
圆斑星鲽piwil2基因的克隆与表达分析*
2020-03-25 06:03杨珍珍边力张岩常青陈四清刘长琳葛建龙胡建成张盛农
渔业科学进展 2020年2期
杨珍珍边力张岩常青陈四清刘长琳葛建龙胡建成张盛农
圆斑星鲽基因的克隆与表达分析*
杨珍珍1,2边 力2,3张 岩2,3常 青2,3陈四清2,3①刘长琳2,3葛建龙2,3胡建成2,3张盛农2
(1. 上海海洋大学水产科学国家级实验教学示范中心 上海 201306;2. 中国水产科学研究院黄海水产研究所农业农村部海洋渔业可持续发展重点实验室 青岛 266071;3. 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室 青岛 266071)
本研究采用RACE末端扩增方法,得到全长为3872 bp的圆斑星鲽()基因序列,命名为,开放阅读框(ORF)长为3192 bp,编码1063个氨基酸,5¢-UTR和3¢-UTR的长度分别140 bp和540 bp。基于ExPASy、SMART、Signal4.1和NCBI的保守结构域(CDD)数据库在线分析对蛋白序列结构进行预测,推断编码的氨基酸分子量为118.6 kDa,理论等电点为9.02,无跨膜结构及信号肽,有3个结构域:ArgoL1结构域、PAZ结构域及PIWI结构域。利用实时荧光定量PCR技术对圆斑星鲽不同发育时期的胚胎、仔稚鱼以及雌雄成鱼的不同组织表达模式进行分析。结果显示,基因从胚胎发育早期至高囊胚期均大量表达,之后呈下降趋势,直至孵化阶段。由于胚胎从卵裂至囊胚时期的发育过程主要受细胞质成分引导,直至原肠早期,mRNA开始大量转录合成,实现由母源型向合子型的过渡,推断是母源性基因。孵化后仔稚鱼68 d时,基因表达量显著高于其他时期,表明的功能可能与圆斑星鲽性腺分化过程相关;基因在雌雄成鱼性腺中的表达量显著高于其他组织,且卵巢中的表达量显著高于精巢,推测基因在卵巢功能的维持中发挥重要作用。本研究结果为解析圆斑星鲽性别决定机制提供了新的靶标基因,为建立全雌化苗种繁育技术打下坚实的理论基础。
圆斑星鲽;;基因克隆;表达分析
(P-element-induced wimpy testis)基因属于Argonaute (AGO)蛋白家族,该蛋白家族包括AGO亚家族、PIWI亚家族和亚家族(Kawaji, 2008)。其中,PIWI亚家族主要与piRNA (Piwi- interacting RNAs)结合形成PIWI/piRNA复合体,在配子发生、DNA甲基化及维持基因组完整性等方面起重要作用(Houwing, 2007; Bak, 2011)。PIWI亚家族的研究起步较晚,Lin等(1997)首次在果蝇()卵巢的生殖干细胞中发现基因,并证实该基因在果蝇生殖干细胞修复方面扮演重要角色,之后在小鼠() (Kuramochi-Miyagawa, 2001)、秀丽隐杆线虫()(Cox, 1998)和斑马鱼()(Tan, 2002)等模式生物中也发现了同源基因。小鼠中同源基因有和(Kuramochi-Miyagawa, 2001; Carmell, 2007);秀丽隐杆线虫中发现和两个同源基因(Cox, 1998);斑马鱼同源基因有和(Tan, 2002; 李丹等, 2010)。基因在生殖系细胞发育中发挥着重要作用,如原始生殖细胞(PGC)特化、生殖系细胞分化和生殖干细胞(GSC)维持(Megosh, 2006)。基因突变体果蝇表现为生殖干细胞分裂失调,最终导致不育(Hartig, 2007);小鼠的基因分别在精子发生的不同时期表达,、和的基因突变都会导致雄性个体的不育(Kim, 2006);秀丽隐杆线虫中和活性的降低会导致其生殖细胞数目明显减少(Cox, 1998);斑马鱼中和两个基因在生殖腺特异性高表达,突变体的生殖细胞出现细胞调亡现象(Houwing, 2007),突变体的生殖细胞不能实现雌雄分化(Houwing, 2014)。研究表明,基因编码的氨基酸具有2个高度保守的结构域,分别为PAZ(Piwi Argonaute Zwille)和PIWI(Parker, 2006)。PAZ结构域位于氨基酸链中间部分,包括1个寡聚核苷酸折叠区,结合PiRNAs 3¢端的核苷酸突出端;PIWI结构域靠近氨基酸链的C末端,具有RNaseH活性(Simon, 2011),PAZ和PIWI结构域可能在piwi-piRNA通路中起结合piRNAs的作用。随后,在鲤鱼()(Zhou, 2012)、鲫鱼()(Tao, 2018)、黄鳝()(Yi, 2014)、尼罗罗非鱼()(Xiao, 2013)、牙鲆()(Wang, 2016)、半滑舌鳎()(张丽燕, 2014)和大菱鲆()(Wang, 2017)等硬骨鱼类中也陆续发现基因。大菱鲆基因的研究显示,可能在胚胎和雌雌鱼性腺发育中发挥重要作用(Wang, 2017);基因是半滑舌鳎雄性精子发生过程中的关键基因,在精子发生早期,特别是精原干细胞发育过程中起重要作用(张丽燕, 2014);基因不仅能决定青鳉()生殖干细胞数量,甚至能控制生殖干细胞的迁移(Li, 2012)。
圆斑星鲽()俗称花斑宝,是一种冷温型的大型底栖鱼类,分布于我国北部的黄渤海和东海,具有食性范围广、生长速度快、抗病能力强、营养价值高、口味鲜美等特点,是优良的养殖品种。圆斑星鲽在生长过程中雌性个体的生长明显快于雄性个体,同龄雌雄个体的大小更是相差悬殊(谢忠明等, 2004)。鱼类性别决定机制研究有助于推动单性繁育技术的深入开展,目前,圆斑星鲽性别决定和生殖调控的分子机制仍不清楚,相关研究较少。柳学周等(2013)在圆斑星鲽脑中克隆到3种基因,认为可能是圆斑星鲽生殖调控的关键类型;张乐乐(2018)分离到2个性别相关基因(和),基因在卵巢特异性高表达,而基因在精巢高表达,但在圆斑星鲽中尚未确定性别决定基因。在大菱鲆等鲽形目鱼类及其他模式生物中均发现,基因在生殖细胞系发育、胚胎发育及雌雄性腺发育等过程中起着重要的作用,这种功能上的保守性在圆斑星鲽中是否存在值得进一步研究。
