
紫外辐射对小球藻光合性能及油脂积累的影响
2020-01-16 02:12:44孙小琴李鹏飞刘兴社成智文
中国油脂 2019年12期
孙小琴,孙 昕,李鹏飞,刘兴社,成智文,李 青
(1.西安建筑科技大学 陕西省环境工程重点实验室,教育部西北水资源和环境生态重点实验室,西安 710055; 2.咸阳陶瓷研究设计院有限公司,陕西 咸阳 712000)
随着化石能源的日益枯竭和人们对环境保护的不断重视,生物柴油作为新型的清洁能源,受到广泛关注[1-2]。小球藻油脂含量较高,生长速度快,是一种理想的能源微藻[3-4]。高昂的生产成本是微藻产业化生产的一大阻碍,而提高油脂产量是有效的解决途径之一[5]。有研究表明,胁迫生长可诱导微藻以中性脂质或多糖的形式积累能量[6],而紫外线胁迫被视为是一种良好的处理方式,其操作简单、突变率高,尤其在高脂微藻的培育中具有污染少、辐射剂量改变灵活、诱导快速等优点[7-8]。叶丽等[9]用紫外线辐射三角褐指藻,得到总脂含量高达36.66%的突变藻株;Liu等[10]利用紫外线辐射小球藻,突变藻株的油脂含量增加了21.97%。然而在紫外诱变育种中,优良性状藻株的筛选过程耗时长,筛选后的藻株存在不稳定性,这对能源微藻的产业化生产造成一定的阻碍,如能省去复杂的筛选程序,利用紫外线直接对藻液进行辐射,既可提高微藻的油脂产量,又能缩短培育时间,将明显发挥紫外诱变的优势,但目前大多研究仍停留在利用诱变筛选突变株这一层次上。本文创新利用紫外诱变的优势,避免了长时间的筛选过程,以期突破微藻产业化的瓶颈。
本文采用紫外线直接对小球藻藻液进行辐射,通过测定不同紫外辐射剂量下小球藻的生物量、叶绿素荧光参数、油脂含量以及脂肪酸组成及含量,确定最佳紫外辐射剂量,并利用荧光定量PCR检测与油脂合成相关基因的表达情况,分析紫外辐射促进微藻产油的可能生化途径,以期为高产油微藻的培育提供理论基础,为微藻的产业化生产提供应用依据。
1 材料与方法
1.1 实验材料
小球藻,购于中国科学院武汉水生生物研究所淡水藻种库。BG11培养基,主要成分如下:(NH4)3C6H5O7,0.006 g/L;C6H7O8,0.006 g/L;EDTA-MgNa2,0.001 g/L;NaNO3,1.5 g/L;K2HPO4,0.039 g/L;MgSO4·7H2O,0.075 g/L;CaCl2·2H2O,0.036 g/L;Na2CO3,0.02 g/L;H3BO4,2.86 mg/L;MnCl2·4H2O,1.81 mg/L;ZnSO4·7H2O,0.222 mg/L;NaMoO4·2H2O,0.391 mg/L;CuSO4·5H2O,0.079 mg/L;Co(NO3)·6H2O,0.049 mg/L。
光照培养箱,SW-CJ-1D净化工作台,UVC-254型紫外辐照度计,UV2600型紫外可见分光光度计,真空冷冻干燥机,M系列IMAGING-PAM调制叶绿素荧光成像系统,7890A/7000B型气相色谱质谱联用仪,CFX96TM实时定量PCR仪。
1.2 实验方法
1.2.1 小球藻的培养
采用BG11培养基接种小球藻获得小球藻藻液。
将装有小球藻藻液的500 mL三角锥形瓶,用无菌膜封闭瓶口,置于恒温光照培养箱中,在平均光照强度70.7 μmol/(m·s)、光照周期12L/12D、培养温度(25±1)℃条件下,采用半连续培养模式培养。每天随机调换锥形瓶的位置并摇动3~4次,定时取样进行分析。
