
MTB和BCG诱导中性粒细胞胞外捕获网形成方法的建立
2020-03-11 07:12
中国现代医药杂志 2020年1期
结核病是由人类特有的胞内病原体—结核分枝杆菌引起的,并通过呼吸道在人与人之间传播的一种慢性传染性疾病。结核分枝杆菌具有特殊的细胞壁结构,含丰富的脂质,对外界环境和药物有很强的抵御能力。细胞内生长的结核分枝杆菌致病机制已经研究得很透彻,而胞外生长的结核分枝杆菌引起的病理变化则很少涉及[1~3]。长期以来,人们围绕结核病免疫相关的T淋巴细胞(尤其是CD4+的Th1细胞)、巨噬细胞和树突状细胞(dendritic cells,DC)进行了大量研究,并取得了一些进展。目前,多数研究者肯定的观点是,Th1型免疫应答在结核病的适应性免疫应答中起关键作用,是介导结核病保护性免疫的主要群体;巨噬细胞作为机体抗MTB感染的主要免疫效应细胞,在结核病的保护性免疫应答中也发挥着重要作用。但是,对于参与结核病免疫的另一个重要细胞群体,也是MTB感染后最先由外周血迁移至感染病灶的细胞—中性粒细胞的研究却明显滞后[4]。本研究拟通过结核分枝杆菌和卡介苗诱导中性粒细胞形成NETs(neutrophil extracellular traps)结构,探讨中性粒细胞在结核感染过程中所扮演的角色,揭示卡介苗预防结核分枝杆菌感染的可能机制。
1 材料与方法
1.1 材料 结核分枝杆菌国际标准株、BCG菌种由北京结核病胸部肿瘤研究所提供,胎牛血清(FBS)、RPMI-1640、Middlebrook 7H9培养基购自美国Sigma公司,Sytox® Green 试剂盒购自美国Eugene公司,人中性粒细胞提取试剂盒购自天津市灏洋生物制品科技有限责任公司,抗-H3Cit、抗-MPO抗体购自Abcam公司,Quant-iTTMPicoGreen ® dsDNA Reagent and Kits购自ThermoFisher公司。人外周血标本来自本院健康体检志愿者,无外伤、感染病史,HBV、HCV、HIV均阴性,并签署知情同意书。
1.2 方法
1.2.1 细菌培养 取标准株H37Rv和BCG接种于5ml无菌7H9液体培养基中,37℃摇床恒温箱中培养3~4周,摇床速度为100~120转/h,吸取100μl菌液接种于5ml新鲜7H9液体培养,继续摇床培养7~10d。取菌液100μl重悬于PBS中,调整至麦氏浓度0.5。
1.2.2 中性粒细胞分离 取10ml离心管一支,将健康志愿者EDTA抗凝血4ml小心加到4ml中性粒细胞分离液之上。3 000r/min,水平离心30min,离心后吸取中性粒细胞层后加入红细胞裂解液,充分裂解后离心,3 000r/min,30min后取中性粒细胞层加入PBS洗涤。用含5%FBS的RPMI-1640培养基重悬后进行细胞计数。