本研究通过RACE末端扩增技术得到基因的全长,分析其序列特征及编码的蛋白结构,并通过实时荧光定量PCR研究基因在各个发育时期的胚胎、仔稚鱼及雌雄成鱼不同组织的表达差异,旨在为阐明基因在性别分化和性腺发育过程中的作用奠定基础。
1 材料与方法
1.1 实验材料
圆斑星鲽受精卵、仔稚鱼和成鱼均取自山东省烟台开发区天源水产有限公司。收集不同发育时期的胚胎和仔稚鱼若干,单细胞期(卵裂前胚盘隆起时期)(0 h)、2细胞期(4 h)、8细胞期(10 h)、16细胞期(12 h)、32细胞期(13 h)、桑椹期(18 h)、高囊胚期(20 h)、低囊胚期(26 h)、原肠早期(27 h)、原肠中期(29 h)、原肠晚期(77 h)、神经胚期(98 h)、晶体形成期(136 h)、心跳出现期(163 h)、孵化前期(188 h)、脱膜孵化期(197 h)的胚胎及孵化后5、10、20、30、45、58、68、78、92 d的仔稚鱼,其中,受精卵孵化水温保持在(8.0± 0.5)℃,DO≥6 mg/L,pH为8.0左右,微量充气。仔鱼孵出后,移入水泥池培育,随着其生长发育,活动能力不断增强,充气量也应逐渐增大。初孵仔鱼的培养温度为(8.0±0.5)℃,以后逐日升高0.5℃,直至(12.0±0.5)℃稳定培育。再分别采取雌雄成鱼各3尾[体长(24.0±0.5) cm,体重(330±20) g]的不同组织(脑、眼、鳃、心、肝、肠、脾、性腺、肾及肌肉)。以上样品均迅速放入装有RNA later试剂的离心管中,将离心管放入液氮中冷冻,然后转到−80℃冰箱保存,用于总RNA的提取。
1.2 总RNA的提取及cDNA的合成
本实验采用Trizol法分别提取圆斑星鲽各个发育时期的胚胎、仔鱼及雌雄鱼不同组织的总RNA,用紫外分光光度计(NanoDrop 2000 Thermo Scientific)和1.2%琼脂糖凝胶电泳检测所提RNA的浓度及质量。cDNA利用反转录试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser(TaKaRa)合成,RACE-cDNA利用SMARTTMRACE cDNA Amplification RACE (TaKaRa)试剂盒合成。
1.3 Vvpiwil2基因全长克隆
从本实验室构建的转录组数据库中查找比对获得基因部分序列,利用Primer 5.0设计引物-S1和-A1(表1),对该cDNA序列进行验证,以圆斑星鲽成鱼性腺组织的cDNA为模板进行扩增。PCR反应体系(20 µl)为:Premix™ (LA™ Version 2.0) 10 µl、-S1(10 µmol/L) 0.8 µl、-A1(10 µmol/L) 0.8 µl、cDNA (1 µg/µl) 1 µl及ddH2O 6.4 µl。PCR 反应条件:94℃ 5 min;94℃ 30 s,50℃ 30 s,72℃ 1 min,35个循环;72℃ 10 min。扩增产物经1.2%琼脂糖凝胶电泳检测,并在紫外线下切胶,之后,用SanPrep柱式DNA凝胶回收试剂盒、pMD18-T Vector试剂盒(TaKaRa)和DH5α进行纯化、连接与转化培养,挑取阳性单克隆,经菌落PCR鉴定后,筛选目的菌液送至华大基因进行测序,测序结果与原始基因cDNA序列进行比对,确定为基因的核心序列。之后根据基因的核心序列设计特异性引物3¢GSP-1、3¢GSP-2、5¢GSP-1及5¢GSP-2,利用巢式PCR反应获得目的片段,PCR反应体系(20 µl):Premix™(LA™ Version 2.0) 10 µl、3¢或5¢特异性引物0.8 µl、UPM 0.8 µl、RACE- cDNA 1 µl及ddH2O 6.4 µl。PCR反应条件:94℃ 5 min;94℃ 30 s,60℃ 30 s,72℃ 1 min,35个循环;72℃ 10 min。将目的片段进行切胶回收、纯化、连接、转化、挑取阳性单克隆,最终将经过菌落PCR鉴定的目的菌液送到华大基因公司测序。
表1 本研究所使用的引物
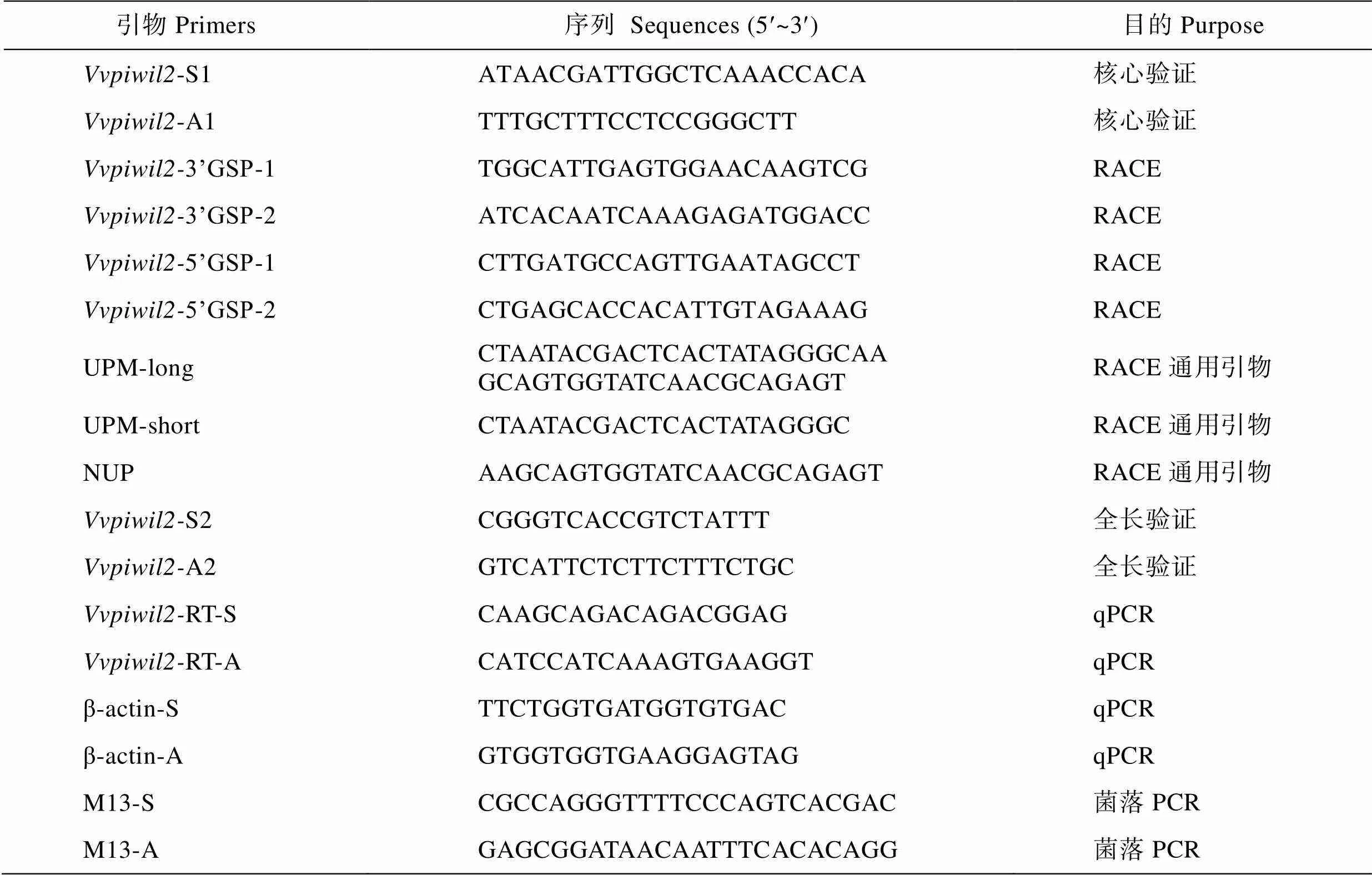
Tab.1 Nucleotide sequences of the PCR primers used in this study
1.4 Vvpiwil2基因序列分析
使用软件Contig Express对基因的测序结果进行拼接、验证,利用ORF Finder (http://www. ncbi.nlm.nih.gov/projects/gorf/orfig.cgi)预测基因的开放阅读框。利用ExPASy(https://web.expasy. org/compute_pi/)、SMART(http://smart.emblheidelberg. de/)、Signal4.1(http://www.cbs.dtu.dk/services/SignalP/)和NCBI(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)在线生物学软件分析编码蛋白的基本物理性质、信号肽、跨膜结构及结构域。用NCBI搜索同源蛋白序列,并使用DNAMAN软件对其进行蛋白序列的多重比较,利用MEGA5.2(Tamura, 2011)软件,采用邻接法(Neighbor-joining)构建系统进化树。