1.2.2 紫外辐射诱变
实验用紫外灯管功率20 W,发射波长254 nm,实验前将紫外灯打开照射30 min以稳定光强并杀菌。取200 mL小球藻藻液于18 cm玻璃培养皿中,置于磁力搅拌器上,保证紫外线能均匀辐射藻液。通过改变紫外辐射时间设置5个辐射剂量梯度(0、20、40、60、80 mJ/cm2),每个梯度设2个平行样。藻液经紫外辐射后用锡纸包裹培养皿置于恒温光照培养箱中避光培养24 h避免“光复活”[11],再转移至500 mL三角锥形瓶中按1.2.1条件继续进行培养。
本实验中藻液与紫外灯的垂直距离为30 cm,通过紫外辐照度计测量培养皿圆心处和圆周4个切点处藻液表面处的紫外辐射强度,得到平均辐射强度(I)为0.27 mW/cm2,根据下式[12]计算辐射时间。
D=I×t
式中:D为辐射剂量,mJ/cm2;t为辐射时间,s。
1.2.3 生物量的测定
光密度法:取一定量的藻液于1 cm石英比色皿中,采用紫外可见分光光度计测定680 nm处的吸光度(OD)。
称重法:将藻液在10 000 r/min、4℃下离心10 min,并用蒸馏水洗涤2~3次,除去原培养基中的代谢产物等杂质,再用冷冻干燥机获得干燥藻粉,称取藻粉质量。
1.2.4 叶绿素荧光参数的测定
采用调制叶绿素荧光成像系统测定小球藻的叶绿素荧光参数Fv/Fm、Y(Ⅱ),检测前将藻液暗适应5 min。
1.2.5 油脂含量的测定
采用改进的Bligh-Dyer法[13]测定油脂含量。取10 mg冻干的藻粉,加入1.6 mL蒸馏水、4 mL甲醇、2 mL氯仿,用涡旋振荡器将混合液混匀振荡2 min,超声破碎5 min,再加入2 mL氯仿继续涡旋振荡1 min,加入2 mL蒸馏水涡旋振荡1 min,使最终的氯仿、甲醇、蒸馏水体积比为1∶1∶0.9,再次离心,静置分层,取下层氯仿相,再向原管中加入氯仿继续萃取1次,合并氯仿相并在氮气下吹脱,得到小球藻油,称重计算小球藻油脂含量。
1.2.6 荧光定量PCR的测定
本次测定的目的基因是accD、gapA、Me、Pk。gapA是GADPH(甘油醛-3-磷酸脱氢酶)的编码基因,GADPH是Calvin循环的重要参与酶,与碳代谢有密切联系。accD是ACCase(乙酰辅酶A羧化酶)的编码基因,在自养条件下,其转录量与油脂积累量呈正相关[11]。Me是ME(苹果酸酶)的编码基因,ME可将苹果酸催化为丙酮酸盐,并产生NADPH(还原型辅酶Ⅱ),NADPH可以作为还原力和能量,对脂肪酸的生物合成过程是必要的。Pk是PK(丙酮酸激酶)的编码基因,PK活性降低时,糖酵解途径受阻,对油脂的积累产生一定的影响[13]。
具体测定按如下流程进行操作:①用Trizol法提取总RNA。②反转录。反转录体系见表1,添加试剂成分见表2。③引物设计。设计内参基因18SRNA和目的基因的引物[14],如表3所示。④荧光定量PCR。按表4配制qPCR反应体系,将样品放入CFX96TM实时定量PCR仪中检测,设置检测参数为95℃预变性5 min,95℃变性10 s,60℃退火30 s,循环54次。
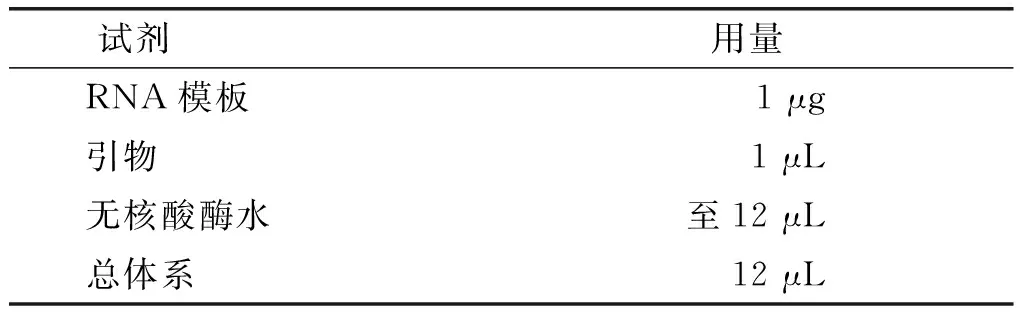
表1 反转录体系

表2 添加试剂成分

