008; Squires, 2014)。总之,头足类的性选择研究虽取得了一些成果,但仅限于部分种类,而且大部分研究只是实验室交配行为观察,缺乏系统的科学数据支持与机制研究。
多态性标记在亲权鉴定方面的应用,为性选择研究提供了新的工具。利用分子标记进行亲权检验,可以检验动物的交配模式,并协助验证交配后性选择机制假说(于红等, 2016)。微卫星标记因具有共显性、多态性高、易检测、重复性好等优点,在头足类父权检验中得到应用(Iwata, 2005; van Camp, 2004; 汪金海等, 2017)。
短蛸()隶属于软体动物门、头足纲、八腕目、蛸科、蛸属,是一种浅海底栖物种,在我国沿海地区均有分布,尤以黄、渤海产量最大,是我国北部沿海最重要的经济种之一。短蛸生长迅速,生命周期短,为雌雄异体,体内受精,一般在春季产卵,亲蛸有护卵行为。对于短蛸的交配模式已有相关研究(刘文芬等, 2018),但有关短蛸的性选择机制还未见报道。
本研究利用微卫星标记对短蛸子代胚胎进行父权鉴定与分析,研究亲本有无子代与父本形态特征、父权比例与父本形态特征及父母本遗传相似性的相关性,探讨短蛸交配前后的性选择机制,以期为短蛸种质资源保护及人工繁育提供科学依据,为海洋头足类动物性选择机制研究提供基础资料。
1 材料与方法
1.1 实验材料
本研究所用亲体与子代胚胎样本采集于山东东营养殖基地。将17头野生雄蛸和19头雌蛸于池中暂养,2天换1次水,温度控制在16℃~20℃,待其交配产卵后雄蛸陆续死亡,取腕部组织于无水乙醇中,-20℃保存备用(编号♂1~♂17)。雌蛸分批产卵,卵呈“葡萄串”状附着于池内事先放置的瓦罐中。为保证雌蛸与子代对应,将产卵的雌蛸与瓦罐放于已扎孔的塑料罐中。待受精卵孵化至出现眼点时,随机取3头雌蛸的腕部组织(编号♀1~♀3)及其对应所有子代胚胎(♀1~♀3子代数分别为57、79、40)保存备用。
1.2 实验方法
1.2.1 短蛸形态参数测量及相关分析 将短蛸充分伸展开,用数显电子游标卡尺测量36头亲蛸的胴长、胴宽和腕长(8个),用天平测量亲蛸总重。
1.2.2 短蛸基因组DNA的提取和检测 用CTAB法抽提亲蛸及176粒子代胚胎的基因组DNA,于30 μl 1×TE buffer (pH=8.0)溶解,用1%琼脂糖凝胶电泳检测基因组DNA的完整性,Thermo Scientific NanoDrop™ OneC超微量紫外分光光度计测定DNA浓度和纯度,将DNA浓度调至50 ng/μl,-20℃保存备用。
1.2.3 微卫星PCR特异性扩增 从实验室开发的微卫星标记中,挑选特异性好的14对引物DS152、DS220、DS132、DS280、DS310、DS135、DS210、DS226、DS25、DS150、DS106、DS137、DS290和DS116 (Feng, 2017; Yu, 2018),对17头雄蛸、3头雌蛸及对应的176粒子代胚胎的DNA样品进行PCR扩增。PCR反应体系为10 µl:模板DNA 1.0 µl (50 ng),10× PCR buffer (Mg2+plus) 1.0 µl,2.5 mmol/L dNTPs 1.0 µl,10 µmol/L正反引物对共2.0 µl,5 U/µlDNA聚合酶0.05 µl,ddH2O 4.95 µl。PCR反应程序:94℃预变性5 min;94℃变性45 s,退火温度45 s,72℃延伸45 s,循环35次;最后72℃延伸5 min。PCR产物用8%非变性聚丙烯酰胺凝胶电泳检测,硝酸银法染色,10 bp DNA ladder作为Marker检测等位基因的大小。
1.2.4 数据统计分析 利用SPSS 18.0软件分别对雄蛸和雌蛸的各形态参数进行相关性分析,并对二者的11个形态参数分别进行检验。用Cervus 3.0进行亲子鉴定分析,母本已知,参数设置:候选父本19,模拟子代为10000,亲本检测率100%,分型误差率1%,位点检测率100%,置信区间95%。结合亲子鉴定结果,采用检验检测有无子代与父本的11个形态参数、父权比例与父本形态参数的关联性。利用POPGENE 32分别检测父权比例与父母本遗传相似性和遗传距离的相关性。
2 结果
2.1 各形态参数的相关性分析
17头雄蛸形态参数的相关性分析结果显示,胴长、胴宽、8个腕与总重之间均存在相关性,其中,胴长、腕2、腕6与总重存在强相关性(相关系数>0.8)。胴宽、腕8与胴长,腕4、腕6与腕2也存在强相关性(表1)。
