
杉木成熟林细根形态与功能特征的海拔梯度变异特点
2020-03-13 05:20李爱琴张莎莎王会荣张丹丹赵晓雅关梦冉徐小牛
生态学报 2020年2期
李爱琴,张莎莎,王会荣,张丹丹,赵晓雅,关梦冉,徐小牛
安徽农业大学林学与园林学院, 合肥 230036
细根(d≤ 2 mm)是植物吸收水分和养分的主要器官,其快速生产和周转率对生态系统碳收支平衡、养分循环过程发挥着重要作用[1- 2]。细根也是植物对环境变化响应最敏感和最活跃的部分[3],其形态参数和养分含量不仅影响细根生理生态特性[4- 5]、反映林地养分有效性[6],还表达了植物生存环境的变化信息[7]。在全球气候变化日益加剧的时代背景下,研究其性状指标对环境变化的适应机制有重要现实意义[8]。
海拔梯度变化作为一种自然地理变化,影响着森林土壤理化性质[9]、植被分布[10]以及因植被密度不同导致的凋落物量变化[11],为细根对环境的响应提供了合适的研究场所。近年来不少学者在海拔梯度上开展了研究,初步表明不同海拔植被细根生物量与性状指标有明显的垂直变化规律[12-15]。同时土壤水分状况是海拔梯度上变化最明显的因子之一,土壤水分的多少成为细根生长的决定性因素[16]。但受地理条件的限制,这些研究的对象集中于不同海拔的不同自然林群落,针对不同海拔梯度上同一植被类型的细根特征研究较少,对人工林细根在海拔上的响应研究更是鲜见报道。
杉木(Cunninghamialanceolata)是中国南方集体林区重要的优质、速生、高产的用材树种,在我国人工林资源中占据重要地位[17]。对杉木林细根在不同林龄[18]、林分结构[19- 20]等方面的特征研究已有初步进展,但对其细根在海拔梯度上的变化规律尚不清晰。鉴于以上原因,对同一区域不同海拔同类植被细根的研究,有助于了解细根沿海拔的变化状况。本文针对安徽大别山马鬃岭杉木人工林细根生物量、形态指标和碳氮含量的海拔梯度变异特点进行了探讨,目的为深入了解林木细根对环境变化的适应机制提供理论依据,为大别山区杉木林科学经营管理乃至揭示北亚热带地区杉木人工林生长规律及其生产力形成机制提供基础资料。
1 研究地区与研究方法
1.1 研究区概况
本研究地位于安徽省金寨县西南天马国家自然保护区(31°10′—31°20′N和115°31′—115°50′E),总面积34.7 km2,试验区内群山林立、沟谷纵横、地形起伏较大,30°—40°陡坡较多。气候类型属亚热带湿润季风气候,四季分明,年均气温13—15 ℃,年均降水量约1510 mm,年蒸发量1266 mm,相对湿度80%,无霜期210 d。调查区岩层主要是寒武纪前的变质岩,成土母质主要来自花岗岩风化物,土壤质地为沙壤至轻粘壤土,偏酸性。土壤垂直分布规律为:海拔800 m以下为山地黄棕壤,800 m以上为山地棕壤,1400 m以上局部地区有山地草甸土和沼泽土存在[21]。森林类型丰富,森林覆盖率达94%,植被分布垂直变化明显。区内主要树种有杉木、青冈栎(Cyclobalanopsisglauca)、黄山松(Pinustawanensis)、马尾松(Pinusmassoniana)、紫楠(Phoebeshearer)等。
1.2 研究方法
1.2.1样地设置
根据研究区内杉木人工林生长和分布情况,选择26—35 a生的成熟林分,于2018年7月,按750 m、850 m、1000 m和1150 m四个海拔设置典型调查样地,每个海拔高度设置3个基本特征一致的重复样地,每个样地面积为20 m × 20 m。不同海拔梯度样方内杉木成熟林基本情况见表1。