表3 内参基因和目的基因的引物序列

表4 qPCR反应体系
1.2.7 小球藻油脂肪酸组成分析
将抽提的小球藻油用NaOH-CH3OH溶液皂化,加入30%的二氧化碳乙醚络合物甲醇溶液进行酯化,再加入2 mL正己烷振荡5 min,静置后取上层的正己烷液上机分析[15]。用气相色谱质谱联用仪测定脂肪酸的组成和含量。GC条件:DB-23石英毛细管色谱柱(60 m×0.25 mm×250 μm);进样量1 μL;分流比50∶1;进样口温度210℃;氦气流速1 mL/min;升温程序为80℃保持5 min,以4℃/min的速度升到290℃,并在290℃条件下保持5 min。MS条件:电子能量70 eV;四极杆温度150℃;离子源温度250℃;溶剂延迟2 min。质谱图利用NIST标准谱库进行检索,采用面积归一化法定量。
2 结果与分析
2.1 不同紫外辐射剂量下的小球藻藻液的吸光度
图1为不同紫外辐射剂量下小球藻藻液吸光度随培养时间的变化情况。
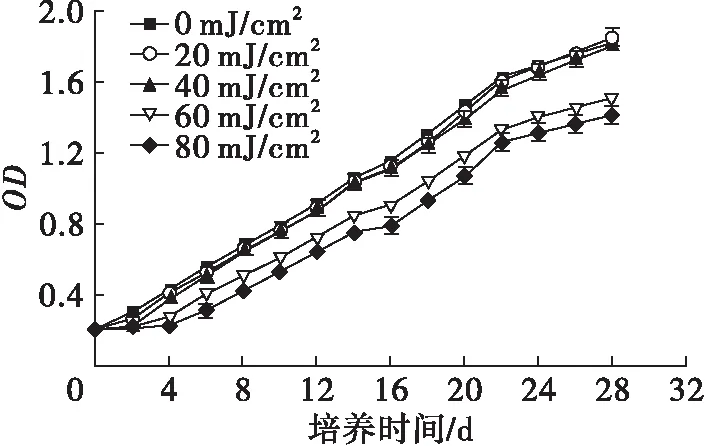
图1 不同紫外辐射剂量下小球藻藻液吸光度随培养时间的变化
由图1可见,经过紫外辐射后,小球藻的生长受到了不同程度的抑制,辐射剂量越大,抑制作用越强。20、40 mJ/cm2剂量对小球藻的抑制作用较弱,2 d后小球藻即可恢复生长,生长速度与对照组(紫外辐射剂量为0 mJ/cm2,下同)基本一致。60、80 mJ/cm2剂量对小球藻的抑制作用较强,需经4 d恢复生长,生长速度慢于对照组。从22 d开始,小球藻的生长速度变慢,进入稳定期,剂量为20、40 mJ/cm2的小球藻吸光度与对照组的基本相等,剂量为60、80 mJ/cm2的小球藻吸光度明显低于对照组的。Liu等[10]用紫外诱变小球藻,突变藻株的生物量比原始藻株提高了7.6%。说明在适宜的紫外辐射条件下,微藻的生长并不会受到抑制。
2.2 不同紫外辐射剂量下小球藻的叶绿素荧光参数
2.2.1Fv/Fm
Fv/Fm反映植物潜在的最大光合作用能力。不同紫外辐射剂量下小球藻的Fv/Fm随培养时间的变化情况见图2。

图2 不同紫外辐射剂量下小球藻Fv/Fm随培养时间的变化
由图2可见,紫外辐射剂量越大,小球藻的Fv/Fm值越低,说明紫外辐射使小球藻的光合作用能力降低,降低程度随紫外辐射剂量增大而增强。不同紫外辐射剂量下的Fv/Fm在一定培养时间内会恢复到相对稳定值,剂量越大,恢复时间越长,最终的稳定值越低。这可能是小球藻的光系统启动了各种热耗散形式的光保护途径,受损的光系统逐渐修复,但修复能力随紫外辐射剂量的增加而削弱。20、40 mJ/cm2剂量下的最高Fv/Fm值分别为0.593、0.588,与对照组的相当,表明其光合性能良好。而60、80 mJ/cm2剂量下的Fv/Fm值较对照组的低,说明其光合性能变差。
2.2.2Y(Ⅱ)
Y(Ⅱ)表征光系统的实际光能捕获效率,是植物光合能力的重要衡量指标[16]。不同紫外辐射剂量下小球藻的Y(Ⅱ)随培养时间的变化情况见图3。

图3 不同紫外辐射剂量下小球藻Y(Ⅱ)随培养时间的变化