19头雌蛸形态参数的相关性分析结果显示,胴长、胴宽、8个腕与总重之间存在弱相关性(0.3<相关系数<0.8)。腕与腕之间,除腕2、腕4~腕8与腕1之间为弱相关外,其余均具有强相关性(表2)。
2.2 性别二态性
对雌蛸(19头)和雄蛸(17头)的11个形态参数进行检验分析,结果发现,雌蛸和雄蛸的总重和胴宽之间均差异显著(表3,<0.05),其他各参数之间差异不显著(>0.05),表明雌蛸和雄蛸的总重和胴宽均存在性别二态性。
2.3 父权分析
使用Cervus 3.0对3个家系子代及17个候选父本进行亲权鉴定,基于似然法从非排除亲本中选出最可能亲本,结果显示,176个子代鉴定率为100%,♀1家系57个子代来源于9个父本,子代比例分别为43.86% (♂14)、15.79% (♂17)、12.28% (♂3)、7.02% (♂2)、5.26% (♂6)、3.57% (♂4、♂5、♂13、♂16);♀2家系79个子代来源于10个父本,子代比例分别为24.05% (♂16)、21.52% (♂14)、17.72% (♂5)、13.92% (♂3)、10.13% (♂9、♂11)、6.33% (♂12)、3.80% (♂8)、2.53% (♂7、♂15);♀3家系40个子代来源于11个父本,子代比例分别为17.50% (♂2、♂14)、10.00% (♂12、♂16)、7.50% (♂3、♂9、♂13、♂17)、5.00% (♂5、♂10、♂15)。此外,鉴定出的10个父本♂2、♂3、♂5、♂9、♂12~♂17均与2个以上的母本发生了交配(表4)。
2.4 父本有无子代与形态参数的关联性分析
分别对3个家系有子代雄蛸和无子代雄蛸进行总重、胴长、胴宽和腕长(8个)共11个形态参数的检验,均未发现显著性差异(>0.05)(表5),表明短蛸雄蛸有无子代与其形态参数无相关性。
2.5 父权比例与父本形态参数的关联性分析
比较3个家系中子代比例高的父本(♂2、♂3、♂5、♂12、♂14、♂16和♂17)与其他父本(♂4、♂6、♂7、♂8、♂9、♂10、♂11、♂13和♂15)的11个形态参数 (表6),发现父权比例与父本形态大小无显著性差异(>0.05),表明父权比例与父本的形态参数无相关性。
表1 雄蛸各形态参数的相关系数
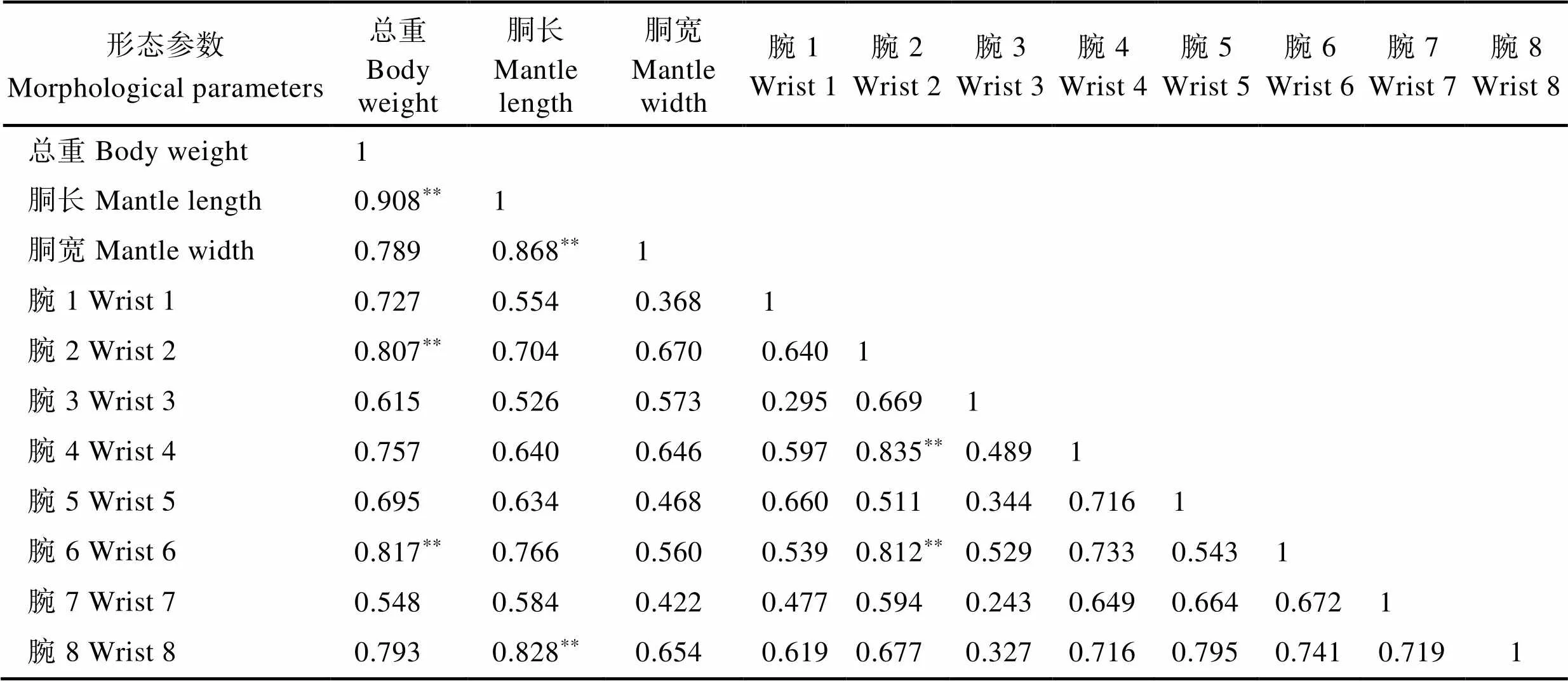
Tab.1 Correlation coefficient between morphological parameters in male octopus
**表示强相关性(相关系数>0.8)。下同 ** means significant correlation (coefficient>0.8). The same as below
表2 雌蛸各形态参数的相关系数
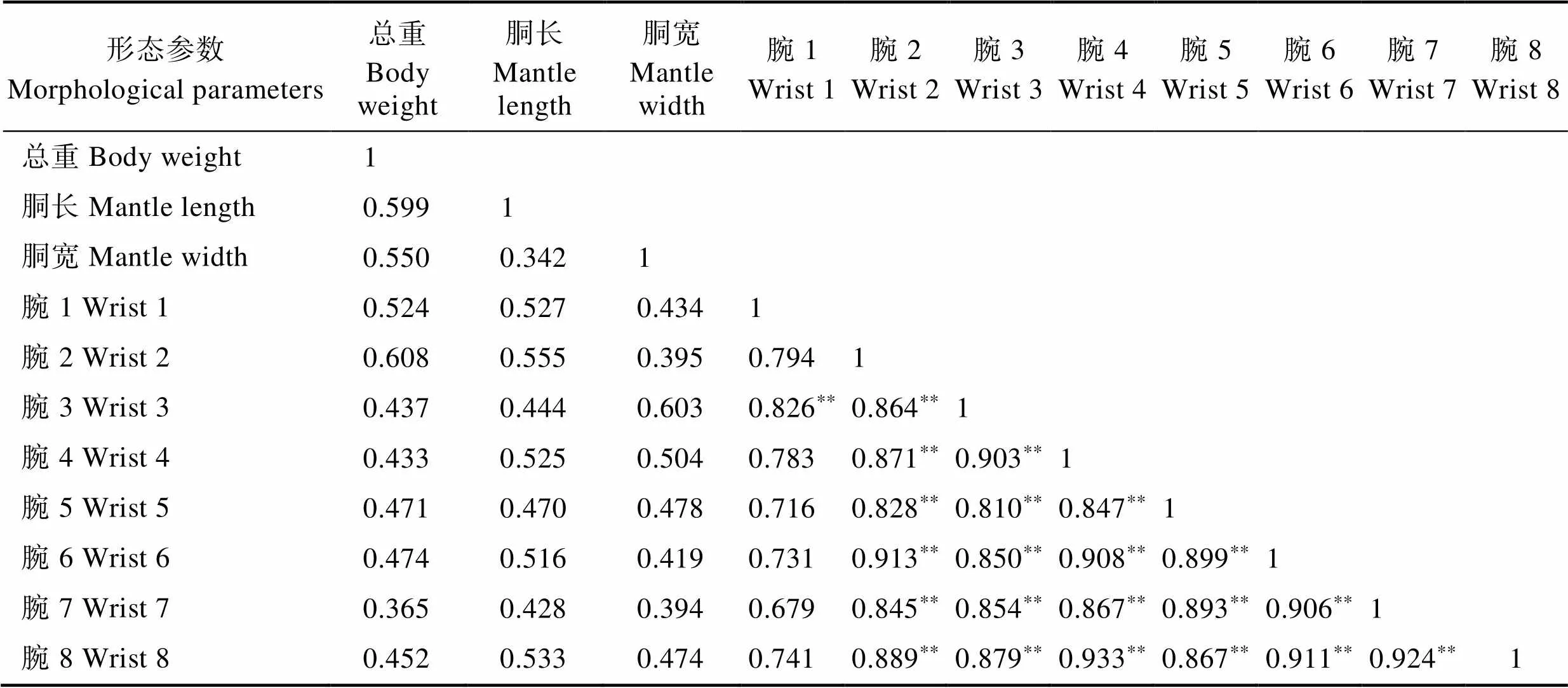
Tab.2 Correlation coefficient between morphological parameters in female octopus
表3 雄蛸与雌蛸各形态参数的检验分析
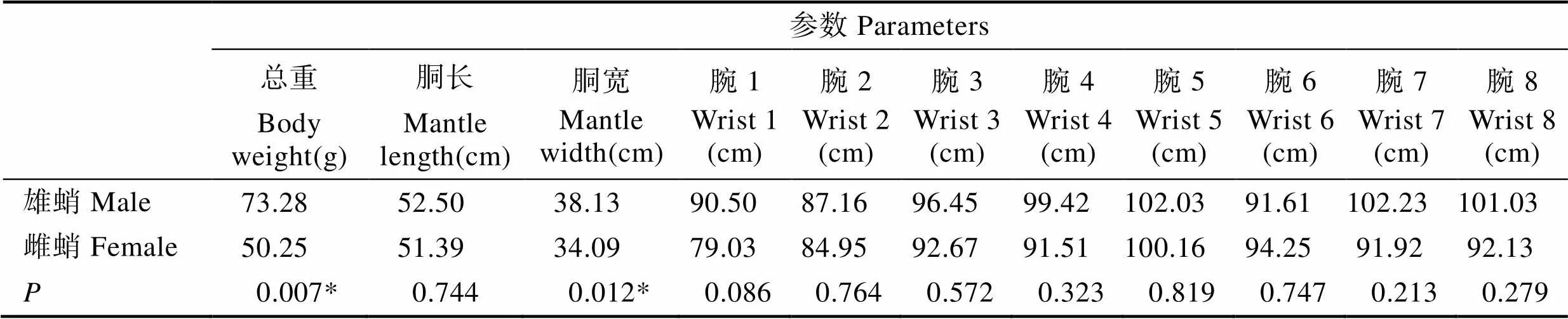
Tab.3 t-test analysis of morphological parameters of male and female octopus
*表示差异性显著(<0.05)。* means significant difference (<0.05)
表4 短蛸3个家系的亲子鉴定结果
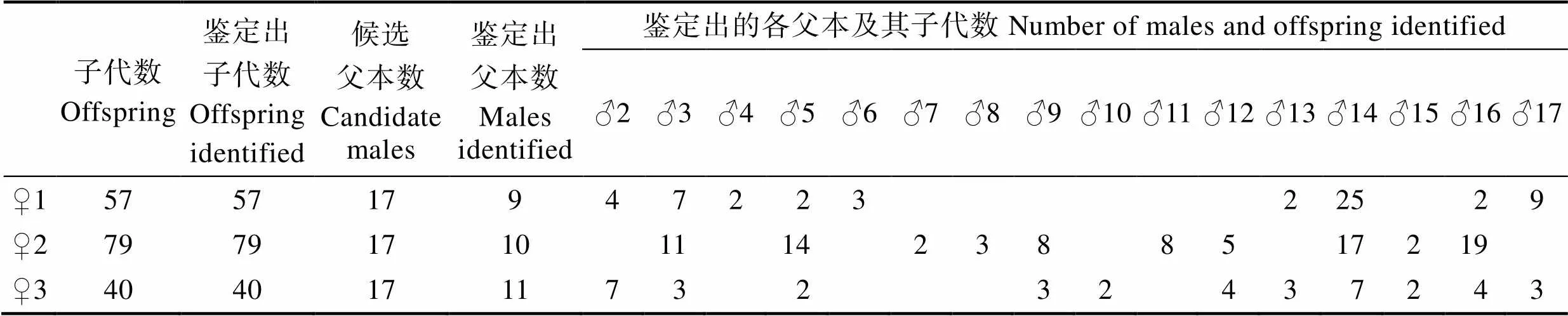
Tab.4 The paternity testing of three families in O. ocellatus
2.6 父权比例与父母本遗传相似性的关联性分析
利用Popgene 32分别计算3个家系子代比例高和子代比例低的父本与母本的遗传相似性与遗传距离(表7)。♀1家系中子代比例高的父本(♂3、♂14和♂17)与母本♀1的遗传相似性为0.4479,♀2家系中子代比例高的父本(♂3、♂5、♂14和♂16)与母本♀2的遗传相似性为0.5872,♀3家系中子代比例高的父本(♂2、♂12、♂14和♂16)与母本♀3的遗传相似性为0.4959,均大于子代比例低的父本与母本的遗传相似性。遗传距离结果与遗传相似性相反。父权比例与父母本遗传相似性及遗传距离有关,父母本遗传相似性越高,遗传距离越小,后代比例越高。
