
类表皮生长因子域7在创伤性脑损伤后脑组织中的表达变化及意义
2020-02-07 13:30徐鹏翔许琼冠李强罗孟亚男成凯谢镇明付宙锋
中国老年学杂志 2020年2期
徐鹏翔 许琼冠 李强 罗孟亚男 成凯 谢镇明 付宙锋
(海南医学院第二附属医院 1神经外科,海南 570311;2放射治疗科)
创伤性脑损伤(TBI)已成为全球死亡率和发病率的主要原因〔1〕。脑外伤幸存者经常遭受认知、生理和心理社会功能的暂时或永久性损害。血管生成不仅存在于身体的生长发育过程中,而且是伤口愈合、缺血、缺氧和炎症过程中的一种生理现象,与多种疾病的发生、发展和预后密切相关〔2〕。表皮生长因子样结构域(EGFL)7是近年来发现的内源性血管生成调节因子,在胚胎血管生成及大多数正常成年组织在生理或病理性血管生成过程中起关键作用〔3,4〕。EGFL7编码蛋白含有分泌信号肽序列,该序列具有半胱氨酸(EMI)结构的N-末端以调节细胞间黏附,两个与蛋白质鉴定相关的EGF-样结构及富含赖氨酸和缬氨酸的高保守C-末端〔5〕。体外研究显示,神经干细胞(NSC)可通过EGFL7参与调控血管的形成,然而EGFL7在TBI中的表达和功能尚不清楚〔6〕。本研究拟通过建立TBI大鼠模型检测脑组织中EGFL7的表达情况及EGFL7在脑血管生成中的作用机制。
1 材料和方法
1.1实验动物 成年雄性SD大鼠,6周龄,体重291~342 g,23℃的室温下以12 h的暗-光循环,自由获取食物和水。随机分为假手术组(sham,n=10)和TBI组(n=60),TBI组分为TBI后1 h、6 h、24 h、3 d、7 d、14 d 6个亚组各10只。TBI组进行实验性脑外伤造模,并在受伤后的上述6个时间点处死。
1.2大鼠TBI模型建立 参照文献方法建立了弥漫性头部损伤的大鼠模型〔7〕。每只大鼠腹腔注射4%水合氯醛(50 mg/kg体重)麻醉,同时使用加热垫维持正常体温。随后将头部固定在立体定位框架中,沿中线制作10 mm切口以暴露颅骨。在保持硬脑膜完整的同时,在冠状缝线后3.0 mm和矢状缝侧面2.0 mm处进行右开颅(直径4 mm)。应用1.8~2.0 atm的冲击压力制造TBI模型。然后缝线封闭切口,让大鼠从麻醉中恢复。假手术组仅进行开颅手术。所有手术均在无菌环境中操作。
1.3收集脑组织标本 分别在TBI后不同时间后处死并采集大脑组织标本。在4%水合氯醛(50 mg/kg体重,腹腔注射)深麻醉下对每组6只进行开胸手术。用0.9%的生理盐水500 ml加肝素钠12 500 U通过左心室内灌注,打开右心房,直到流出液清澈为止,采集挫伤区周围皮层样本,立即置于液氮中,进行Western印迹和实时荧光PCR分析。采用0.9%生理盐水灌注脑组织进行免疫组化分析,浸泡于4℃的4%多聚甲醛缓冲液24 h。
1.4Western印迹分析 将储存在液氮中的大脑挫伤区周围组织样本解冻,溶解在放射免疫沉淀法(RIPA)缓冲液〔1%乙基苯基聚乙二醇(NP40),0.5%脱氧胆酸钠,0.1%十二烷基硫酸钠(SDS),1 mmol/L钒酸钠(Na3VO4),1 mmol/L乙二醇双(2-氨基乙基醚)四乙酸(EGTA),0.5 mmol/L二硫苏糖酸(DTT),20 mmol/L氟化钠(NaF),1 mmol/L苯甲基磺酰氟(PMSF)〕中,并补充蛋白酶抑制剂。