由图3可见,在3~5 d,不同紫外辐射剂量下小球藻的Y(Ⅱ)不同程度地降低,剂量越大,Y(Ⅱ)值越低,说明光系统PSⅡ因紫外辐射而受到损伤,光能转化效率降低,且PSⅡ的损伤程度随紫外辐射剂量的增加而增大。PSⅡ的损伤可能表现在两方面:一是光合器官受到损伤,光系统中的捕光色素蛋白复合体自身解体甚至失去捕光功能[17];二是紫外线导致PSⅡ核心复合体的反应中心D1蛋白甚至D2蛋白受损,为避免这种损伤,藻类会相应地提高D1蛋白的周转速度以促进自我修复[18]。因此,在5~11 d,Y(Ⅱ)逐渐升高,不同剂量间的差异缩小。在11~23 d,Y(Ⅱ)基本达到稳定,23 d开始,小球藻进入生长后期,光化学反应不如对数生长期活跃,Y(Ⅱ)逐渐降低。与Fv/Fm 的变化特性类似,剂量低于40 mJ/cm2下的Y(Ⅱ)最终稳定在较高值,20、40 mJ/cm2剂量下的Y(Ⅱ)最高值分别为0.598、0.594,与对照组的相当,表明光系统的光能捕获效率高;剂量高于40 mJ/cm2下的Y(Ⅱ)则相反。
2.3 不同紫外辐射剂量下小球藻的干重与产油情况
对不同紫外辐射剂量下的小球藻培养28 d,测定其干重和产油情况,结果见图4、图5。

图4 不同紫外辐射剂量下小球藻的干重与油脂含量
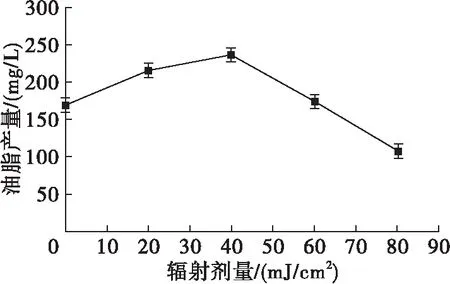
图5 不同紫外辐射剂量下小球藻的油脂产量
由图4可见,20、40 mJ/cm2剂量下小球藻的干重分别为530、515 mg/L,与对照组相差不大,其他剂量下的干重则小于对照组。20、40 mJ/cm2剂量下的油脂含量均有提升,其中40 mJ/cm2剂量下的油脂含量最高,为46%,比对照组提高了39.40%,之后随着紫外辐射剂量的增大,小球藻的油脂含量下降。由图5可见,与对照组相比,20~40 mJ/cm2剂量下的油脂产量均有提高,其中40 mJ/cm2剂量下油脂产量最高,达237 mg/L,比对照组提高了39.22%,之后随着紫外辐射剂量的增大,小球藻的油脂产量下降。Sharma等[19]利用适宜的辐射剂量对小球藻进行紫外诱变,得到突变藻株的总脂含量几乎加倍。这些研究结果说明适宜的紫外辐射可在一定程度上提高微藻的产油能力,可能是紫外线辐射导致小球藻内与油脂代谢相关的基因或其调控序列发生突变,基因产物的活性发生变化,从而引起油脂含量的变化[20]。
2.4 不同紫外辐射剂量下小球藻的荧光定量PCR结果
图6显示了不同紫外辐射剂量下小球藻目的基因的相对表达情况(以对照组的基因表达量为标准)。

图6 不同紫外辐射剂量下小球藻目的基因的相对表达量
由图6可见:各辐射剂量下accD、gapA的相对表达量均提高,其中40 mJ/cm2剂量的提升幅度最大,分别提高了约2.5倍和5倍;除了80 mJ/cm2剂量下Me的相对表达量降低,40 mJ/cm2剂量下Me的相对表达量基本变化不大外,其他剂量下的Me相对表达量均提高,其中60 mJ/cm2剂量的提升幅度最大,提高了约1倍;除40 mJ/cm2剂量下Pk的相对表达量未降低外,其他剂量下Pk的相对表达量均有较大幅度的降低。理论而言,小球藻的油脂积累可能与ACCase有关,ACCase所催化的反应是脂肪酸合成的第一步,在脂肪酸的合成与代谢中发挥重要作用[21],accD表达量的提高会促进藻体内脂肪酸的合成。Calvin循环为自养藻类提供合成油脂的碳源,相关酶的作用可能与油脂积累密切相关,GAPDH是Calvin循环的作用酶之一,gapA表达量的提高会促进更多碳源流向油脂合成。