表5 3个家系有子代和无子代父本各形态参数的检验分析
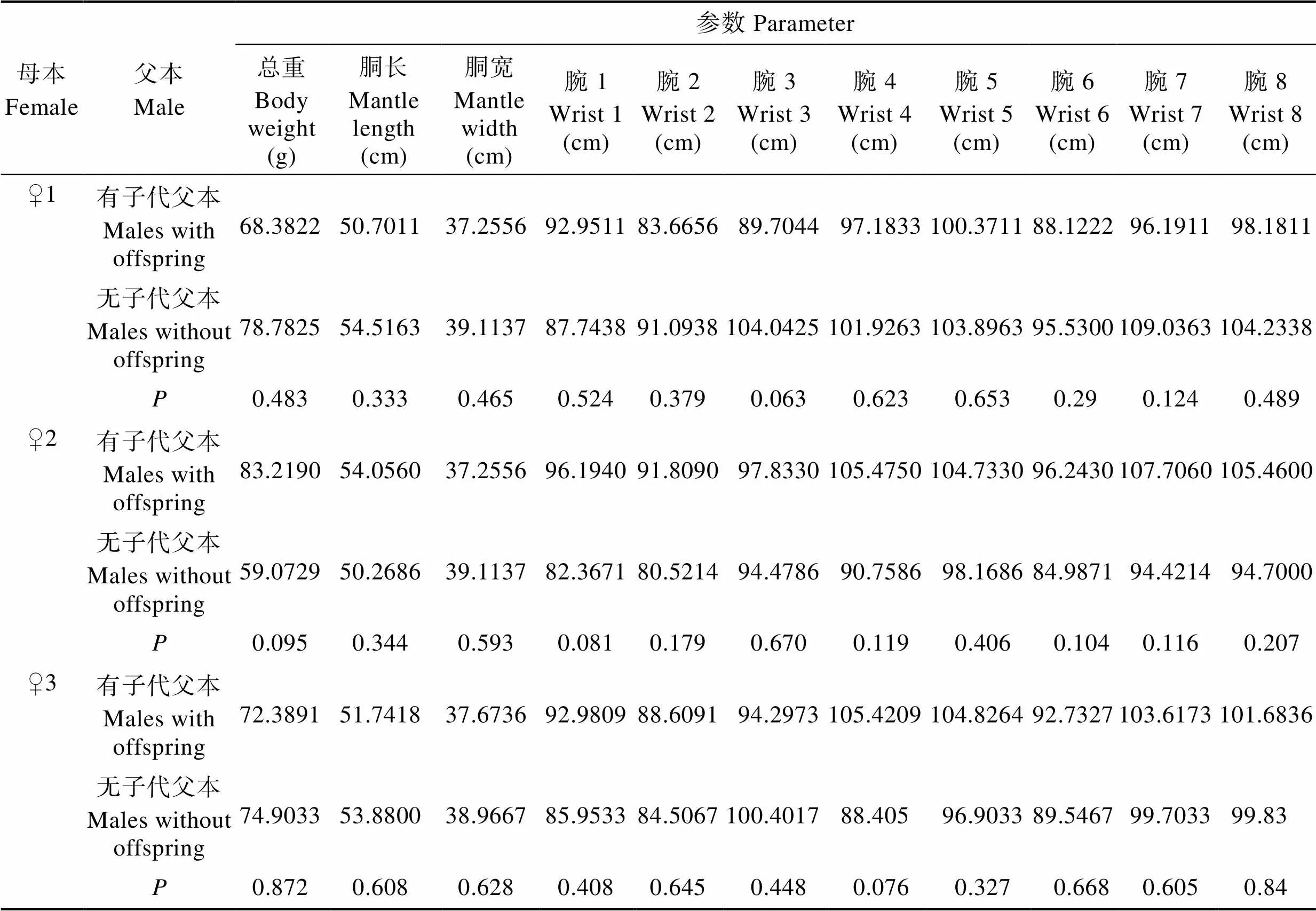
Tab.5 t-test analysis of morphological parameters between males with and without offspring
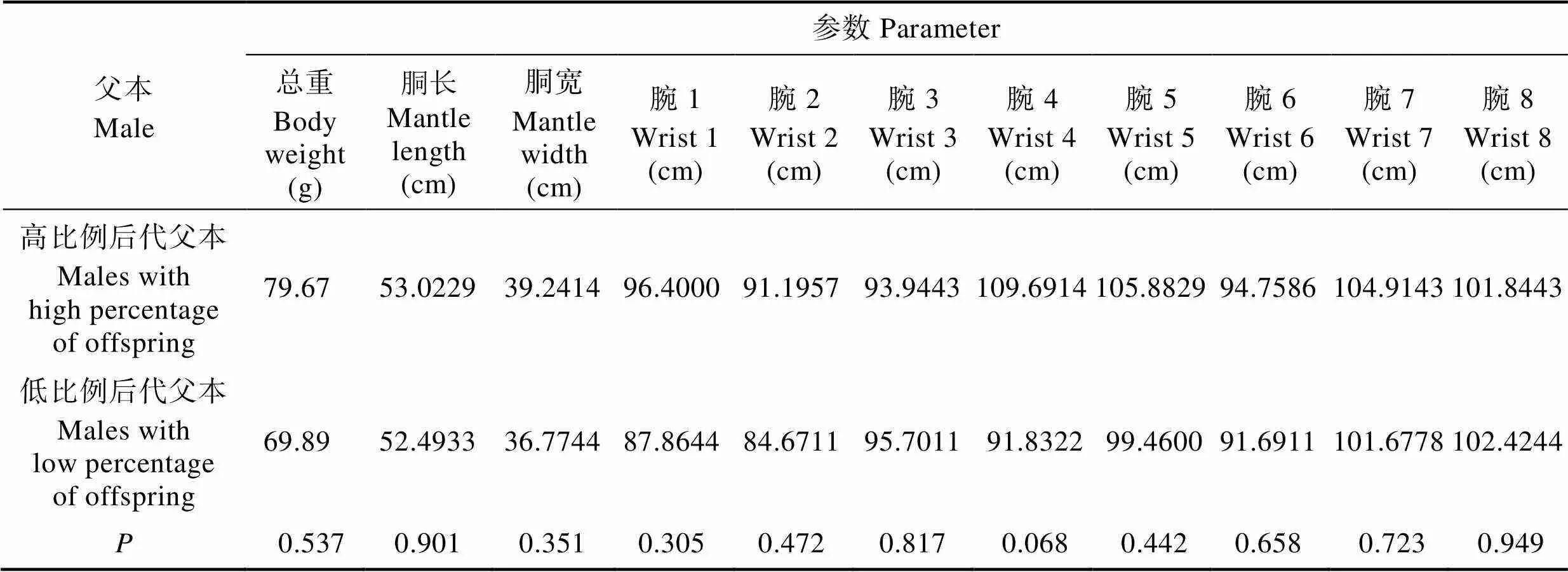
表6 高比例后代父本与低比例父本各形态参数的t检验分析
表7 高低比例父本与母本的Nei’s遗传相似性和遗传距离
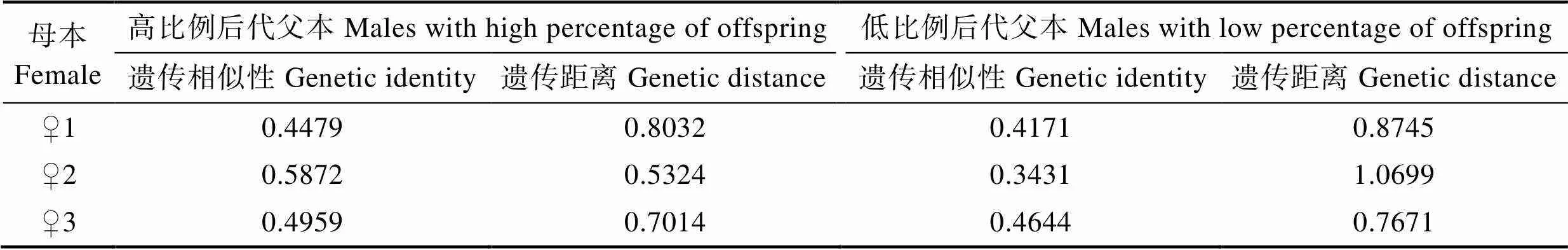
Tab.7 Nei’s genetic identity and distance between female parents and male parents with high and low offspring
3 讨论
本研究通过分析雄蛸的11个形态参数发现,胴长、腕2、腕6与总重存在显著相关性(相关系数>0.8),胴长与总重的相关系数最大,为0.908。形态参数之间存在显著相关性现象在贝类中有报道,刘小林等(2002)采用通径分析方法发现,栉孔扇贝()壳长、壳厚、壳高与活体重、净肉重、熟肉重的相关系数均达到极显著水平。孙秀俊等(2008)研究发现,虾夷扇贝的形态指标与鲜重具有显著相关性。利用多元分析研究物种的形态性状与体重等产量性状之间的关系,通过对形态性状的选择达到选中产量性状的目的。因此,在短蛸选育中,可以用胴长作为检测雄蛸个体大小的标准。本研究通过分析比较短蛸两性的11个形态参数发现,雌蛸和雄蛸的体重和胴宽存在性别二态性。性别二态性现象在其他蛸类中也有报道。雌性毯状章鱼()的体型可达2 m长,几乎比2.4 cm长的雄性大2个数量级,雌雄的体重比例可达10000 : 1~ 40000 : 1,是目前已知的雌雄形态特征差异最大的物种,具有显著的性别二态性(Norman, 2002)。
利用微卫星亲权鉴定技术对3个家系亲本和子代样品进行亲权鉴定,结果显示,3个家系的父本数分别为9、10和11,证实短蛸子代存在多父性现象,并且鉴定出10个父本(♂2、♂3、♂5、♂9、♂12~♂17)与2个以上的母本具有子代,说明短蛸的交配模式为多雌多雄,为真正意义的雌性混交动物,与刘文芬等(2018)的研究结果一致。雌性混交动物的交配模式为一雌多雄或多雌多雄,但观察到多重交配行为并不代表多重受精成功,只有后代中存在多父性现象,才算是真正遗传意义上的雌性混交动物。父本♂14在3个家系中的后代比例分别高达43.86% (25/57)、21.52% (17/79)、17.5% (7/40),略高于其他父本,说明父本♂14在精子竞争或雌性隐性选择中占优势。精子竞争模型分为最先精子优势、最后精子优势和无精子优势3种(朱道弘, 2003)。Wada等(2005)指出,由于金乌贼的精子取代行为使得最后一个与雌性金乌贼交配的雄性精子可能与更多的卵子结合。于红等(2016)研究发现,脉红螺()为最先雄性精子优先受精模式。本研究由于无法对短蛸进行标记,因而无法确认雄蛸与雌蛸的交配顺序,具体精子竞争模式有待进一步研究。
本研究发现,雄蛸有无子代与其形态参数无相关性。雄蛸没有子代可能是因为没有与雌蛸发生交配,或因为发生了交配,但精子被淘汰。由此可以推测,交配前及交配后雌性隐性选择中均不以雄性外部形态特征作为选择依据。父权比例与父本的形态大小参数无相关性,与脉红螺的研究结果一致(于红等, 2016);父权比例与父母本遗传相似性相关,父母本遗传相似性越高,后代比例越高。雌性和雄性同源性状间的遗传相关是普遍存在的,是多效基因作用的结果(Halliday, 1987)。短蛸两性在交配习性上存在遗传相关,推测符合Halliday等(1987)提出的遗传相关假说,认为精卵结合使得控制两性交配习性的一组共同的多基因座位发生交流,延缓自然选择驱动的性二型分化过程,进而促进雌性多次交配的进行,这可能有助于保持种群间的遗传相关平衡(Lande, 1980)。
本研究首次利用微卫星亲权鉴定技术探讨了短蛸交配前后的性选择机制,发现雌蛸在交配前未对雄蛸的形态特征进行选择,交配后的子代父权比例与父本形态参数无关,而与父母本遗传相似性有关。研究结果为揭示短蛸的性选择机制提供了重要线索,也为海洋头足类性选择机制及物种进化等研究提供了基础资料。
