
乳腺癌中SLC5A8基因的甲基化及转录水平
2020-03-23 10:55:06刘雅涵谢天皓孟庆旭靳小石2
医学研究与教育 2020年1期
刘雅涵 谢天皓 孟庆旭 靳小石2
(1.河北大学临床医学院,河北 保定 071000;2.河北省普通外科数字医学基础研究重点实验室,河北 保定 071000;3.河北大学附属医院,河北 保定 071000)
根据2019年1月份国家癌症中心发布的最新全国癌症统计数据,目前恶性肿瘤发病率最高的为肺癌,其次为乳腺癌,而女性发病率最高的为乳腺癌,死亡率在女性占到第5位[1]。在这样的背景下,乳腺癌的关注和研究显得极为重要。基因的改变在乳腺癌中有着重要的作用。SLC5A8基因自2002年被Rodriguez等[2]发现以来,有越来越多的学者研究该基因在恶性肿瘤中的作用。发现基因表达沉默这一现象在脑胶质瘤、结肠癌、甲状腺癌、胰腺癌、肺癌、宫颈癌中普遍存在[3-6]。随着对SLC5A8基因研究的深入,发现在结直肠癌、甲状腺癌、肺癌、胃癌、食管癌等恶性肿瘤的发生中存在着基因的表观遗传学改变现象,比如抑癌基因的甲基化,从而抑制了抑癌基因的表达。SLC5A8基因作为目前的候选抑癌基因,是溶质转运体家族第5家族第8成员。目前鲜有对于抑癌基因SLC5A8在乳腺癌中的作用机制及与乳腺癌的发生、发展关系方面的研究。本研究通过对乳腺癌及其癌旁组织中SLC5A8基因甲基化及SLC5A8转录表达水平的实验性研究,以明确SLC5A8基因在乳腺癌中的表达情况,进一步探究其作用机制,为后一步的基因治疗和预后评估奠定实验理论基础。
1 材料与方法
1.1 材料
2017年5月至2019年12月收集河北大学附属医院基本外科行乳腺癌改良根治术的32例乳腺癌患者标本,将肿瘤标本及癌旁正常组织标本分为实验组和对照组。患者术前未行任何治疗,术后病理回示为恶性,所有患者均为女性,年龄42~56岁,平均为(49.00±5.23)岁。
Omega E.Z.N.A.TM总RNA提取试剂盒、反转录试剂盒、实时荧光定量PCR反应试剂盒、组织DNA提取试剂盒、琼脂糖、氯仿、无水乙醇、5×RNA loading buffer、微量高速冷冻离心机、全波长酶标仪、全自动凝胶成像仪、实时荧光定量PCR仪、实验所用引物由上海英潍捷贸易有限公司提供,引物序列见表1。

表1 基因引物序列及扩增片段
1.2 实验方法
1.2.1 SLC5A8基因甲基化检测
采用甲基化特异性PCR法检测,按照DNA提取试剂盒说明书步骤提取样本组织DNA并修饰,将对应引物进行亚硫酸氢盐修饰DNA样本的PCR扩增。PCR反应条件:50 ℃ 2 min;95 ℃中预变性2 min;95 ℃中15 s;60 ℃中30 s,如此40次循环。最后对反应产物进行琼脂糖凝胶电泳照相。
1.2.2 SLC5A8基因转录表达检测
运用RT-PCR法检测SLC5A8基因的转录表达。实验按照RNA提取试剂盒说明提取总RNA,对RNA总量进行定量配平后对其行反转录。检测引物序列特异性后,分别用以上2对引物行RT-PCR。PCR反应条件:50 ℃ 2 min;95 ℃预变性2 min;95 ℃ 15 s;60 ℃ 30 s,如此40次循环后,对反应产物琼脂糖凝胶电泳照相。
1.3 统计学处理

2 结果
2.1 乳腺癌和乳腺癌旁正常组织SLC5A8基因甲基化检测
在乳腺癌组织中SLC5A8基因发生甲基化20例(62.50%),而在乳腺癌旁正常组织的基因发生甲基化7例(21.87%)。癌旁正常组织的基因发生甲基化的概率相较癌组织的基因甲基化的概率降低,2组差异有统计学意义(P<0.05),见表2。
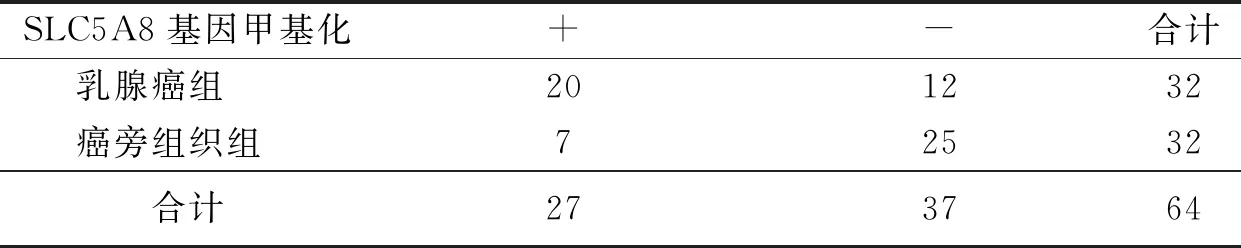
表2 乳腺癌及癌旁组织SLC5A8基因甲基化检测
2.2 乳腺癌及癌旁组织SLC5A8基因扩增后凝胶电泳结果
乳腺癌及乳腺癌旁组织的SLC5A8基因RT-PCR结果的凝胶电泳如图1,32例乳腺癌组织中SLC5A8基因明显过表达占43.75%(14/32),而乳腺癌旁正常组织该基因71.25% (26/32)出现过表达现象,提示癌组织中SLC5A8基因的过表达水平明显低于乳腺癌癌旁正常组织水平,2组差异有统计学意义(P<0.05),见表3。
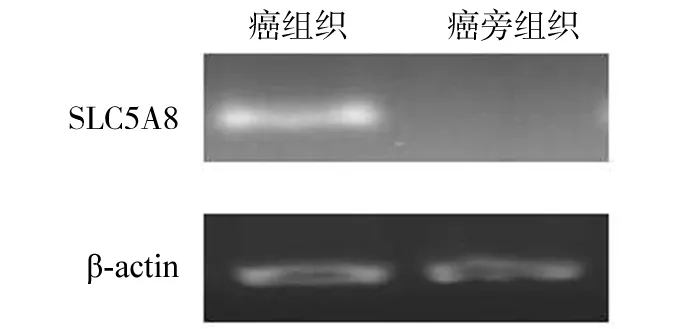
图1 SLC5A8基因扩增后凝胶电泳

