
猴源DDX3X基因的克隆及生物信息学分析
2020-01-14 08:59张升波张喜懿温贵兰程振涛王开功周碧君
贵州农业科学 2019年12期
杨 洪,田 浪,张升波,张喜懿,温贵兰*,程振涛,王开功,周碧君
(1.贵州大学 动物科学学院 预防兽医实验室,贵州 贵阳 550025;2.贵州省动物生物制品工程技术研究中心,贵州 贵阳 550016)
DEAD-box家族蛋白是一类ATP依赖的RNA解旋酶,在RNA代谢的过程中发挥重要作用[1],可通过调节和抑制癌基因的表达及肿瘤相关信号通路等影响细胞的增殖、分化与凋亡,进而发挥促癌或抑癌的作用[2]。有研究显示,DDX3X基因特异突变可导致发育障碍和智力残疾或发育迟缓等现象[3],并且可能影响女性和男性之间疾病传播和表型变异,DDX3X对正常发育和疾病具有剂量依赖性和性别特异性影响。有研究报道,在胚胎发育过程中,靶向敲除小鼠DDX3X基因后,导致外胚层广泛凋亡和异常生长,最终胚胎发生消融并且死亡[4]。DEAD-box蛋白家族是RNA解旋酶中最大的亚家族,其具有9个保守结构域,因motifyⅡ的保守氨基酸序列Asp-Glu-Ala-Asp (DEAD)而命名[4]。DEAD-box解螺旋酶蛋白家族在各种生物体内普遍存在并且结构上高度保守,主要包括DEAD-box RNA解螺旋酶和DEAD-box DNA解螺旋酶2大类[5],而DDX3X是通过利用ATP产生的能量结合或者重塑RNA以及核糖核蛋白复合体的RNA解螺旋酶,其作为DEAD-box RNA解螺旋酶蛋白家族中重要成员而被广泛研究[6]。DDX3X参与许多病毒的复制过程,如猪繁殖与呼吸综合征病毒(PRRSV)、丙型肝炎病毒(HCV)和人类免疫缺陷病毒(HIV)等[7-8]。
猴源肾细胞源于恒河猴肾上皮样细胞,其对猪繁殖与呼吸综合征病毒(PRRSV)易感[9]。基于DDX3X基因的关键生物学功能,研究拟克隆猴源DDX3X基因,并分析其序列特征及蛋白质性质和功能结构域,旨在为后续深入研究猴源DDX3X基因与病毒间的相互作用及分子机制奠定基础。
1 材料与方法
1.1 材料
1.1.1 试剂 RNA提取试剂盒、质粒提取试剂盒、反转录试剂盒和2×Es Tag Master Mix 购自康为世纪生物科技有限公司,2 000 bp DNA marker、限制性核酸内切酶EcoR I、 Xho I和pMD19-T载体购自宝生物工程有限公司,胶回收试剂盒购自上海生工生物工程技术服务有限公司。
1.1.2 仪器 供试仪器为PCR仪(ABI公司)、倒置荧光显微镜(OLYMPUS公司)和凝胶成像系统(Gene公司)。
1.1.3 实验细胞 猴源肾细胞和DH5α感受态细胞由贵州大学动物科学学院预防兽医实验室保存并提供。
1.2 方法
1.2.1 引物设计与合成 根据NCBI公布的猪源DDX3X基因序列(登录号HQ266638)设计特异性引物,上游引入EcoR I、下游引入Xho I作为酶切位点。上游引物(DDX3X-EcoR I-F)为GAATTCATGAGTCATGTGGCGGTGG,下游引物(DDX3X-Xho I-R )为CCCTCGAGAGTTGCCCCACCAGTCAAC,送生工生物工程(上海)股份有限公司合成。