Beebee TJC, Rowe G. An introduction to molecular ecology. New York: Oxford University Press, 2008
Birkhead TR, Pizzari T. Postcopulatory sexual selection. Nature Reviews Genetics, 2002, 3(4): 262273
Buresch KM, Hanlon RT, Maxwell MR,. Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid. Marine Ecology Progress Series, 2001, 210: 161165
Corner BD, Moore HT. Field observations on the reproductive behavior of. Micronesia, 1980, 16: 235260
Darwin C. The Descent of man, and selection in relation to sex. London: John Murray, 1871
Evans JP, Zane L, Francescato S,. Directional postcopulatory sexual selection revealed by artificial insemination. Nature, 2003, 421(6921): 360363
Feng Y, Liu W, Xu X,. Construction of a normalized full-length cDNA library of cephalopod, and development of microsatellite markers. Journal of Ocean University of China, 2017, 16(5): 897904
Halliday T, Arnold S J. Multiple mating by females: A perspective from quantitative genetics. Animal Behaviour, 1987, 35(3): 939941
Hanlon RT, Messenger JB. Cephalopod behaviour. Cambridge: Cambridge University Press, 1996: 9197, 101110, 125127, 135
Hanlon RT, Smale MJ, Sauer WHH. The mating system of the squid(Cephalopoda, Mollusca) off South Africa: Fighting, guarding, sneaking, mating and egg laying behavior. Bulletin of Marine Science, 2002, 71(1): 331345
Huffard CL, Caldwell RL, Boneka F. Mating behavior of(d’Orbigny 1834) (Cephalopoda: Octopodidae) in the wild. Marine Biology, 2008, 154: 353362
Iwata Y, Munehara H, Sakurai Y. Dependence of paternity rates on alternative reproductive behaviors in the squid. Marine Ecology Progress Series, 2005, 298: 219228
Jennions MD, Petrie M. Why do females mate multiply? A review of the genetic benefits. Biological Reviews of the Cambridge Philosophical Society, 2000, 75(1): 2164