1.5 实时荧光定量PCR
根据基因的核心序列,利用Primer 5.0设计荧光定量特异性引物RT-A/S,β-actin-A/S作为内参,以不同发育时期的胚胎、仔稚鱼及雌雄成鱼的不同组织的cDNA为模板,使用Applied Biosystems™ 7500 Real Time PCR instrument定量仪,采用2–∆∆Ct的计算方法分析基因的相对表达量。20 μl反应体系: SYBR Premix Ex(2×) 10 µl、ROX Reference Dye Ⅱ (50x) 0.4 µl、RT-piwil2-S (10 μmol/L) 0.8 µl、RT-piwil2-A (10 μmol/L) 0.8 µl、cDNA(50ng/µl) 2 µl及ddH2O 6 µl,反应程序:95℃ 10 min;95℃ 30 s,95℃ 5 s,60℃ 34 s,40个循环;95℃ 15 s;60℃ 1 min;95℃ 15 s。
1.6 数据统计分析
数据处理为基因相对表达量的平均值±标准误(Mean±SE),生物学重复=3。使用SPSS 20.0软件对实时荧光定量PCR结果进行单因素方差分析(One-way ANOVA)和Duncan多重比较,<0.05表示差异显著,并用OriginPro 2017软件作图。
2 结果
2.1 Vvpiwil2基因序列特征及其编码的蛋白结构分析
克隆得到基因全长为3872 bp,将其命名为,其开放阅读框(ORF)长为3192 bp,编码1063个氨基酸,5¢-UTR和3¢-UTR分别长140 bp和540 bp。经过ExPASy、SMART、Signal4.1和NCBI在线生物学软件分析,推断的1063个氨基酸分子量为118.6 kDa,理论等电点为9.02,无跨膜结构及信号肽,有3个结构域,分别为428~473位46个氨基酸的ArgoL1结构域、477~616位140个氨基酸的PAZ结构域及602~1046位445个氨基酸的PIWI结构域(图1)。
2.2 Vvpiwil2基因同源性和系统发育分析

2.3 实时荧光定量PCR检测结果
通过实时荧光定量PCR研究基因在胚胎发育中的表达量。结果显示(图4),基因在整个胚胎发育阶段都有表达,从发育早期开始直到高囊胚期都大量表达,之后呈下降趋势,直至孵化阶段。研究基因在仔鱼不同发育时期中的表达量,结果显示(图5),基因在68 d时的表达量显著高于其他时期(<0.05)。
通过实时荧光定量PCR研究基因在雌雄成鱼各个组织的表达量,结果见图6。基因在雌雄成鱼的各个组织中均有表达。在雄鱼中,基因在精巢的表达量显著高于其他组织(<0.05);在雌鱼中,基因在卵巢的表达量也显著高于其他组织(<0.05)。雌雄成鱼之间进行比较可以发现,基因在卵巢中的表达量显著高于精巢(<0.05);此外,雄性成鱼的脑、眼的表达量高于雌性成鱼,雌性成鱼的心脏、肝的表达量高于雄性成鱼。

图1 Vvpiwil2基因cDNA序列全长及其编码的氨基酸序列
起始密码子ATG和终止密码子TGA均用黑色阴影标出,PolyA用下划线标出,ArgoL1结构域、PAZ结构域及PIWI结构域分别用双线边框、单线边框及灰色阴影标出
Start codon (ATG) and stop codon (TGA) are marked with dark shadow, and PolyA is marked with underline. The ArgoL1, PAZ and PIWI domains are separately marked with double lines border, single line border and gray shadow

图2 Vvpiwil2基因编码氨基酸与其他物种piwil2基因编码氨基酸的多序列比对
Black frame: PZA domain; Red frame: PIWI domain
黑色边框为PZA结构域,红色边框为PIWI结构域

The GenBank accession numbers ofgene are followed:(XP_019949653.1),(XP_018524808.1),(XP_023249908.1),(AQY15996.1),(XP_020445956.1),(XP_003445710.1),(NP_001281165.1)
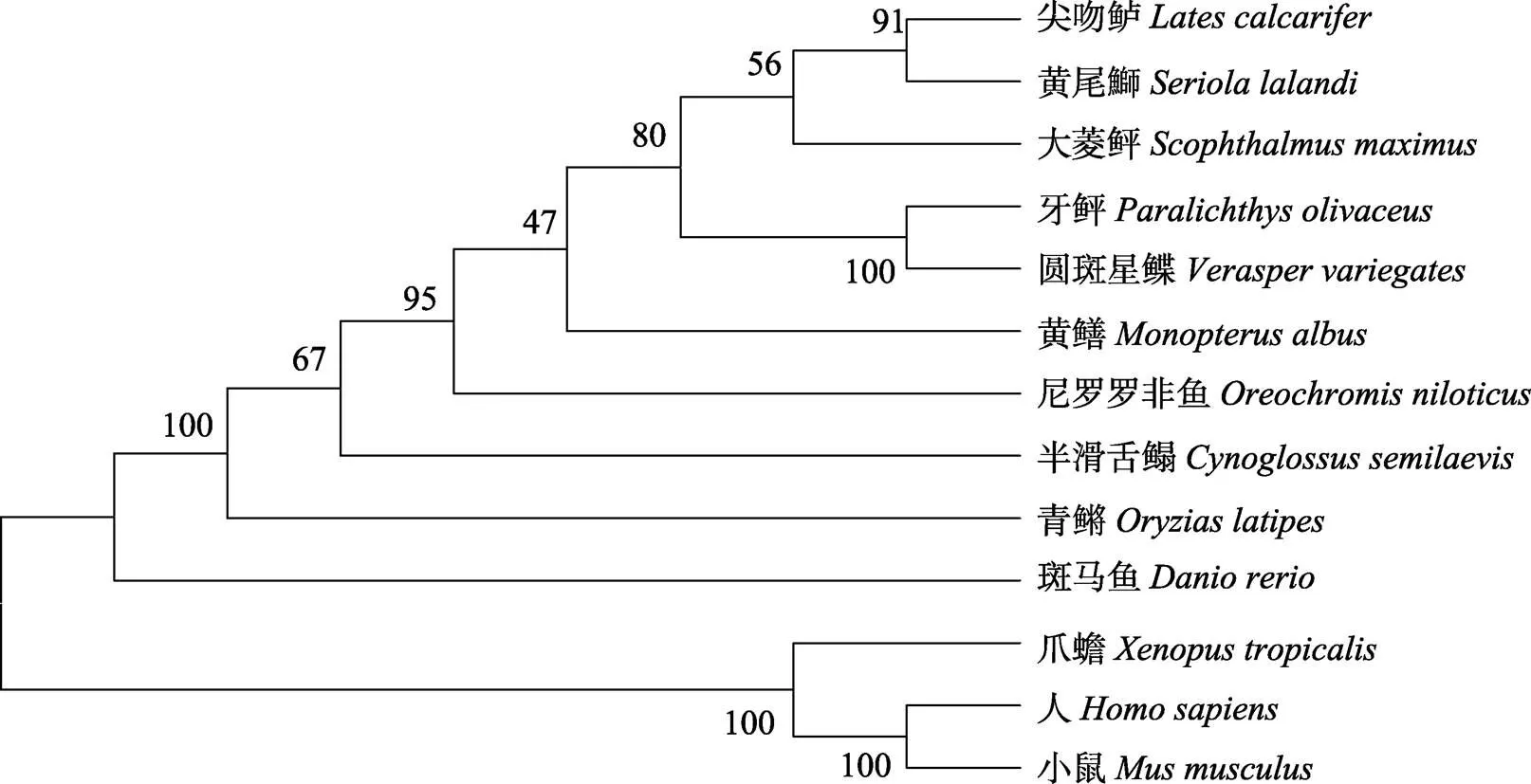
图3 piwil2氨基酸序列NJ进化树

The GenBank accession numbers ofgene are following:(XP_019949653.1),(XP_018524808.1),(XP_023249908.1),(AQY15996.1),(XP_020445956.1),(XP_003445710.1),(NP_001281165.1),(ACH96370.1),(NP_001153909.1),(NP_060538.2),(NP_067283.1),(NP_001106470.1)
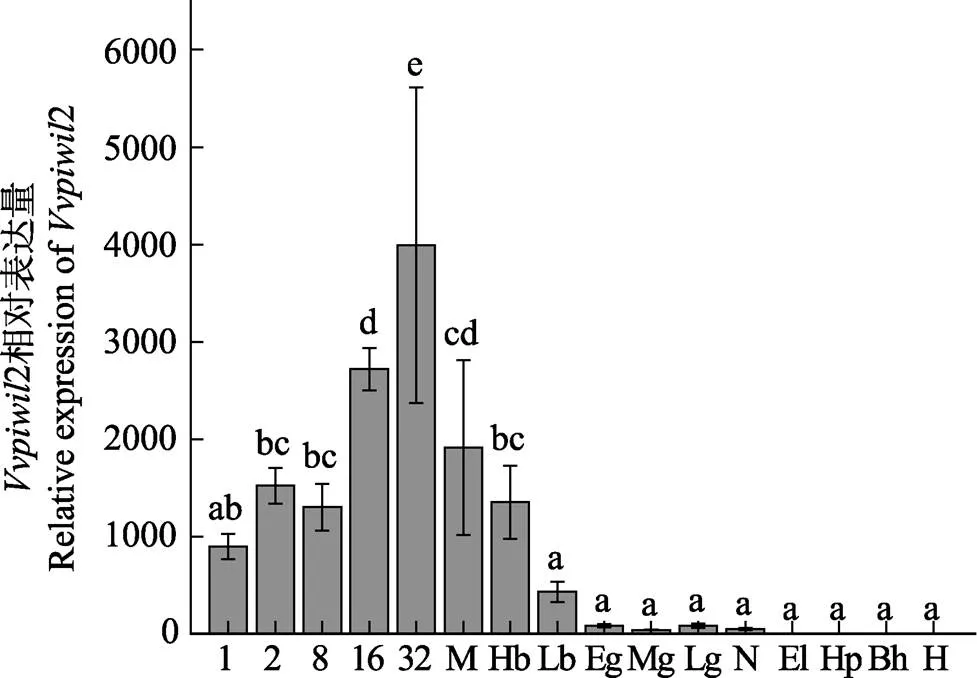
图4 Vvpiwil2基因在胚胎发育不同阶段中的表达