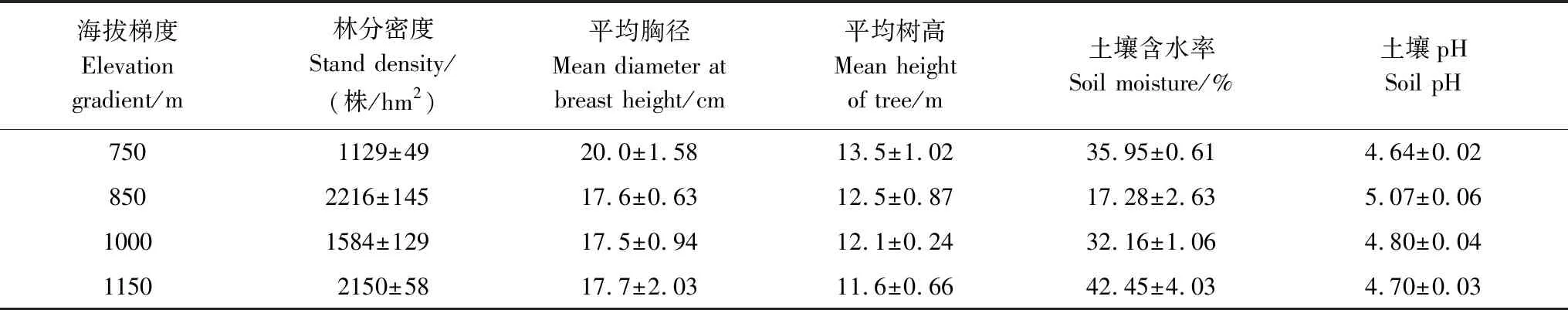
表1 杉木林调查样地基本情况
以上数据均为平均值±标准误
1.2.2样品采集与处理
根系样品:在每个样地内随机选取9个样点,将地表凋落物清理干净后,用内径5.50 cm的土钻按0—10 cm、10—20 cm和20—30 cm土层分层取样,放入自封袋中标记编号,用冰盒低温保存带回实验室。将带有根样的土芯放入低温(1—2 ℃)去离子水浸泡1 d,随后用60目孔径的分样筛反复淘洗过滤,使根系与杂质和泥土分离。洗净后用镊子、游标卡尺等工具分拣出直径≤ 2 mm的所有目标树种活根(根据根系外形、颜色、气味、根皮、弹性等区分),剔除草根。将获得的活根依次装入标记好的自封袋中放入冰箱保存待测。
土壤样品:在根系取样的同时,在每个样地随机选取3个点,每个点采用多点混合分层取样法,即:用土钻(内径为3 cm)按0—10 cm、10—20 cm和20—30 cm土层取样,放入自封袋中标记编号,共计108份土壤样品,与根系一同带回实验室待测。
1.2.3细根样品指标测定
将获得的样品用Microtek数字扫描仪(MRS- 600A3LED)进行扫描,保存其扫描图像,用配套根系分析系统WinRhizo进行分析得到平均直径、根长、表面积、体积等指标。待所有根系扫描完成后将根样装入信封标记,于65 ℃烘箱烘干至恒重(72 h),测定各样品生物量(mg/cm3)。经研磨机粉碎后使用元素分析仪(EA3000 CNHSO Analysis,Euro Vector)测定全碳(C),全氮(N)。将各根样长度、表面积、体积分别除以取样土体体积计算出根长密度(cm/cm3)、表面积密度(cm2/cm3)、体积密度(mm3/cm3)等细根特征指标;将各根样细根长度除以其生物量得到各根样的比根长(m/g)。
1.2.4土壤含水测定
采用烘干法即在105 ℃条件下烘干至恒重测定土壤含水率,计算公式为:土壤含水率(SWC,%)=(鲜土质量-干土质量)/鲜土质量×100%。
1.3 数据处理
采用双因素方差分析法分析不同海拔梯度和土壤深度对细根形态和养分含量的影响,并用Duncan多重比较进行差异显著性检验;采用皮尔逊(Pearson)相关分析进行细根各指标间相关性检验;取每个样地内的细根特征指标和土壤含水的平均值,采用回归分析研究细根各指标与土壤含水率的相关关系。所有数据处理、绘图、统计分析在Excel 2016和SPSS 20.0中完成。
2 结果与分析
2.1 不同海拔细根生物量与形态指标特征
随海拔梯度增加,0—30 cm土层杉木细根的平均生物量、平均根长密度、平均比根长、平均表面积密度、平均体积密度均呈先减少后增加的变化趋势(图1)。双因素方差分析表明(表2),海拔对细根根长密度和表面积密度有极显著影响(P<0.01),对体积密度有显著影响(P<0.05),对生物量和比根长无显著影响(P>0.05);土层对细根比根长无显著影响(P>0.05),对其余形态指标均有极显著影响(P<0.01);海拔与土层的交互作用只对细根表面积密度有显著影响(P<0.05),对细根生物量、根长密度、比根长、体积密度均无显著影响(P>0.05)。
2.1.1生物量