Lande R. Sexual dimorphism, sexual selection and adaptation in polygenic characters. Evolution, 1980, 34: 292305
Larson S, Ramsay C, Cosgrove JA. Multiple paternity and preliminary population genetics of giant pacific octopuses,, in Oregon, Washington and the southeast coast of Vancouver Island, BC. Diversity, 2015, 7: 195205
Liu WF, Feng YW, Wang WJ,. Development of microsatellite multiplex PCR sets and its application in the reproduction model analysis of. Progress in Fishery Sciences, 2018, 39(3): 152157 [刘文芬, 冯艳微, 王卫军, 等. 短蛸微卫星多重PCR体系建立及交配模式分析. 渔业科学进展, 2018, 39(3): 152157]
Liu XL, Chang YQ, Xiang JH,. Analysis of effects of shell size characters on live weight in Chinese scallop. Oceanologia et Limnologia Sinica, 2002, 33(6): 673678 [刘小林, 常亚青, 相建海, 等. 栉孔扇贝壳尺寸形状对活体重的影响效果分析. 海洋与湖沼, 2002, 33(6): 673678]
Naud MJ, Havenhand JN. Sperm motility and longevity in the giant cuttlefish,(Mollusca: Cephalopoda). Marine Biology, 2006, 148: 559566
Norman MD, Paul D, Finn J,. First encounter with a live male blanket octopus: The world’s most sexually size- dimorphic large animal. New Zealand Journal of Marine and Freshwater Research, 2002, 36(4): 733736
Panhuis TM, Butlin R, Zuk M,. Sexual selection and speciation. Trends in Ecology & Evolution, 2001, 16(7): 364371
Qi ZY. Economic Mollusca of China. Beijing: China Agriculture Press, 1998 [齐钟彦. 中国经济软体动物. 北京: 中国农业出版社, 1998]
Quinteiro J, Baibai T, Oukhattar L,. Multiple paternity in the common octopus(Cuvier, 1797), as revealed by microsatellite DNA analysis. Molluscan Research,2011, 31(1): 1520
Shaw PW, Sauer WHH. Multiple paternity and complex fertilization dynamics in the squid. Marine Ecology Progress Series, 2004, 270: 173179
Sifner SK, Vrgoc N. Population structure, maturation and reproduction of the European squid,, in the Central Adriatic Sea. Fisheries Research (Amsterdam), 2004, 69: 239249