1: 单细胞期; 2: 2细胞期; 8: 8细胞期; 16: 16细胞期; 32: 32细胞期; M: 桑椹胚期; Hb: 高囊胚期; Lb: 低囊胚期; Eg: 原肠早期; Mg: 原肠中期; Lg: 原肠晚期; N: 神经胚期; El: 晶体形成期; Hp: 心跳出现期; Bh: 孵化前期; H: 脱膜孵化
不同字母间表示差异显著(<0.05),下同
1: 1-cell stage; 2: 2-cell stage; 8: 8-cell stage; 16: 16-cell stage; 32: 32-cell stage; M: Morula stage; Hb: High blastula; Lb: Low blastula; Eg: Early gastrula; Mg: Mid gastrula; Lg: Late gastrula; N: Neurula stage; El: Formation of eye len; Hp: Heart pulsation; Bh: Before the hatch; H: Hatching
Different letters indicate significant difference (<0.05), the same as below
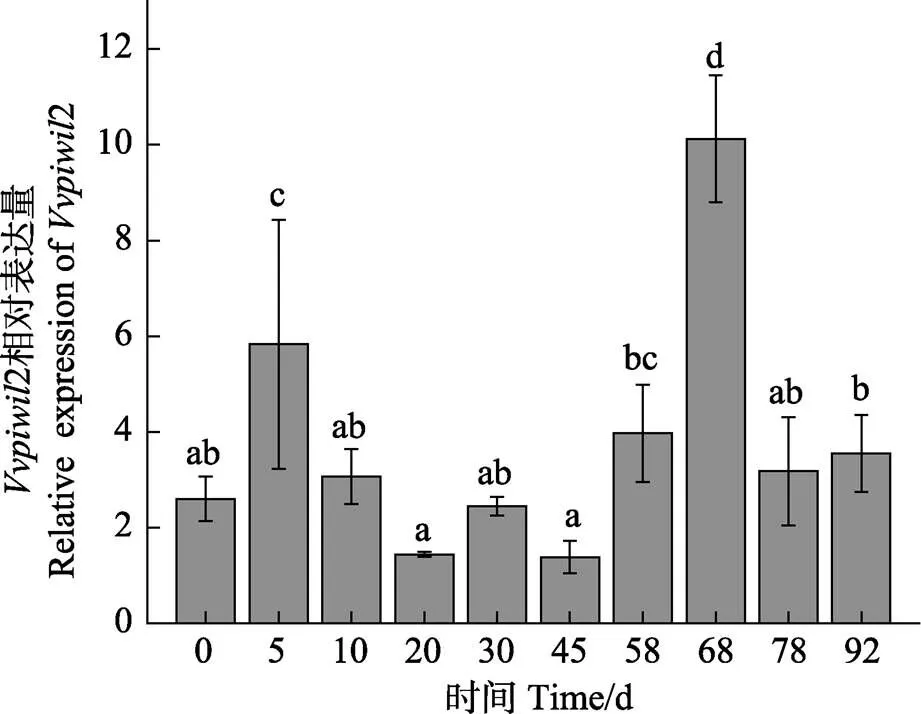
图5 Vvpiwil2基因在不同发育时期仔鱼中的表达
3 讨论
本研究从实验室构建的圆斑星鲽转录组数据库中查找比对获得基因部分序列,通过RACE末端扩增技术获得基因,将其命名为,基因全长为3872 bp,ORF长为3192 bp,编码一个由1063个氨基酸组成的多肽。同源性比对结果显示,包括圆斑星鲽在内的鱼类均含有保守的PAZ结构域和PIWI结构域,表明基因表达的蛋白具有结合小分子RNA的潜力。分子系统进化树分析显示,圆斑星鲽与硬骨鱼类单独聚成了一支,Houwing等(2014)的研究显示,斑马鱼基因在生殖细胞分化中起重要作用,Li等(2012)研究表明,在青鳉中基因不仅能决定生殖干细胞数量,甚至能控制生殖干细胞的迁移。在进化树中,圆斑星鲽与鲽形目中的牙鲆、大菱鲆及半滑舌鳎关系较近,大菱鲆基因可能在胚胎和雌雄性腺发育中发挥重要作用,而半滑舌鳎基因在雄性精子发生过程中起关键作用,特别是在精原干细胞发育的过程中起重要作用。基于系统进化分析,推测基因在圆斑星鲽可能具有相似的功能,即在生殖细胞系发育、胚胎发育及雌雄性腺发育等过程中发挥作用。
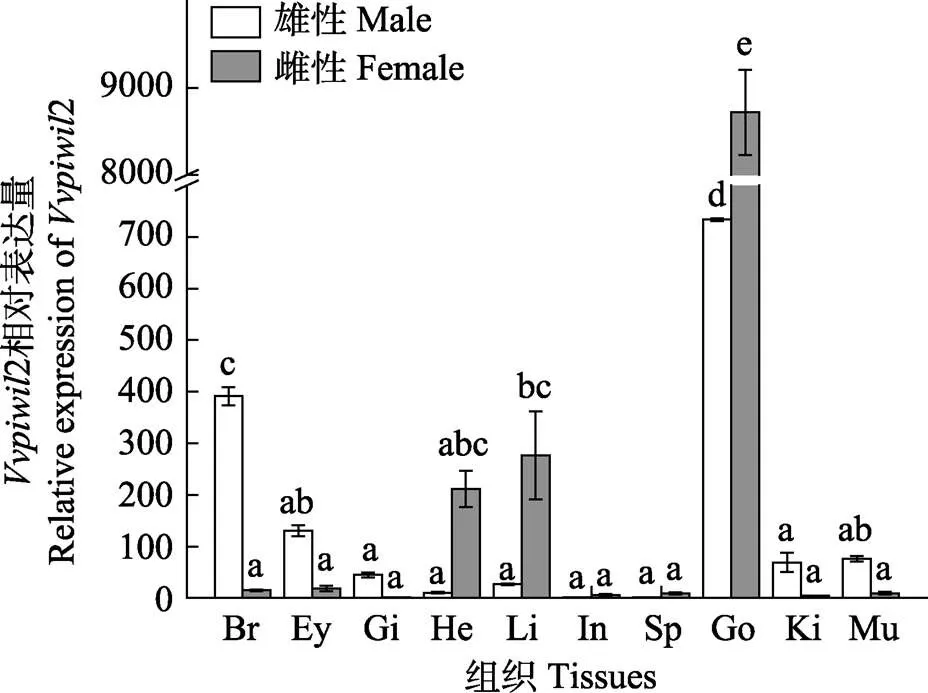
图6 Vvpiwil2基因在不同组织中的表达
Br: 脑; Ey: 眼; Gi: 鳃; He: 心脏; Li: 肝脏; In: 肠; Sp: 脾; Go: 性腺; Ki: 肾; Mu: 肌肉
Br: Brain; Ey: Eye; Gi : Gill; He: Heart; Li: Liver; In: Intestines; Sp: Spleen; Go: Gonad; Ki: Kidney; Mu: Muscle
利用实时荧光定量PCR技术对基因在胚胎发育不同阶段的表达模式进行了分析。结果显示,该基因在整个胚胎发育期间都有表达,从发育早期直到高囊胚期都呈高表达,之后开始呈下降趋势,直至孵化阶段,这一表达模式与斑马鱼(Tan, 2002)、青鳉(李名友, 2008)相似。从卵裂开始至囊胚时期的发育阶段主要受细胞质成分引导,一直到卵裂较晚的原肠早期,胚胎mRNA才大量转录合成,实现由母源型向合子型的过渡,因此,认为基因是母源性基因。研究基因在不同发育时期仔鱼中的表达量,结果显示,基因在仔鱼68 d时的表达量显著高于其他发育时期。张乐乐(2018)研究表明,圆斑星鲽仔鱼在60日龄左右性腺开始分化,幼鱼出现卵巢腔,与本研究中基因表达量显著上升的时间接近,表明基因可能在圆斑星鲽的性别分化过程中发挥功能。张丽燕(2014)研究表明,基因不是性别分化的相关基因,半滑舌鳎的基因与雄鱼精子发生相关,主要作用于精子发生的早期阶段,而非性别分化期,表明基因的功能存在一定的种间差异。
利用实时荧光定量PCR对雌雄鱼中基因的表达模式进行了分析。结果显示,基因在精巢与卵巢中的表达量均显著高于其他组织,与大菱鲆(Wang, 2017)、半滑舌鳎(张丽燕, 2014)、斑马鱼(Tan, 2003)及青鳉(Zhao, 2012)的同源基因表达相似,与人和鼠等哺乳动物仅在精巢中特异表达不同(Zeeman, 2002; Beyret, 2011)。基因在圆斑星鲽卵巢中的表达量显著高于精巢,说明基因可能在卵巢功能的维持中发挥重要作用。这一结果与大菱鲆中的表达相似,但在半滑舌鳎、斑马鱼等研究中,基因在精巢的表达量显著高于卵巢,再次表明基因的功能在不同种类间存在一定差异。本研究还发现,基因在脑、眼、鳃、心脏、肝、肠、脾、肾及肌肉组织中均有表达,并且雄鱼的脑、眼的表达量高于雌鱼,雌鱼的心脏、肝的表达量高于雄鱼。李名友(2008)研究发现,在青鳉中不仅在性腺组织中大量表达,在眼和脑组织中也有表达;Tao等(2016)研究发现,罗非鱼和除在性腺中大量表达外,在骨骼肌中也有表达;Wang等(2017)研究发现,在大菱鲆中还在肌肉、鳃和肾脏在内的体细胞组织中表达,认为Piwi蛋白不仅在生物生殖发育过程中是必需的,而且在体细胞组织中也发挥重要功能;张丽燕等(2014)研究发现,半滑舌鳎基因在脾脏、肝脏、心肌、肠、脑、皮肤、肾脏、肌肉、鳃及血液中也有微量表达。本研究也表明,基因不仅在性别分化等生殖发育过程中发挥作用,而且在其他的生命过程中也具有重要功能。关于基因在圆斑星鲽雄鱼的脑、眼的表达量高于雌鱼,雌鱼的心脏、肝的表达量高于雄鱼这一结论,尚未在其他相关物种中发现相似观点,仍需要进一步研究。