1.2.3 NETs诱导 加入0.5ml中性粒细胞重悬液(含细胞数4×105个)于24孔细胞培养板,放入多聚赖氨酸处理的无菌玻片,5%的CO2培养箱中1h静置处理细胞,对照组、MTB组、BCG组分别加入PBS、MTB、BCG各0.5ml,各组设平行3组。5% 的CO2培养箱中3h后收集细胞上清,用于检测游离DNA。各孔加入0.5ml 4%多聚甲醛,4℃固定过夜,用于免疫荧光染色。
1.2.4 NETs检测 免疫荧光染色:从6孔板中取出细胞爬片,PBS洗涤3次,0.5%Triton X-100透化1min,PBS洗涤3次,加入5%FBS于37℃湿盒中封闭30min。加入一抗(抗-H3Cit 1∶400,抗-MPO 1∶400)于37℃湿盒中封闭1h。PBS洗涤3次后加入二抗(抗-H3Cit 1∶1000,抗-MPO 1∶1000)于37℃湿盒中封闭1h。PBS洗涤3次后加入DAPI避光染色10min,加封片剂封片。激光共聚焦显微成像仪检测NETs。
1.2.5 游离DNA定量检测 按Quant-iTTMPicoGreen ®dsDNA Reagent and Kits试剂盒说明书进行操作,倍比稀释标准品制作标准曲线,激发光波长480nm,发射光波长520nm,荧光检测仪检测荧光信号并记录结果。
1.3 统计学分析 采用SPSS 19.0统计软件进行数据分析,计量资料采用均数±标准差(±s)表示,采用t检验,以P<0.05为差异有统计学意义。
2 结果
2.1 NETs的激光共聚焦显微成像 激光共聚焦显微成像分析发现:中性粒细胞在BCG和MTB的刺激下,可见丝状或环状DNA骨架结构,H3Cit和MPO附着于DNA骨架结构上。中性粒细胞的体积增大,且产生环状或丝状骨架结构的细胞比例增多。BCG组和MTB组标记H3Cit和MPO颗粒物的荧光强度较对照组明显增强,即DNA骨架结构上附着的H3Cit和MPO颗粒数量增多,见图1。
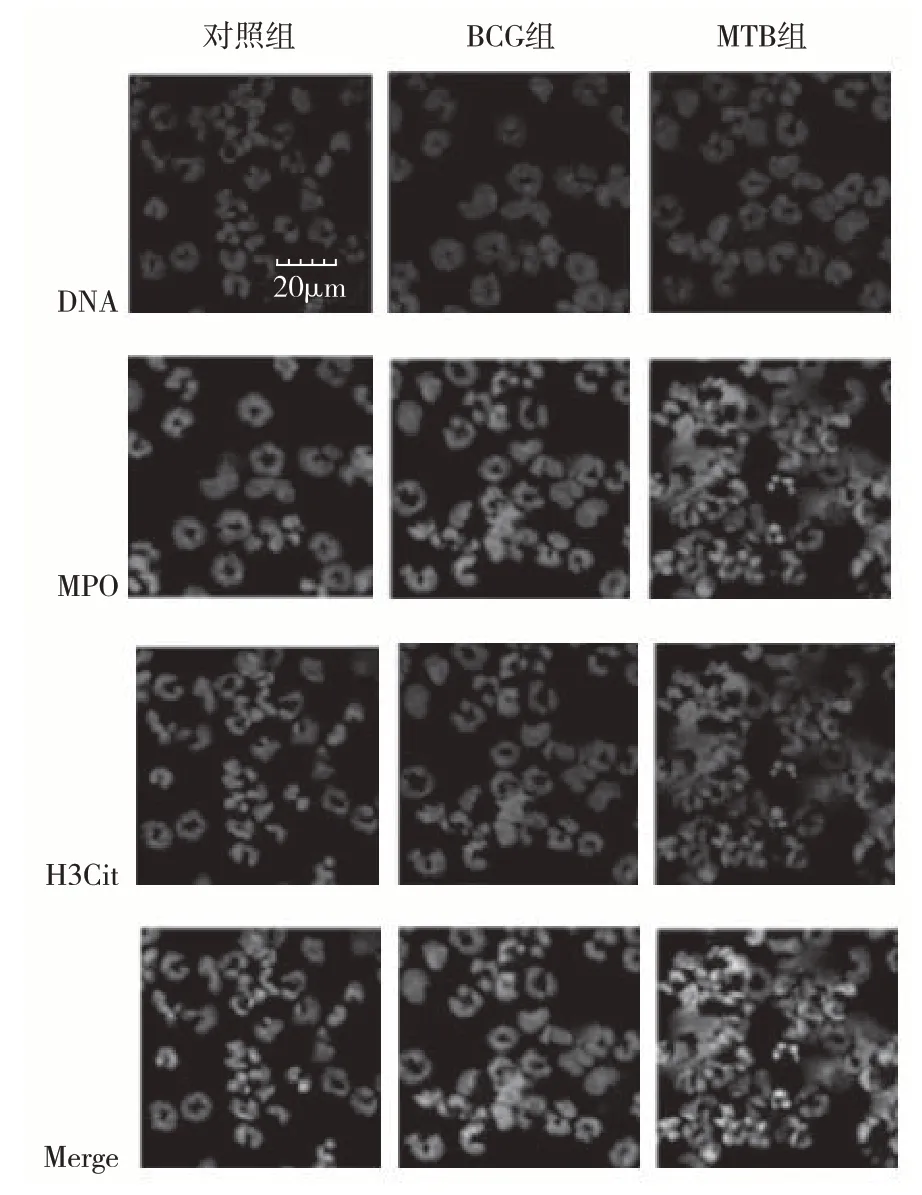
图1 NETs免疫荧光染色
2.2 游离DNA含量 中性粒细胞经BCG和MTB刺激后,上清中游离DNA含量较对照组显著增高(P<0.05),MTB刺激组游离DNA含量明显高于BCG刺激组(P<0.05)。该结果与免疫荧光染色结果一致,证明中性粒细胞经BCG和MTB刺激可形成NETs,且MTB刺激效率明显强于BCG,见图2。
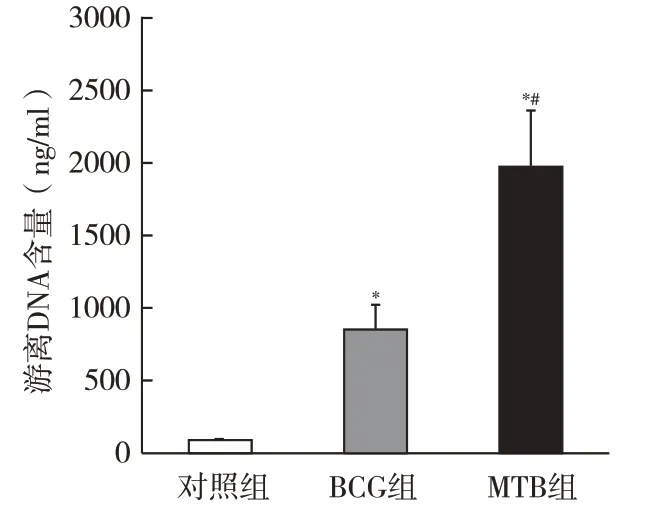
图2 游离DNA含量