同一土层不同海拔细根生物量存在差异,0—10 cm和10—20 cm土层细根生物量均为海拔850 m<1000 m<1150 m<750 m(图1),20—30 cm土层细根生物量为海拔1000 m<850 m<1150 m<750 m。同一海拔不同土层的细根生物量随土层深度增加表现出减少趋势。各海拔土层间差异性水平不同,其中,海拔1000 m处,三个土层间差异显著(P<0.05),海拔1150 m处,0—10 cm土层与20—30 cm土层差异显著(P<0.05)。20—30 cm土层间细根生物量在0—30 cm土层细根总生物量中贡献最小,在750、850、1000、1150 m处分别仅占0—30 cm细根总生物量的18.5%、20.9%、16.8%、17.0%。

表2 海拔梯度和土壤层次对细根生物量,形态特征,C、N含量影响的方差分析
*,P< 0.05; **,P< 0.01
2.1.2根长密度
随海拔梯度增加,0—30 cm土层细根平均根长密度在1150 m处最大,850 m处最小,两者之间差异显著(P<0.05)。三个土层细根根长密度均为海拔850 m<1000 m<750 m<1150 m(图1)。同一海拔梯度细根根长密度随土层深度增加呈明显减少趋势。海拔750 m处10—20 cm和20—30 cm土层细根根长密度差异不显著(P>0.05),但都与0—10 cm土层差异显著(P<0.05)。同一土层不同海拔梯度间,850 m和1000 m海拔处3个土层间根长密度差异极显著(P<0.01)。海拔1150 m处0—10 cm土层与20—30 cm土层间细根根长密度差异显著(P<0.05)。
2.1.3比根长
0—30 cm土层细根平均比根长在海拔1150 m最大,850 m最小,两者之间差异显著(P<0.05)。同一土层细根根长密度在不同海拔间的变化趋势存在差异(图1)。0—10 cm和20—30 cm土层细根比根长为海拔850 m<750 m<1000 m<1150 m,10—20 cm土层细根比根长为海拔750 m<850 m<1000 m<1150 m。同一海拔不同土层间细根比根长差异均不显著(P>0.05),且在土层间的变化趋势不一致,海拔850 m细根比根长随土层深度增加呈减少的趋势,其他海拔比根长均随土层深度增加呈先减少后增加的趋势。
2.1.4表面积密度
测定海拔范围内,0—30 cm土层细根平均表面积密度在海拔1150 m处最大,750 m次之。0—10 cm土层细根表面积密度为海拔850 m<1000 m<750 m<1150 m,10—20 cm和20—30 cm土层细根表面积密度均为海拔850 m<750 m<1000 m<1150 m(图1)。同一海拔不同土层细根表面积密度变化趋势基本一致,均为20—30 cm<10—20 cm<0—10 cm。海拔750 m、850 m和1000 m不同土层间差异显著(P<0.05);海拔1150 m处0—10 cm与10—20 cm土层间细根表面积密度在差异不显著,但都与20—30 cm土层差异显著(P<0.05)。
2.1.5体积密度
由图1可以看出,细根体积密度在海拔1150 m最大,海拔750 m次之。0—10 cm和10—20 cm土层细根体积密度为海拔850 m<1000 m<750 m<1150 m,20—30 cm土层细根体积密度在海拔750 m最大,海拔850 m最小。同一海拔细根体积密度随土层深度增加呈减少的趋势,海拔750、850和1150 m处,0—10 cm土层细根体积密度显著高于20—30 cm土层,海拔1000 m处各土层间差异显著(P<0.05)。
2.2 不同海拔梯度细根碳氮含量变异特征