4 结论
本研究以圆斑星鲽性腺为模板,扩增得到了基因全长,并将其命名为。在线保守结构域分析显示,基因具有PIWI亚家族典型的PAZ结构域和PIWI结构域。利用实时荧光定量PCR技术对圆斑星鲽不同发育时期胚胎、仔稚鱼的表达模式进行了分析。结果显示,基因为母源性基因,可能与圆斑星鲽性腺分化过程相关。组织表达分析发现,基因在雌雄成鱼性腺中的表达量显著高于其他组织中,卵巢中的表达量显著高于精巢,推测基因可能在卵巢功能的维持中发挥重要作用。本研究结果为解析圆斑星鲽性别决定机制提供了新的靶标基因,为建立全雌化苗种繁育技术奠定了理论基础。
Bak CW, Yoon TK, Choi Y. Functions of PIWI proteins in spermatogenesis. Clinical and Experimental Reproductive Medicine, 2011, 38(2): 61–67
Beyret E, Lin H. Pinpointing the expression of piRNAs and function of the PIWI protein subfamily during spermatogenesis in the mouse. Developmental Biology, 2011, 355(2): 215– 226
Carmell MA, Girard A, van de Kant HJG,. MIWI2 is essential for spermatogenesis and repression of transposons in the mouse male germline. Developmental Cell, 2007, 12(4): 503–514
Cox DN, Chao A, Baker J,. A novel class of evolutionarily conserved genes defined byare essential for stem cell self-renewal. Genes and Development, 1998, 12(23): 3715– 3727
Hartig JV, Tomari Y, Frstemann K. piRNAs-the ancient hunters of genome invaders. Genes and Development, 2007, 21(14): 1707–1713
Houwing S, Kamminga LM, Berezikov E,. A role for Piwi and piRNAs in germ cell maintenance and transposon silencing in zebrafish. Cell, 2007, 129(1): 69–82
Houwing S, Berezikov E, Ketting RF. Zili is required for germ cell differentiation and meiosis in zebrafish. European Molecular Biology Organization, 2014, 27(20): 2702–2711
Kawaji H, Hayashizaki Y. Exploration of small RNAs. Public Library of Science Genetics, 2008, 4(1): 3–8
Kim VN. Small RNAs just got bigger: Piwi-interacting RNAs (piRNAs) in mammalian testes. Genes and Development, 2006, 20(15): 1993–1997
Kuramochi-Miyagawa S, Kimura T, Yomogida K,. Two mouse-related genes:and. Mechanisms of Development,2001, 108(1): 121–133
Li D, Sun HQ, Zhao J,. Initial study on zili functions to the development of the germ layers during zebrafish early embryogenesis. Journal of Sichuan University (Medical Science), 2010, 41(6): 923–926 [李丹, 孙华钦, 赵君, 等. zili在斑马鱼早期胚胎发育胚层分化中作用的初步研究. 四川大学学报(医学版), 2010, 41(6): 923–926]
Li M, Hong N, Gui J,. Medaka piwi is essential for primordial germ cell migration. Current Molecular Medicine, 2012, 12(8): 1040–1049
Li MY. Expression and functional analysis ofandin Medaka. Doctoral Dissertation of Institute of Hydrobiology, Chinese Academy of Sciences, 2008 [李名友. 青鳉和基因的表达模式和功能分析. 中国科学院水生生物研究所博士研究生学位论文, 2008]