匀浆在10 000 r/min下4℃离心15 min,然后收集上清液,并测定其中蛋白质浓度。在12% SDS-聚丙烯酰胺凝胶上加载等量(每道 40 μg)蛋白质,在70 V的恒定电压下电泳30 min,然后在120 V下电泳90 min,最后在300 mA下转移到聚偏氟乙烯膜。用5%脱脂乳在20℃封闭膜2 h,然后加入兔抗EGFL7抗体(5%脱脂乳中稀释1∶1 000;美国BioWorld技术公司)、兔抗CD34抗体(5%脱脂乳中稀释1∶500;美国BioWorld技术公司)和β-actin(5%脱脂乳中稀释1∶4 000;美国BioWorld公司)孵育过夜。在三乙醇胺缓冲盐水溶液(TBS)中清洗3次,并在室温下用适当的辣根过氧化物酶(HRP)结合二级抗体(在吐温20中稀释1∶5 000;美国Santa Cruz公司)覆盖2 h。膜再次清洗3次,并在增强化学发光检测系统(英国Amersham Biosciences公司)中孵育,之后暴露于射线照相胶片(日本Fujihyperfilm公司)。根据平均像素密度计算为靶蛋白/β-actin表达的比率。
1.5RNA分离与实时荧光定量PCR分析 使用Trizol试剂(中国Takara生物技术公司)从冷冻的皮层组织中提取总RNA。用分光光度法(OD260/280范围1.8~2.0)和1%琼脂糖凝胶电泳测定RNA的浓度和纯度。利用PrimescriptTMRT试剂盒(中国Takara生物技术公司)和寡核苷酸引物将分离出的RNA反转录成cDNA。引物由上海生工生物工程股份有限公司合成,所用序列来自大鼠EGFL7和GAPDH的NCBI数据库。大鼠EGFL7正向和反向引物分别为5′-CCGGCCATTTGATGCCT-3′、5′-GCGAGTGCTTTGAGTAGAG-3′;大鼠GAPDH正向和反向引物分别为5′-AATGTCCGTCGTGGATCTGA-3′、5′-AGTGTAGCCCAAGATGCCTT C-3′。采用实时SYBR Green qPCR Master Mix(中国Takara生物技术公司)进行定量实时PCR分析。反应混合物中含有10 μl SYBR预混物Ex Taq、0.8 μl引物(10 μmol/L)、2 μl cDNA和7.2 μl的核酸酶游离水,最终体积为20 μl。在95℃循环30 s后进行40个PCR循环,每个循环包括变性步骤(95℃,5 s)和退火步骤(60℃,30 s)。用GAPDH mRNA的数量对每个样品的总RNA浓度进行归一化,EGFL7基因的表达水平由EGFL7 mRNA与GAPDH mRNA的比率来评估。
1.6免疫组化分析 采用4%多聚甲醛在4℃固定脑组织,石蜡包埋。切割4 μm厚的脑冠状部(包括挫伤区和海马区)进行免疫组化。将切片在磷酸盐缓冲液(PBS)中用3%的H2O2脱蜡培养10 min,然后用5%的正常胎牛血清在PBS中阻断2 h。切片在4℃下用兔抗EGFL7抗体(稀释1∶200;美国BioWorld技术公司)和兔抗CD34抗体(稀释1∶500;美国BioWorld技术公司)孵育过夜,然后在PBS中洗涤30 min,用HRP标记的山羊抗兔免疫球蛋白(Ig)G(稀释1∶500;美国Santa Cruz公司)室温下孵育1 h。再次冲洗30 min后,用二氨基联苯胺进行免疫标记,苏木精复染。每个脑组织样本中随机选取5个区域用于细胞计数。在光学显微镜下测定EGFL7、CD34阳性细胞的数量,并由一名不了解该分组的研究者进行分析。采用6个随机非重叠区的EGFL7、CD34阳性细胞平均数进行统计分析。