1.2.2 猴源肾细胞培养、总RNA提取及cDNA合成 将复苏的猴源肾细胞于37℃细胞培养箱培养24 h,于倒置荧光显微镜下观察细胞生长状态,待其汇合度为90%~100%时,用预冷无菌PBS清洗3次,加入1 mL Trizol于冰上裂解10 min,参照RNA提取步骤获得总RNA,并取1 μL用DNA2000检测纯度。根据试剂盒说明书,取2 000 ng总RNA配置20 μL反转录体系。将反应体系先在42℃孵育40 min,然后转入80℃水浴锅继续孵育5 min后置于-20℃冰箱终止反应并保存。
1.2.3 猴源DDX3X基因扩增 以猴源肾细胞cDNA为模板,构建20μL PCR反应体系(2×Es Taq Master Mix10μL,DDX3X-EcoR I-F 0.5μL,DDX3X-Xho I-R 0.5μL,模板2μL,灭菌水7μL),并在PCR仪中94℃ 预变性4 min;94℃ 变性40 s,54℃ 退火30 s,72℃ 终止1 min,35个循环;在72℃孵育10 min终止反应,反应结束后用1.2%琼脂糖凝胶检测。
1.2.4 猴源DDX3X基因克隆及鉴定 根据目的基因序列大小,切割特异性条带,运用琼脂糖凝胶回收试剂盒回收PCR产物。将纯化产物于pMD-19T载体16℃水浴锅连接过夜(胶回收产物4.5μL,pMD-19T 0.5 mL,Solution I 5μL)。取连接产物转化至DH5α感受态细胞中(冰浴30 min,42℃热激90 s,冰浴3 min,加入900μL LB培养液,于37℃ 160 r/min摇床摇1.5 h,5 000 r/min,离心5 min,取200 μL转化产物涂布至氨苄抗性的LB培养基中,37℃培养16~24 h),挑取4个单菌落进行培养,并进行菌液PCR及重组质粒的双酶切鉴定。反应体系为:重组质粒8 μL,10×Buffer 2 μL,EcoR I 1 μL,EcoR I,Xho I 1 μL,灭菌水8 μL。
1.2.5 猴源DDX3X基因序列及蛋白质特性分析 使用Megalign及MEGA6软件进行基因同源性比对及遗传进化树的构建,参照在线软件Protpara(https: //web.expasy.org/cgi-bin/protparam/protparam)分析猴源DDX3X蛋白理化性质,SMART(http://smart.embl-heidelberg.de/smart/change_mode.PL)分析猴源DDX3X蛋白的功能结构域;SOPMA (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.PLpage= npsa_sopma.html)分析猴源DDX3X蛋白的二级结构;SWISS MODEL(http://swiss model.expasy.org/interactive)分析猴源DDX3X蛋白的三级结构;STRING(https://string-db.org/cgi/input.pl)预测分析DDX3X蛋白的相互作用蛋白。
2 结果与分析
2.1 猴源DDX3X基因PCR扩增
以猴源肾细胞cDNA为模板进行RT-PCR扩增,琼脂糖凝胶电泳检测显示,在2 000 bp左右出现单一目的条带(图1),与预期结果相符合。
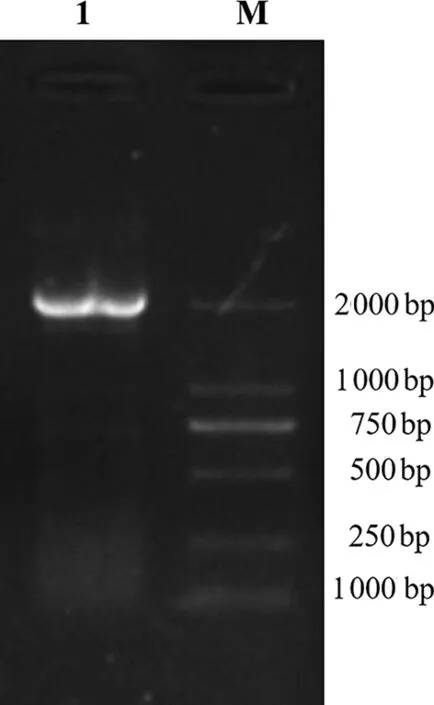
注:M:DL2000 Marker;1:猴源DDX3X基因。
Note: M,DL 2000 Marker; 1,monkeyDDX3Xgene.