3 讨论
尽管中性粒细胞占外周循环白细胞数量的50%~80%,但其吞噬作用一直被低估,免疫学的研究重点主要集中在单核吞噬细胞如单核细胞和巨噬细胞。中性粒细胞通过直接吞噬细菌、脱颗粒及细胞因子的分泌,如肿瘤坏死因子-α(TNF-α)和白介素-1(IL-1)来发挥先天性免疫作用。2004年Brinkmann等[5]首先报道了中性粒细胞活化后会在胞外形成类似网状的结构—中性粒细胞胞外捕获网,并认为这是中性粒细胞发挥先天性免疫作用的一种重要机制。NETs由双链DNA的骨架结构和具有抗菌活性的蛋白质:髓过氧化物酶(myeloperoxidase,MPO)、组蛋白和中性粒细胞弹性蛋白酶(neutrophilelastase,NE)等组成。病原微生物(包括结核分枝杆菌)、多种促炎因子(IL-8、TNF-α等)、活化血小板、脂多糖(lipopolysaccharide,LPS)和乙酸肉豆蔻佛波酯(phorbolmyristateacetate,PMA)等多种生物化学因子均可以刺激中性粒细胞NETs形成[5]。NETs是血浆中游离DNA(cell-free DNA,cf DNA)的重要来源,因此有研究认为检测血浆中cf DNA浓度可以作为反映NETs形成的一种方式[6]。目前主要有3种方法通过检测NETs组分用于NETs的检测[7]:①检测NETs的DNA骨架结构,通过SYTOX染色的方式进行检测,这种染料可以直接插入游离的DNA中且不会对活细胞产生影响;②通过免疫荧光染色检测NETs组分髓过氧化物酶和组蛋白H3Cit,用来区分核DNA与细菌、线粒体DNA的组蛋白;③检测中性粒细胞弹性蛋白酶(NE)浓度来反映NETs的水平。本研究通过免疫荧光染色检测NETs组分MPO和H3Cit及检测cf DNA证实MTB和BCG能够在体外诱导NETs形成并建立成熟的诱导方法。
结核分枝杆菌是逃避免疫系统最成功的胞内病原体之一,主要感染呼吸系统,但也可能影响其他器官,与其他致病菌不同的是不产生毒素。由于结核分枝杆菌属于胞内感染,细胞外的中性粒细胞在结核病中可能扮演的角色被忽略。中性粒细胞可以分泌活性氧(ROS)、弹性蛋白酶、胶原蛋白酶和髓过氧化物酶,通过这些物质以一种非选择性的方式直接杀灭结核分枝杆菌和受感染的宿主细胞或者通过调节早期炎症反应增强巨噬细胞对结核杆菌的清除能力,这些对于控制结核感染至关重要[8,9]。结核分枝杆菌利用吞噬细胞复制,也作为传播至整个宿主生物体的一种方式。中性粒细胞联合巨噬细胞可以有效地抑制结核感染,并有助于早期肉芽肿形成[10]。肉芽肿是一种由免疫细胞组成的结构,用于应对原发性感染。结核分枝杆菌通过增加中性粒细胞的凋亡以阻止其产生肉芽肿来包裹这些细菌,凋亡中性粒细胞的胞葬作用导致免疫应答向促炎方向转化。不同基因型的结核分枝杆菌都可诱导中性粒细胞形成NETs。NETs 能够有效地捕获侵入的分枝杆菌,隔离有毒物质以保护周围组织免受损害,但无法杀灭捕获的结核分枝杆菌[11,12]。