Lin H, Spradling AC. A novel group ofmutations affects the asymmetric division of germline stem cells in theovary. Development, 1997, 124(12): 2463–2476
Liu XZ, Xu YJ, Liao MJ,. Cloning and expression characteristics of gonadotropin releasing hormone genes in spotted halibut. Journal of Fishery Sciences of China, 2013, 20(1): 12–24 [柳学周, 徐永江, 廖梅杰, 等. 圆斑星鲽促性腺激素释放激素基因克隆及表达特性. 中国水产科学, 2013, 20(1): 12–24]
Megosh HB, Cox DN, Campbell C,. The role of PIWI and the miRNA machinery in Drosophila germline determination. Current Biology, 2006, 16(19): 1884–1894
Parker JS, Barford D. Argonaute: A scaffold for the function of short regulatory RNAs. Trends in Biochemical Sciences, 2006, 31(11): 622–630
Simon B, Kirkpatrick JP, Eckhardt S,. Recognition of 2'-O-methylated 3'-end of piRNA by the PAZ domain of a Piwi protein. Structure, 2011, 19(2): 172–180
Tamura K, Peterson D, Peterson N,. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution, 2011, 28(10): 2731–2739
Tan CH, Lee TC, Weeraratne SD,., the zebrafish homologue of the Drosophila: Co-localization with vasa at the embryonic genital ridge and gonad-specific expression in the adults. Mechanisms of Development, 2002, 119(2): S221–S224
Tao M, Li S, Song C,. Elevated expression ofgene in the pituitaries of allotriploid crucian carp. Journal of the World Aquaculture Society, 2018, 49(2): 302–314
Tao W, Sun L, Chen J,. Genomic identification, rapid evolution, and expression of Argonaute genes in the tilapia,. Development Genes and Evolution, 2016, 226(5): 339–348
Wang CL, Wang ZP, Wang JQ,. Identification of candidate piRNAs in the gonads of(Japanese flounder). Zoological Research, 2016, 37(5): 301–306
Wang H, Wang B, Liu X,. Identification and expression ofin turbot, with implications of the involvement in embryonic and gonadal development. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2017, 208–209: 84–93
Xiao J, Zhou Y, Luo Y,. Suppression effect of LHRH-A and hCG on Piwi expression in testis of Nile tilapia. General and Comparative Endocrinology, 2013, 189(1): 43–50
Xie ZM, Chen SQ, Zhang Y,. Fish culture technology of flatfish sole. Beijing: Jindun Publishing House, 2004 [谢忠明, 陈四清,张岩, 等. 鲽鳎鱼类养殖技术. 北京: 金盾出版社, 2004]