随海拔梯度增加,细根平均C、N含量呈先增加后减少的变化趋势,平均C/N比呈先减少后增加再减少的趋势。双因素方差分析结果显示(表2):海拔对杉木细根C含量有极显著影响(P<0.01),对N含量有显著影响(P<0.05),对C/N比无显著影响(P>0.05);土层对细根N含量、C/N比有极显著影响(P<0.01),对C含量无显著影响(P>0.05);海拔与土层的交互作用对C、N含量和C/N比均无显著影响(P>0.05)。
2.2.1碳含量
海拔梯度上0—30 cm土层细根平均C含量在850 m最大,1000 m次之。0—10 cm土层细根C含量为海拔1150 m<1000 m<850 m<750 m,10—20 土层细根C含量为海拔1150 m<750 m<850 m<1000 m,20—30 cm细根C含量为海拔1150 m<750 m<1000 m<850 m(图2)。同一海拔不同土层细根C含量的变化趋势一致,海拔750 m、850 m、1000 m、1150 m细根C含量均随土层加深呈先减少后增加的趋势,各海拔不同土层间细根C含量差异不显著(P>0.05)。
2.2.2氮含量
海拔梯度上0—30 cm土层细根平均N含量在850 m最大,1150 m最小,两者之间差异极显著(P<0.01)。0—10 cm和10—20 cm土层细根N含量为海拔1150 m<750 m<1000 m<850 m,20—30 cm土层细根N含量为海拔1150 m<1000 m<750 m<850 m。同一海拔不同土层细根N含量随土层加深呈减少的趋势(图2)。750 m和850 m海拔细根N含量变化基本一致,即10—20 cm与20—30 cm差异不显著(P>0.05),但都与0—10 cm差异显著(P<0.05)。1000 m和1150 m处土层间无显著差异(P>0.05)。
2.2.3C/N比
由图2可以看出,0—30 cm土层细根平均C/N比在海拔1000 m最大,海拔850 m最小。0—10 cm土层细根C/N比在海拔750 m最大,10—20 cm和20—30 cm土层细根C/N比在海拔1000 m最大。同一海拔不同土层下,细根的C/N比均随土层深度增加呈缓增趋势。其中,海拔1000 m,0—10 cm与10—20 cm、20—30 cm土层细根C/N比均差异显著(P<0.05)。海拔1150 m处0—10 cm与20—30 cm土层间细根C/N比存在显著差异(P<0.05)。
2.3 细根形态和功能特征与土壤水分的关系
对杉木细根生物量、形态指标、碳氮含量与土壤含水量进行Pearson相关分析(表3)。结果表明:土壤含水率与细根生物量、根长密度、比根长呈极显著正相关关系(P<0.01),与表面积密度,体积密度呈显著正相关关系(P<0.05);与细根C含量、C/N比呈负相关关系,但均不显著(P>0.05),与细根N含量呈正相关关系,也不达不到显著水平(P>0.05)。对不同海拔梯度杉木细根在不同土层深度中各形态指标与土壤含水率进行回归分析(图3)。结果表明:细根各形态指标随土壤含水率的升高呈二项式型增长趋势,但拟合度均不高,其中根长密度的回归方程拟合度相对较高(R2=0.445),与体积密度的拟合度最低(R2=0.182)。
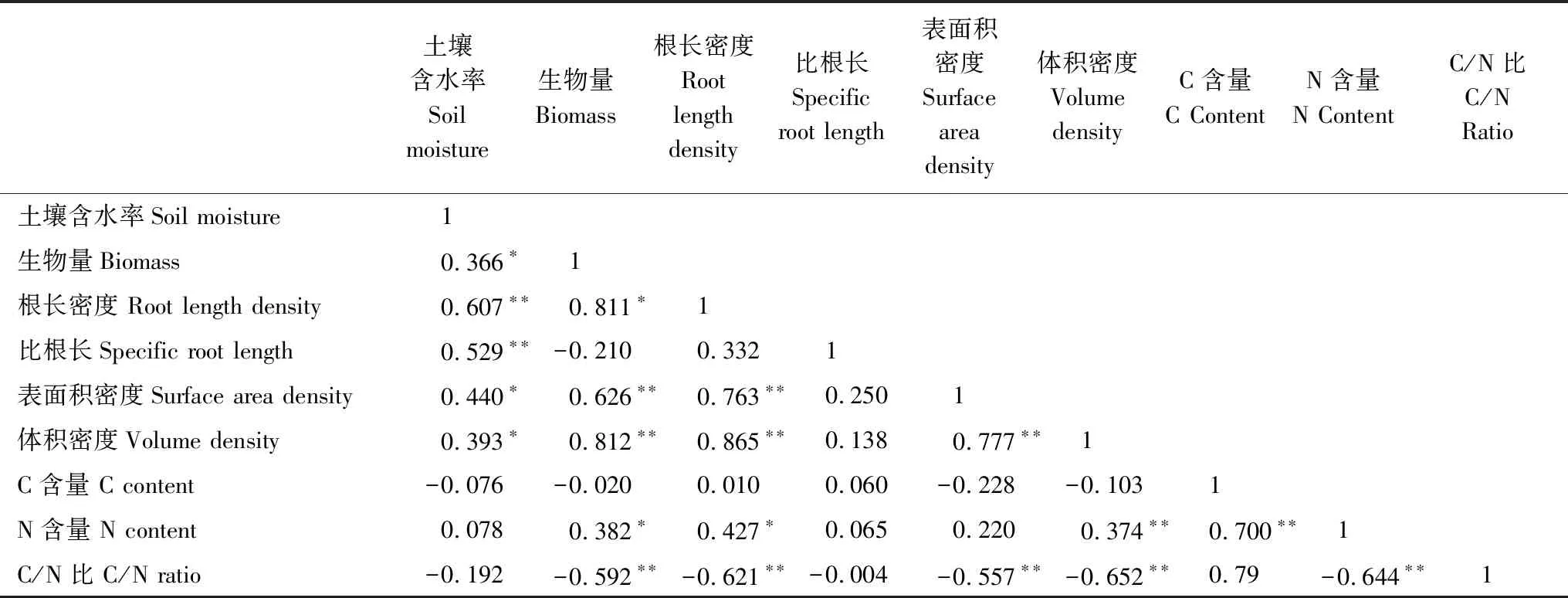
表3 杉木人工林细根形态和功能特征相关性
*,P< 0.05; **,P< 0.01