大量的体内和体外试验研究证实中性粒细胞具有吞噬结核分枝杆菌的能力。在小鼠感染BCG或MTB一天后,中性粒细胞聚积在小鼠肺组织和呼吸道中,其中1.6%的中性粒细胞内含有结核分枝杆菌[13]。在体外分离的感染结核分枝杆菌的人肺组织中,大约7%的感染细胞是中性粒细胞[14,15]。而关于中性粒细胞杀死结核分枝杆菌的数据相互矛盾。有研究发现中性粒细胞的抗细菌活性较差,即使被IFN-γ刺激(能够激活巨噬细胞的抗菌性能),抗细菌活性也没有明显增强[13,16]。而另外的研究报道中性粒细胞和中性粒细胞衍生的杀菌分子确实能在体外杀死结核分枝杆菌[17,18]。中性粒细胞对MTB的杀伤能力较差,这使得一些研究者认为它们隐藏着结核分枝杆菌,从而逃避巨噬细胞的杀伤作用[19]。另一方面,中性粒细胞增加了巨噬细胞的杀菌活性:巨噬细胞吞噬凋亡的中性粒细胞,并利用其杀菌肽对抗细胞内MTB[20]。部分中性粒细胞的杀菌活性是由中性粒细胞胞外捕获网介导的,中性粒细胞经MTB刺激能释放含有中性粒细胞弹性酶和组蛋白的胞外捕获网,但不能杀死MTB[21]。此外,有人认为胞外捕获网可能为细胞外结核的生长提供了一个平台,从而有助于迅速扩大肺部病变[22]。另外中性粒细胞胞外捕获网可通过降低T细胞的活化阈值直接激活T细胞[23]。热休克蛋白72(HSP-72)对消除结核分枝杆菌具有重要作用,HSP-72存在于凋亡细胞和坏死细胞中,同时在NETs内的DNA也被发现,这些对于遏制和消除结核分枝杆菌具有重要意义[24]。
综上所述,关于中性粒细胞是否能够杀死MTB存在很大争议。这些具有极高杀菌潜力的中性粒细胞及NETs对分枝杆菌是否有明确的杀菌作用,还有待进一步研究探讨。
猜你喜欢
安徽建筑大学学报(2022年4期)2022-09-27
实用手外科杂志(2022年2期)2022-08-31
皮肤病与性病(2021年3期)2021-07-30
昆明医科大学学报(2021年4期)2021-07-23
生物化工(2020年5期)2020-10-30
科学导报·学术(2020年29期)2020-10-21
农业环境科学学报(2020年9期)2020-10-09
美与时代·美术学刊(2019年9期)2019-11-29
家庭影院技术(2018年9期)2018-11-02
女友·家园(2017年7期)2017-07-19