Squires ZE, Wong BBM, Norman MD,. Multiple paternity but no evidence of biased sperm use in female dumpling squid. Marine Ecology Progress Series, 2014, 511: 93103
Sun XJ, Yang AG, Liu ZH,. Comparative analysis of morphological indices of Japanese scallops with 2 shell colors. Journal of Anhui Agricultural Sciences, 2008, 36(23): 1000810010 [孙秀俊, 杨爱国, 刘志鸿, 等. 2种壳色虾夷扇贝的形态学指标比较分析. 安徽农业科学, 2008, 36(23): 1000810010]
van Camp LM, Donnellan SC, Dyer AR,. Multiple paternity in field- and captive-laid egg strands of(Cephalopoda: Loliginidae). Marine and Freshwater Research, 2004(55): 819823
Wada T, Takegaki T, Mori T,. Sperm displacement behavior of the cuttlefish(Cephalopoda: Sepiidae). Journal of Ethology, 2005, 23: 8592
Wada T, Takegaki T, Tohru M,. Reproductive behavior of the Japanese spineless cuttlefish. Venus, 2006, 65(3): 221228
Wang JH, Han S, Zheng XD. Molecular identification of reproduction model for(Cephalopoda: Sepiida). Oceanologia et Limnologia Sinica, 2017, 48(1): 184189 [汪金海, 韩松, 郑小东. 金乌贼()繁殖模式的分子学鉴定. 海洋与湖沼, 2017, 48(1): 184189]
Wodinsky J. Reversal and transfer of spermatophores byand. Marine Biology, 2008, 155: 91103
Yu AQ, Fu CX, He CB,. Microsatellite records for volume 10, issue 3. Conservation Genetics Resources, 2018, 10: 127140
Yu H, Sun XQ, Yu RH,. Mechanism of postcopulatory sexual selection in. Journal of Fisheries of China, 2016, 40(6): 911916 [于红, 孙晓琪, 于瑞海, 等. 脉红螺交配后的性选择机制. 水产学报, 2016, 40(6): 911916]
Zheng XD, Han S, Lin XZ,. Research progress in Cephalopod reproductive behaviors. Journal of Fishery Sciences of China, 2009, 16(3): 459465 [郑小东, 韩松, 林祥志, 等. 头足类繁殖行为学研究现状与展望. 中国水产科学, 2009, 16(3): 459465]
Zhu DH. Sperm competition and adaptive significance of prolonged post-copulatory mounting in(Orthoptera: Catantopidae). Acta Ecologica Sinica, 2004, 24(1): 8488 [朱道弘. 小翅稻蝗的精子竞争及交配行为的适应意义. 生态学报, 2003, 24(1): 8488]
Sexual Selection inBased on Microsatellite Paternity Identification Technology