与Calvin循环密切相关的戊酮磷酸途径,可产生大量NADPH,为油脂合成提供能量,所以Me表达量的提高可能使小球藻的油脂合成具备更多的能量,有研究表明Me与脂质积累量显著相关,其表达情况会影响脂质的积累[22]。PK是将磷酸烯醇式丙酮酸转化为丙酮酸的酶,丙酮酸的一部分进行发酵产生NAD+,一部分转化为乙酰辅酶A,这两种物质均与油脂合成密切相关,因此Pk表达量的提高可能会促进油脂合成。在40 mJ/cm2剂量下,accD和gapA的相对表达量呈较大幅度提高,Me和Pk的相对表达量基本不变,说明accD和gapA表达量的提高是促进油脂积累的主要驱动力。虽然80 mJ/cm2剂量下accD和gapA的表达量有所提升,但Me和Pk的表达量均下降,说明Me和Pk表达量的降低可能对油脂的积累产生一定的抑制。
2.5 不同紫外辐射剂量下小球藻油的脂肪酸组成和含量(见表5)

表5 不同紫外辐射剂量下小球藻油的脂肪酸组成和含量
由表5可见,共检测出10种主要脂肪酸,包括5种饱和脂肪酸(SFA)、2种单不饱和脂肪酸(MUFA)、3种多不饱和脂肪酸(PUFA)。小球藻油主要的脂肪酸为棕榈酸(C16∶0)、棕榈油酸(C16∶1)、油酸(C18∶1)、亚油酸(C18∶2)和亚麻酸(C18∶3)。其中,C16∶1和C18∶1变化较大,与对照组相比,20~60 mJ/cm2剂量下的C16∶1和C18∶1含量明显增加,40 mJ/cm2剂量下的提升幅度最大,MUFA含量达到40.17%,比对照组提高了16.5%。而MUFA更有利于生物柴油的生产,表明该剂量条件适合培育制备生物柴油的小球藻,在实际应用方面具有较高的价值。
3 结 论
综合比较小球藻液紫外辐射后小球藻的生长、光合性能、油脂积累以及脂肪酸含量,确定40 mJ/cm2为最佳紫外辐射剂量。在40 mJ/cm2剂量下,小球藻的生长情况及光合性能较好,生物量与对照组(未进行紫外辐射)基本相等,叶绿素荧光参数降低幅度较小,且在很短的时间内即可恢复,最终的Fv/Fm,Y(Ⅱ)与对照组的荧光参数值接近。另外,辐射剂量为40 mJ/cm2的小球藻产油性能最佳,相对于对照组,油脂含量和油脂产量分别提高了39.40%和39.22%;与油脂合成相关的accD和gapA基因的相对表达量分别提高了约2.5倍和5倍,Me和Pk基因的相对表达量基本不变。而且,紫外辐射剂量对小球藻油脂肪酸组成及含量有一定影响,辐射剂量为40 mJ/cm2的小球藻油脂肪酸组成中的MUFA含量有较大幅度的提升,含量达到40.17%,比对照组提高了16.5%。
猜你喜欢
古今农业(2022年1期)2022-05-05 06:58:42
天津造纸(2021年2期)2021-11-29 11:50:08
科学养鱼(2020年9期)2020-10-16 07:08:24
陶瓷学报(2019年5期)2019-01-12 09:17:44
生物加工过程(2018年4期)2018-07-24 02:13:22
环境保护与循环经济(2017年3期)2017-09-26 11:42:30
电源技术(2016年2期)2016-02-27 09:04:55
化工进展(2014年10期)2014-07-05 16:06:08
食品工业科技(2014年23期)2014-03-11 18:19:31
华东理工大学学报(自然科学版)(2014年1期)2014-02-27 13:48:31