Yi M, Chen F, Luo M,. Rapid evolution of piRNA pathway in the teleost fish: Implication for an adaptation to transposon diversity. Genome Biology and Evolution, 2014, 6(6): 1393–1407
Zeeman AM. Molecular characterization of, a human member of thegene family whose overexpression is correlated to seminomas. Oncogene, 2002, 21(25): 3988– 3999
Zhang LL. The study of gonadal differentiation process and sex-related gene,of spotted halibut. Master´s Thesis of Shanghai Ocean University, 2018 [张乐乐. 圆斑星鲽性腺分化过程及性别相关基因的研究. 上海海洋大学硕士研究生学位论文, 2018]
Zhang LY. Molecular cloning and expression analysis ofandgene in the half-smooth tongue-sole,. Master´s Thesis of Dalian Ocean University, 2014 [张丽燕. 半滑舌鳎和基因的克隆与表达分析. 大连海洋大学硕士研究生学位论文, 2014]
Zhao H, Duan J, Cheng N,. Specific expression ofandin medaka () germ cells. Biochemical and Biophysical Research Communications, 2012, 418(4): 592–597
Zhou Y, Wang F, Liu S,. Human chorionic gonadotropin suppresses expression of Piwis in common carp () ovaries. General and Comparative Endocrinology, 2012, 176(2): 126–131
Cloning and Expression of theGene in the Spotted Halibut ()
YANG Zhenzhen1,2, BIAN Li2,3, ZHANG Yan2,3, CHANG Qing2,3, CHEN Siqing2,3①, LIU Changlin2,3, GE Jianlong2,3, HU Jiancheng2,3, ZHANG Shengnong2
(1. National Demonstration Center for Experimental Fisheries Science Education, Shanghai Ocean University, Shanghai 201306; 2. Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071; 3. Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266071)
The spotted halibut () is a rare and valuable marine fish species that inhabits the coast of northern China. Mature females are much larger than males because of their faster growth rate. It will create substantial economic benefits to establish an all-female breeding technique for. A better understanding of sex-related genes will contribute to the improvement of a single-sex breeding technique. In this study, we successfully isolated thegene of, which was namedThe total length was 3872 bp, including a 3192 bp open reading frame (ORF), encoding 1063 amino acids; the 5′UTR was 140 bp and the 3′UTR was 540 bp. Based on ExPASy, SMART, Signal4.1, and the NCBI Conservative Domain Database (CDD) biological analysis, the ORF encoded a putative protein, with a predicted molecular weight of 118.6 kDa and an isoelectric point of 9.02. No transmembrane structure or signal peptide site was detected. There were