FAN Tingting1,2, FENG Yanwei2①, LIU Wenfen3, WANG Weijun4, YANG Jianmin4①
(1. National Demonstration Center for Experimental Fisheries Science Education, Shanghai Ocean University, Shanghai 201306; 2. Shandong Provincial Key Laboratory of Restoration for Marine Ecology, Shandong Marine Resource and Environment Research Institute, Yantai 264006; 3. Laboratory of Quality and Safety Risk Assessment for Aquatic Products, Ministry of Agriculture and Rural Affairs, Yangtze River Fisheries Research Institute, Chinese Academy of Fishery Sciences, Wuhan 430070; 4. School of Agriculture, Ludong University, Yantai 264025)
affiliated to Octopodidae under the phylum Mollusca, is one of the most important economic species along the northern coastal areas of China.It has been considered as a candidate for aquaculture enhancement and release owing to its short life span and rapid growth. However, little is known about sexual selection before and after copulation in this species. To explore the mechanism of sexual selection in, 11 morphological parameters of 36 adult octopuses were first measured and analyzed. Afterwards, 14 polymorphic microsatellite DNA markers were used to identify the paternity of 176 offspring and 3 female and 17 male parent candidates. The morphological parameters of male parents with and without offspring and those with high and low proportion of offspring were analyzed in three families by the-test. The genetic similarity between female and male parents which had high and low percentage of offspring was also calculated. The results showed that there was a strong correlation between mantle length, wrist 2, wrist 6, and body weight in male octopuses, and that sexual dimorphism in the total weight and mantle width between male and female octopuses existed. Paternity testing revealed all three females mated with more than two males and 10 males mated with at least