图3 杉木细根各指标与土壤含水率的关系Fig.3 Regression analysis of fine root indices and soil moisture in Chinese fir plantation
3 讨论
3.1 不同海拔杉木细根生物量与形态指标的变异特征
本研究结果表明马鬃岭天马自然保护区杉木成熟林0—30 cm土层细根在不同海拔梯度的平均生物量为3.13 mg/cm3(313.70 kg/m2),介于Jackson等[22]综述的世界上有关亚热带森林细根的生物量范围内(1.1—5.8 t/ha-1)。略高于刘聪[23]对亚热带杉木细根生物量的研究结果(268.75 g/m2),但远低于权伟等[15]对武夷山针叶林细根生物量(7.45 kg/m3),本研究取样时间为根系生长旺盛季,植被所处生态环境更优质可能造成了生物量估算值偏高于刘聪的研究结果,而武夷山的针叶林的细根主要由竹根组成是造成本研究与其差异过大的重要原因。研究发现各海拔梯度细根生物量存在差异,这种差异与其所在土壤类型、坡向、坡度、气候带、以及自身遗传特性等有关[24]。郭炳桥[25]对黄山松细根在海拔梯度(1400—2000 m)的适应研究发现不同海拔细根生物量存在差异,最大值和最小值分别出现在1800 m和1400 m。本研究中海拔750 m细根生物量最高,这是因为低海拔处林分综合生境适宜,根系分布性好。另有关人工林研究中发现细根生物量随林分密度增加呈现出逐渐减少的趋势[26- 27],850 m细根生物量最低,林分密度最高,这是因为高密度林分中,林冠挤压遮挡,导致林木对光照利用率降低,分配到地下部分的地下产物减少[28];林下根系生长空间压缩,竞争力增加,导致生物量偏低,因此本研究生物量出现先减少后增加的趋势。
构型可塑性的响应是植物适应外界环境变化的一种重要模式[29]。本研究发现细根表面积密度在海拔1150 m显著高于低海拔,与Girardin等[30]在亚马逊海拔1000 m以上得出的森林细根表面积明显大于Metcalfe等[31]在低地所获取的数据的结果一致,也与燕辉[32]在研究中指出的土壤含水量与细根各形态参数之间存在不同程度的正相关性的结果一致。这是因为细根表面积作为与土壤直接接触面,受土壤水分影响明显。有研究显示当生物量低时,其比根长更长,以便更好的吸收水分和养分,本研究显示细根比根长与生物量呈负相关关系但不显著,与Alvarezuria等[33]对阿尔卑斯山3种树线8种树木的研究中,针叶树种比根长并未随海拔梯度的变化发生明显变化的结果相一致,可见,细根比根长与生物量之间的具体联动机制尚需成为之后的研究关注点。在海拔850 m细根各形态指标均较低,与此处林地坡度较陡,土壤含水率和温度低,限制了细根的高速周转有关。Vanguelova 等[34]指出在偏酸性土壤条件下根系生长的相关指标会更高,也印证了本研究区850 m海拔处土壤pH相对较高的现象。
森林土壤具有高度的空间异质性[35],不同土层之间养分差异明显,使得根系在不同土层间可以表现出不同的生理活性[36-37]。本研究结果显示细根生物量随土层深度增加呈减少趋势,这与韩畅[8]、郭忠玲[38]、辛福梅[39]的研究结果一致。这源于根系趋肥特点,表层土壤腐殖质丰富,温度适宜,水分充足,养分含量高,有利于获取充足的水分和养分[40-41]。比根长是反映细根吸收能力的最直接指标,本研究中,0—10 cm到10—20 cm土层细根比根长呈降低趋势,主要与土壤含水极显著相关。土层深度20 cm以下,各海拔细根比根长20—30 cm均高于10—20 cm,取样时发现,研究区土质层较薄,石砾增加,土壤孔隙度增大,根系盘结,故细根根长相对增加。土壤含水与细根根长密度、表面积密度和体积密度呈显著正相关,故随着土层的加深,土壤含水率降低,细根根长密度,表面积密度,体积密度也降低。