图1 猴DDX3X基因PCR扩增结果
Fig.1 PCR amplification of monkeyDDX3Xgene
2.2 猴源DDX3X基因克隆及鉴定
在培养的4个菌落中,3个菌液出现预期大小的特异性目的条带。选择1号菌液进行扩大培养,提取质粒进行双酶切鉴定,酶切结果出现2 000 bp左右目的条带与载体条带(图2)。将已鉴定的阳性质粒送华大基因有限公司测序,测序结果显示,猴源DDX3X基因核苷酸序列大小为1 989 bp(图3)。
注:M:DL2000 Marker; 1、2、3、4均为猴源DDX3X基因。
Note: M,DL 2000 Marker; 1~4,monkeyDDX3Xgene
图2 猴源DDX3X基因克隆菌液PCR鉴定
Fig.2 PCR identification of monkeyDDX3Xgene
2.3 猴源DDX3X基因氨基酸序列分析
将测序所得猴源DDX3X氨基酸序列与GenBank中不同物种的DDX3X氨基酸进行同源性比,结果(图4)显示,猴源DDX3X氨基酸序列与猕猴同源性最高,为99.6%;与人、亚马逊松鼠猴、猫头鹰猴的同源性次之,分别为98.9%、98.2%和97.8%;与马、虎、犬、家猫、绵羊、野猪、牛、家兔、大熊猫等的同源性为91.2%~95.9%;与鸡、蛇、鱼、鸭、雁等的同源性为57.5%~77.3%。猴源DDX3X与哺乳动物的同源性较高,较为保守;而与非哺乳动物同源性相对较低。通过遗传进化树构建(图5)进一步发现,猴源DDX3X氨基酸序列与猕猴处于同一分支,亲缘关系最近。
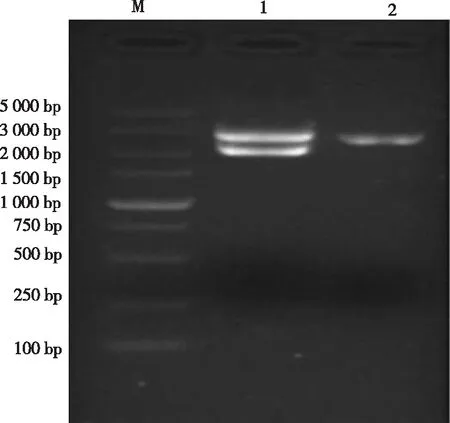
注:M:DL5000 Marker;1:重组质粒双酶切;2:空载体酶切鉴定。
Note: M,DL 5000 Marker; 1,double enzyme digestion of recombinant plasmid; 2,enzyme digestion identification of empty carrier
图3 猴源DDX3X基因重组质粒双酶切鉴定
Fig.3 Identification of monkeyDDX3Xrecombinant plasmid by double enzyme digestion

图4 猴源DDX3X氨基酸同源性比对

图5 猴源DDX3X氨基酸序列系统进化树
2.4 猴源DDX3X蛋白理化性质
蛋白理化性质分析显示,猴源DDX3X基因共编码662个氨基酸,理论蛋白分子质量为73 243.42 kDa,蛋白分子式为C3189H4962N938O1008S21,总原子数10 118个,理论等电点为6.73。猴源DDX3X蛋白是由20种氨基酸(甘氨酸含量最高,为11.5%)组成的脂溶性不稳定蛋白。
2.5 猴源DDX3X蛋白高级结构
蛋白质二级及三级结构预测结果表明,猴源DDX3X蛋白二级结构以α螺旋和无规则卷曲为主,分别占50.45%和28.4%;而β-折叠及β-转角的占比较少,分别为14.2%和6.95%。通过三级结构模型进一步表明该蛋白以α螺旋和无规则卷曲为主(图6),与二级结构预测相符。

图6 DDX3X蛋白质三级结构模型
Fig.6 Protein tertiary structure model of monkeyDDX3Xgene
2.6 猴源DDX3X蛋白功能结构域预测
蛋白功能结构域预测结果(图7)表明,猴源DDX3X蛋白主要存在2个核心功能结构域,分别是分布在199~418位的是DEAD-Like解旋酶超家族(DEXDc)氨基酸结构域及455~536位的解旋酶超家族C端(HELICc)氨基酸结构域。

图7 猴源DDX3X蛋白的保守功能结构域
Fig.7 Conservative functional domain of monkey DDX3X protein
2.7 DDX3X蛋白相互作用蛋白预测
蛋白相互作用蛋白预测结果(图8)显示,猴源DDX3X蛋白与丝氨酸/苏氨酸蛋白激酶(TBK1)、细胞分裂周期蛋白5(CDC5L)、核因子kappa B激酶(IKBKE)、真核起始因子4E家族(EIF4E)、U5小核核糖核蛋白组分亚型a(EFTUD2)、泛素特异性肽酶9(USP9X)、前mRNA处理因子6(PRPF6)、核翻译起始因子1A(EIF1AX)及Aly/REF输出因子(THOC4)等调节细胞进程、RNA加工等关键环节的蛋白均存在相互作用。

图8 猴源DDX3X蛋白的相互作用蛋白