two females, which confirmed the polygyny and polyandry pattern in this species. There was no correlation between male octopuses with or without offspring and their morphological parameters. Moreover, the proportion of offspring was not related to the morphological parameters of males but was correlated to the genetic similarity and genetic distance between female and male parents—higher the genetic similarity, higher the proportion of offspring. This study provides a scientific basis for the resource protection and artificial breeding of, and also, important information for research on the mechanism of sexual selection in marine cephalopods.
; Microsatellite markers; Paternity testing; Mating pattern; Sexual selection
S966
A
2095-9869(2020)02-0168-08
冯艳微,E-mail: fywzxm1228@163.com; 杨建敏,教授,E-mail: ladderup@126.com
2019-01-18,
2019-02-21
* 现代农业产业技术体系专项资金(CARS-49)、山东省优秀中青年科学家科研奖励基金项目(BS2014NY010)和山东省重点研发计划项目(2015GNC110017)共同资助 [This work was supporteded by China Agriculture Research System (CARS-49), Research Award Fund for Outstanding Young Scientist of Shandon Province (BS2014NY010), and Shandong Provincial Primary Research and Development Projects (2015GNC110017)]. 范婷婷,E-mail: fantingtingwork@163.com
10.19663/j.issn2095-9869.20190118002
http://www.yykxjz.cn/
范婷婷, 冯艳微, 刘文芬, 王卫军, 杨建敏. 基于微卫星亲权鉴定技术的短蛸性选择研究. 渔业科学进展, 2020, 41(2): 168–175
Fan TT, Feng YW, Liu WF, Wang WJ, Yang JM. Sexual selection inbased on microsatellite paternity identification technology. Progress in Fishery Sciences, 2020, 41(2): 168–175
FENG Yanwei. E-mail: fywzxm1228@163.com; YANG Jianmin. E-mail: ladderup@126.com
(编辑 冯小花)
猜你喜欢
现代妇产科进展(2022年1期)2022-12-07
中国生育健康杂志(2022年4期)2022-11-25
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
水产科学(2020年2期)2020-03-20
森林工程(2018年1期)2018-05-14
农业与技术(2016年22期)2017-03-07
江苏农业科学(2015年5期)2015-10-20
癌变·畸变·突变(2015年3期)2015-02-27
科技资讯(2014年26期)2014-12-03