1.7细胞转染 采用人EFGL7基因序列5′-GCACCTACCGAACCATCTATA-3′合成siRNA引物:5′-GATCCGTTCTCCGAACGTGTCACGTTTCAAGAGAAC GTGACACGTTCGGAGAACTTTTTTG-3′和5′-AATTCAAAAAAGTTCTCCGAACGTGTCACGTTCTCTTGA AACGTGACACGTTCGGAGAACG-3′。退火后,将引物与线性穿梭质粒病毒载体(pLV3)连接,形成pLV3/siRNA/EGFL7复合物,用于转染课题组前期建立的NSC和内皮细胞(HUVEC)共培养系统〔6〕,具体方法为采用0.4 μm孔径的Transwell小室将NSCs和HUVECs以1∶1比例〔(1×104)∶(1×104)〕分隔培养,Transwell小孔放置NSCs,24孔平板中接种HUVECs。共培养24 h后将含8 μg/ml聚布伦的DMEM全培养基稀释EGFL7-siRNA转染载体加到含有NSC-HUVEC共培养系统中融合,继续培养72 h后,收集细胞并按照1.4中方法分析EGFL7、CD34、磷脂酰肌醇-3-激酶(PI3K)、蛋白激酶B(AKT)、磷酸化(p)-AKT蛋白表达。对照组仅加入DMEM全培养基。
1.8统计学分析 采用SPSS18.0软件进行单因素方差分析、Post Hoc检验。
2 结 果
2.1TBI诱导EGFL7和CD34表达上调 EGFL7 mRNA、蛋白及CD34蛋白水平在脑外伤后24 h显著升高(P<0.05,P<0.01),第3天达峰值,并持续至少14 d。见表1。
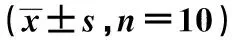
表1 脑外伤后不同时间点挫伤周围皮层EGFL7 mRNA的表达水平和EGFL7和CD34的蛋白表达
与sham组比较:1)P<0.05,2)P<0.01
2.2挫伤周围皮层和同侧海马区EGFL7和CD34阳性细胞数 与假手术组相比,TBI大鼠挫伤周围皮层和海马区EGFL7和CD34阳性细胞显著增加(P<0.01),见图1、2,表2。
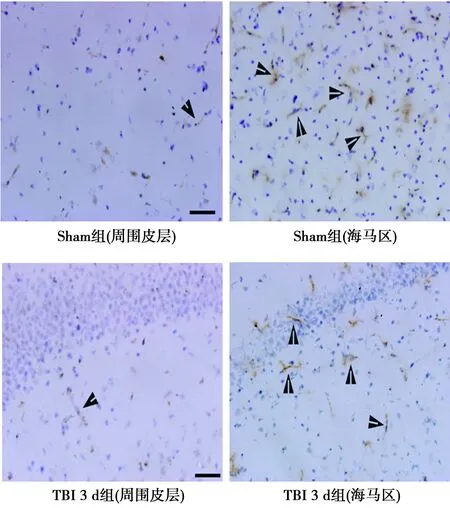
图1 TBI大鼠挫伤周围皮层和海马区EGFL7阳性细胞(免疫组化,×100)
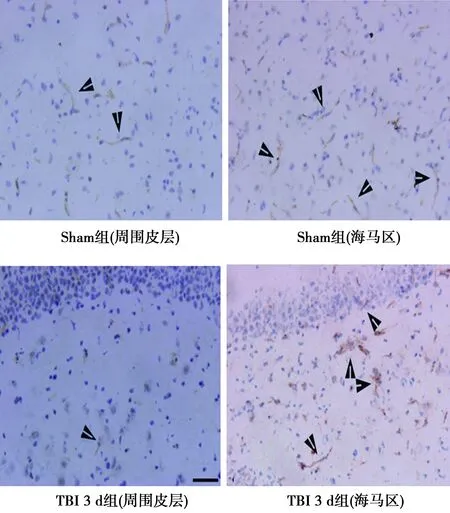
图2 TBI大鼠挫伤周围皮层和海马区CD34阳性细胞(免疫组化,×100)
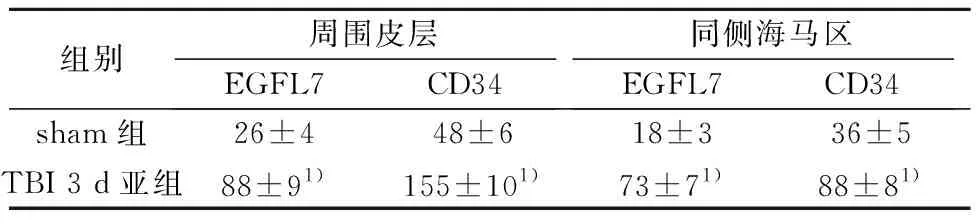
组别周围皮层EGFL7CD34同侧海马区EGFL7CD34sham组26±448±618±336±5TBI 3 d亚组88±91)155±101)73±71)88±81)
与sham组比较:1)P<0.01
2.3沉默EGFL7对NSC-HUVEC共培养系统的p-AKT、AKT、PI3K表达影响 特异性siRNA干扰致EGFL7的蛋白表达明显受到抑制后,EGFL7-siRNA表达载体转染组NSC-HUVEC共培养系统的CD34、AKT、p-AKT、PI3K蛋白的相对表达均较对照组显著降低(P<0.05,P<0.01),见表3、图3。