3.2 不同海拔细根碳氮养分含量的变异特征
植物根系的生理生态功能除与其形态特征有关外,还与根系组织中的C和N浓度有着密切的联系[2]。本研究杉木细根C、N含量、C/N比平均值分别为329.70 g/kg、10.30 g/kg、32.01,其中C含量、C/N比平均值低于中国植物细根C含量、C/N比平均值(473.9 g/kg、59.15)[42],但细根N含量平均值高于中国植物细根N含量平均值(9.16 g/kg),这可能是亚热带地区长期大气氮沉降,使土壤中N有效性增加,从而影响到植物细根N含量。此现象与Kerkhoff等[43]研究发现的亚热带和热带地区大部分表现出高N结果一致。研究中杉木细根C、N含量和C/N比均显示高海拔低于低海拔,这与陈晓萍等[14]对于武夷山不同海拔黄山松细根的研究结果基本一致,主要是随着海拔的升高,温度降低,限制了植物体养分和水分的吸收[44]。
林木根系分布深度决定了林木对地下空间的占有量和对土壤水分和养分的利用率,进而影响地上植被产量[45]。本研究垂直分布中细根养分含量存在差异,细根N含量变化与刘佳等[38]研究结果相似,随土层的加深逐渐降低,这可能与土层加深土壤温度降低有关,马玉珠等[43]研究也表明植物细根N含量受温度影响显著。细根C含量在土层间差异不显著也跟土壤养分含量、树种生物学特性等综合因素有关。研究中发现不同海拔、不同土层杉木细根C/N比差异均不显著,这是因为碳氮作为重要结构性成分,紧密相连,二者在积累与消耗的过程中具有相对稳定性[46]。土壤各层次细根养分含量的差异可反映出细根功能转换及土壤资源有效情况。
4 结论
山地海拔梯度的变化造成的环境异质性对植物根系形态和功能特征产生重要影响[14]。本研究分析了不同海拔梯度上杉木成熟林细根生物量、形态特征和碳氮含量的变异特征,结果表明:随海拔梯度增加,细根生物量和形态特征均呈先减少后增加的趋势;细根C含量和N含量呈先增加后减少变化趋势,C/N比呈先减少后增加再减少变化趋势;细根C含量与各形态指标没有相关性,N含量与生物量、根长密度和体积密度极显著正相关,C/N比与生物量、根长密度、表面积密度和体积密度极显著负相关;土壤含水率对细根形态特征指标有重要影响。不同海拔梯度间杉木细根的形态特征和化学含量均存在差异,这种差异不仅与其自身的遗传特性有关[39],更是植物细根调整自身生理生态状态以更好适应生存环境的结果。因此研究海拔梯度上杉木细根的形态和功能特征的变异特征可以更好的了解森林植被与环境因子之间的关系,同时对研究环境梯度上森林生态系统格局和功能差异具有重要借鉴。
猜你喜欢
小小说月刊·下半月(2021年8期)2021-09-05
绿色科技(2021年5期)2021-04-09
农民致富之友(2020年8期)2020-05-11
乡村科技(2019年21期)2019-02-21
四川农业大学学报(2018年3期)2018-09-04
动漫界·幼教365(大班)(2018年3期)2018-05-14
现代园艺(2017年23期)2018-01-18
现代园艺(2017年23期)2018-01-18
中央民族大学学报(自然科学版)(2017年4期)2017-06-11
湖南大学学报·自然科学版(2016年12期)2017-05-12