Fig.8 Inter-acting protein of monkey DDX3X protein
3 结论与讨论
研究成功从猴源肾细胞中扩增获得猴源DDX3X基因,基因全长1 989 bp,编码662个氨基酸。生物信息学分析显示,猴源DDX3X基因氨基酸序列与猕猴同源性最高,为99.6%;与人及其他猴类次之,与其他非哺乳动物同源性较低,表明其保守性较强。蛋白质理化性质预测显示,DDX3X蛋白理论分子质量为73 243.42 kDa,是由20种氨基酸组成的脂溶性不稳定蛋白。DDX3X蛋白质二级结构以α螺旋和无规则卷曲为主(50.45%和28.4%),而β-折叠及β-转角较少(14.2%和6.95%),分别存在DEAD-Like解旋酶超家族(DEXDc)氨基酸结构域及解旋酶超家族C端(HELICc)氨基酸结构域,并且与调控细胞周期、调节RNA加工等多种关键蛋白存在相互作用。
DDX3X是DEAD-box RNA解旋酶家族的成员,位于人类X染色体p11.3~p11.23上[10-11]。有研究报道,DDX3X参与多种基因调控事件,包括转录调控、RNA解旋、剪接、RNA核输出、核糖体生物发生和mRNA翻译[12]。因此,DDX3X被认为参与基因表达的表观遗传调控,研究显示,DDX3X虽然在物种之间高度保守,但发挥的功能千差万别,如小鼠和人的DDX3X同源型高达99%,而行使的功能却不一致[13]。已有研究者将DDX3X基因作为癌症治疗的靶基因来治疗肺癌患者体内EGFR激活的异质性[14]。
将克隆的序列进行同源性比对及遗传进化树的分析发现,DDX3X基因氨基酸序列在哺乳动物均有较高相似性,证明该基因在物种进化过程中高度保守。蛋白质功能结构域预测显示,猴源DDX3X蛋白存在DEAD-Like解旋酶超家族(DEXDc)氨基酸结构域及解旋酶超家族C端(HELICc)氨基酸结构域。在此之前,有研究者已报道过其他物种的DDX3X均含有类似的2个功能结构域,并且参与机体多种生物学调节过程[15-16]。相关研究表明,DDX3X参与了HIV、VSV、VACV、HCV和HBV等5个病毒的复制过程,作为主要表达于细胞质的蛋白,通过将猪蓝耳病病毒PRRSV感染PAM细胞后可上调DDX3X基因的表达并抑制病毒的复制,并且DDX3X可通过调节干扰素β启动刺激因子及丝氨酸/苏氨酸蛋白激酶的表达来介导β干扰素的产生,从而诱发机体的免疫应答以对抗病毒的感染及复制[17]。
通过同源性比对及遗传进化树证明DDX3X基因序列在不同物种进化过程中高度保守,而其保守结构域主要是DEXDc氨基酸结构域及HELICc氨基酸结构域,而该结构域正是该蛋白行使主要生物学进程的功能区。成功克隆获得的猴源DDX3X基因与刘琴等克隆获得的猪DDX3X基因编码氨基酸数量一致,进一步证明其在不同物种中保守性较高[17]。 在禽类动物的研究中,研究者将鸭坦布苏病毒(DTMUV)感染BHK-21细胞之后进行转录组及蛋白组学分析,DDX3X基因作为差异基因被发现,通过荧光定量PCR及WB验证发现,DDX3X及DDX5基因mRNA水平及蛋白水平均受到抑制。此外为验证DDX3X基因参与病毒复制过程,研究者向BHK-21细胞转染DDX3X基因特异性干扰siRNA,并于转染后用DTMUV感染BHK-21细胞,通过血凝和血凝抑制试验发现,干扰DDX3X基因后,病毒的复制能力增强[18]。
目前虽然发现猪DDX3X基因的表达对PRRSV易感及相关病毒复制存在关联性,但具体相互作用机制还不够清晰[19-20]。由于病毒研究存在较高不可控性,利用活体研究病毒的相关生物学过程代价较高,本次成功从猴源肾细胞中扩增到DDX3X基因,并对其进行基因同源性及相关蛋白质性质、高级结构及保守功能结构域及相互作用蛋白的分析,将为后期厘清DDX3X基因介导的病毒易感及复制的相关机制提供理论数据。
猜你喜欢
红蜻蜓·低年级(2022年5期)2022-05-11
幼儿园(2021年16期)2021-12-06
红领巾·探索(2020年5期)2020-05-19
生物信息学(2020年1期)2020-05-16
家教世界·创新阅读(2020年12期)2020-01-11
中国洗涤用品工业(2019年4期)2019-05-11
广州大学学报(自然科学版)(2019年1期)2019-05-07
中成药(2018年1期)2018-02-02
中成药(2017年3期)2017-05-17
天津科技大学学报(2016年1期)2016-02-28