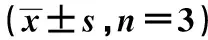
表3 NSC-HUVEC共培养系统经EGFL7-siRNA转染72 h后EGFL-7、CD34、p-AKT、AKT、PI3K蛋白表达
与对照组比较:1)P<0.05,2)P<0.01
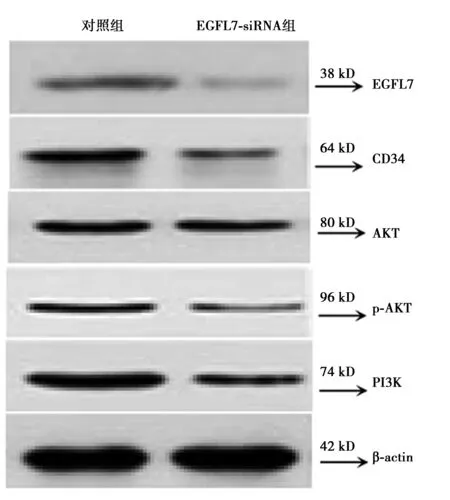
图3 NSC-HUVEC共培养系统经EGFL7-siRNA转染72 h后EGFL-7、CD34、p-AKT、AKT、PI3K蛋白的表达
3 讨 论
血管生成能为组织提供氧气和营养,改善组织缺血,促进受损脑组织的结构重塑,从而加速神经功能的修复〔8〕。因此,促进血管生成可能是治疗脑外伤后缺血性脑损伤的一种新方法。血管生成是一个复杂的过程,对神经细胞的修复和大脑损伤后的功能恢复至关重要〔9〕。EGFL7是一种新的促血管生成因子,在胚胎发育过程中对血管生成起关键调节作用。在正常成人组织中,EGFL7表达下调或不表达,但在富含血管的组织(包括肺、心脏和子宫)中处于高水平表达〔10〕。EGFL7在生理或病理条件下重新激活,参与血管生成〔6〕。此外,EGFL7的高表达与几种肿瘤类型中的血管形成有关,包括结肠、胃、乳腺、肾、肝和脑肿瘤〔11~13〕。本研究表明EGFL7表达可能是促进脑损伤后血管生成的关键因素。尽管以前的研究表明EGFL7是由血管HUVEC或其前体细胞分泌〔12〕,但本研究免疫组织化学染色显示,在TBI大鼠挫伤周围皮层和海马区中,EGFL7均有表达。EGFL7在神经细胞中的表达也被证实。这可能意味着在神经细胞中存在先前未识别的EGFL7功能,即在HUVEC上或两者上存在旁分泌血管生成功能。总的来说,这些发现似乎是EGFL7应对脑损伤后血管生成的关键因素,了解TBI大鼠挫伤进展过程中EGFL7的表达是如何启动的及控制EGFL7的位置和表达的因素将有助于理解脑外伤修复的细胞和分子机制。研究证实,在EGFL7高表达情况下,HUVEC可形成较多的管道样结构,而将EGFL7基因沉默后,上述管道样结构消失,说明NSCs可通过EGFL7参与调控血管的形成〔6,14〕。基于这些发现,本研究显示EGFL-7基因沉默显著抑制了NSC-HUVEC共培养系统中p-AKT、PI3K蛋白表达及HUVEC的血管生成能力。PI3K/AKT信号通路是执行多种生物学功能的信号调控系统,如细胞存活、增殖,代谢调控和细胞迁移等〔15,16〕。PI3K活化后在细胞膜上生成PIP3,并与信号蛋白分子AKT结合并活化AKT〔17〕。Karar等〔18〕研究显示AKT的持续内皮激活诱导血管的发育和血管管腔结构的形成。因此,上述结果提示EGFL-7可能通过PI3K/AKT通路参与NSCs的血管生成。
猜你喜欢
农业工程学报(2022年5期)2022-06-22
中国典型病例大全(2022年10期)2022-05-10
健康体检与管理(2022年2期)2022-04-15
世界科学技术-中医药现代化(2021年8期)2021-12-21
医学前沿(2021年18期)2021-04-14
中国医学影像技术(2020年11期)2021-01-04
中华养生保健(2020年5期)2020-11-16
中国现代医药杂志(2020年3期)2020-05-08
中华耳科学杂志(2020年3期)2020-01-09
中国畜牧杂志(2019年10期)2019-01-12