three domains: the ArgoL1, PAZ, and PIWI domains. Real-time fluorescence quantitative PCR technique was used to analyze the expression patterns of thegene at different stages of embryo and larvae. The results showed that thegene was abundantly expressed from early development to the high blastocyst stage, and then declined until the hatching stage. The developmental stage of the embryo from cleavage to the blastocyst stage was mainly guided by the cytoplasmic component. The mRNAs began to be transcribed and synthesized in a large amount at the early gastrula stage, then the transition from maternal to zygote occurred. Therefore, the results indicated that thegene was a maternal gene. After hatching, the expression of thegene at 68 days post hatching was significantly higher than during other stages, which demonstrated that thegene was associated with gonadal differentiation. The expression level of thegene in gonads was significantly higher than in other tissues, and the expression level in the ovary was significantly higher than in the testis, revealing that thegene might play an important role in the maintenance of ovarian functions. The results of this study provide a potential sex determinant gene forand lay a solid theoretical foundation for the establishment of an all-female breeding technique.
;; Genecloning; Expression analysis
S917.4
A
2095-9869(2020)02-0103-10
陈四清,研究员,E-mail: chensq@ysfri.ac.cn
2019-01-30,
2019-03-05
* 中国水产科学研究院黄海水产研究所基本科研业务费(20603022016005)资助 [This work was supported by Special Scientific Research Funds for Non-Profit Institute, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences (20603022016005)]. 杨珍珍,E-mail: 1163603557@qq.com
10.19663/j.issn2095-9869.20190130001
http://www.yykxjz.cn/
杨珍珍, 边力, 张岩, 常青, 陈四清, 刘长琳, 葛建龙, 胡建成, 张盛农. 圆斑星鲽基因的克隆与表达分析. 渔业科学进展, 2020, 41(2): 103–112
Yang ZZ, Bian L, Zhang Y, Chang Q, Chen SQ, Liu CL, Ge JL, Hu JC, Zhang SN. Cloning and expression of thegene in the spotted halibut (). Progress in Fishery Sciences, 2020, 41(2): 103–112
CHEN Siqing, E-mail: chensq@ysfri.ac.cn
(编辑 冯小花)
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
文萃报·周五版(2022年24期)2022-06-21
四川蚕业(2020年2期)2020-12-18
实用肿瘤学杂志(2020年4期)2020-12-08
中外医学研究(2020年4期)2020-04-27
现代农业研究(2017年10期)2018-01-02
Coco薇(2015年12期)2015-12-10
医学综述(2011年12期)2011-12-09
亚热带农业研究(2011年3期)2011-09-29
大学(2008年4期)2008-07-10