表3 乳腺癌及癌旁组织SLC5A8基因过表达检测
2.3 乳腺癌组织SLC5A8基因甲基化与转录表达
在32例乳腺癌组织中,SLC5A8基因甲基化阳性的有20例标本,基因转录表达者6例(30.00%);基因未转录表达者14例(70.00%)。癌组织中基因甲基化阴性有12例标本,基因转录表达者7例(58.33%);基因未转录表达者5例(41.66%)。无序变量相关性分析显示,乳腺癌组织中SLC5A8基因甲基化和SLC5A8基因转录表达两者一致性较差,反而具有一定的反向一致性,这也验证了乳腺癌组织SLC5A8基因甲基化与基因的转录表达不是完全一致的,癌组织基因甲基化阳性标本的基因转录水平相对较低,并且要弱于基因甲基化阴性标本的基因转录水平,故2组差异有统计学意义(P<0.05),见表4。

表4 乳腺癌SLC5A8基因甲基化与基因转录表达关系
3 讨论
自2002年Rodriguez等[2]发现SLC5A8基因以来,很多学者发现基因表达沉默这一现象在脑胶质瘤、结肠癌、甲状腺癌等恶性肿瘤中普遍存在[3-6]。进一步揭示了SLC5A8基因表达下降的机制主要与该基因外显子1中CPG岛广泛甲基化有关[7]。进一步研究得出,SLC5A8基因与染色体组蛋白脱乙酰基酶主要通过丁酸相关联[8]。在结肠肠腔中的细菌经过一定作用将膳食纤维分解成丁酸,使其浓度升高。结肠上皮细胞的SLC5A8基因表达增高,SLC5A8基因可以将丁酸转运到细胞内,同时他作为组蛋白脱乙酰基酶抑制剂,在上皮细胞的癌前病变方面可以预防的同时还对肿瘤细胞的凋亡有促进作用[9-12]。SLC5A8基因在其他脏器中预防细胞发生癌变则主要是通过与抗凋亡蛋白相互作用实现的[13]。二氯乙酸也是该基因转运体的作用底物中的一个,联合应用5杂氮胞苷,同样具有明显促进细胞凋亡的作用[14]。SLC5A8基因广泛表达于人体的各个组织器官,属于候选抑癌基因,对细胞有保护作用。肿瘤在发生发展过程中选择性地使SLC5A8基因表达下降,维持自身增殖状态,是最终避开凋亡的重要机制。如果SLC5A8基因的表观遗传学变化是乳腺癌组织发生、发展的重要机制,这将在未来对乳腺癌的分子生物学治疗具有重要意义。对乳腺癌中SLC5A8基因的甲基化及转录水平进行研究,以期发现SLC5A8基因甲基化和基因转录表达水平降低与乳腺癌关联性,研究发现实验组的甲基化阳性率显著高于对照组甲基化阳性率(P<0.05)。实验组中甲基化阳性的癌组织基因转录表达率较甲基化阴性的癌组织基因转录表达率显著降低(P<0.05),癌组织SLC5A8基因甲基化与基因的转录表达不是完全一致的,癌组织基因甲基化阳性标本的基因转录水平相对较低,并且要弱于基因甲基化阴性标本的基因转录水平,进一步表明在乳腺癌的发生、发展中可能不止存在表观遗传学现象,还存在其他事件。上述结果和概括得出SLC5A8基因的甲基化是乳腺癌中普遍存在的一种现象,并可能是乳腺癌演变过程的重要一步,并且是SLC5A8基因转录水平降低的主要因素。这为进一步研究SLC5A8基因的表观遗传学变化在乳腺癌发生、发展中的作用机制提供了重要依据,并且对乳腺癌的分子生物学治疗提供可靠证据。
猜你喜欢
健康管理(2016年2期)2016-05-30 21:36:03
湖北农业科学(2015年12期)2015-08-13 08:51:53
安徽理工大学学报·自然科学版(2015年1期)2015-07-21 14:24:00
中国医疗美容(2015年1期)2015-07-12 10:06:52
安徽农学通报(2015年8期)2015-07-02 01:37:32
医学研究杂志(2015年9期)2015-07-01 17:27:46
现代检验医学杂志(2015年2期)2015-02-06 02:00:48
沈阳医学院学报(2014年4期)2014-12-27 13:44:30
遗传(2014年3期)2014-02-28 20:58:49
实验动物与比较医学(2014年5期)2014-02-28 14:53:16
