
茶树根际细菌菌株对种子萌发及幼苗生长的促生效应及机制初探
2020-01-07 03:38韩丽珍邓兆辉朱春艳
种子 2019年12期
韩丽珍,2, 刘 畅, 邓兆辉, 朱春艳
(1.贵州大学生命科学学院, 贵阳 550025;2.贵州大学/山地植物资源保护与种质创新教育部重点实验室, 贵阳 550025)
植物根际促生菌(plant growth-promoting rizobacteria, PGPR)是指以定殖或自由附着的方式生活在植物根际的一类细菌总称[1]。它能改变根际土壤的理化性质、提高根际土壤中对植物生长有利的营养元素的转化[2],也可通过分泌铁载体、有机酸、生物表面活性剂、植物生长激素等,直接或间接促进植物的生长[3]。不同植物根际分离到的促生菌菌株对相应植物的种子萌发可能具有一定的促进作用。从柳枝稷根茎中分离筛选到可分泌IAA的Pseudomonas和Rhizobium细菌菌株,接种后可以提高盐胁迫下柳枝稷种子的发芽率,促进胚根和胚芽的生长[4]。从西藏黑青稞根际筛选到的4株菌株对青稞种子发芽促进作用显著,但不同菌株影响有差异[5]。而从大豆种子中分离到的内生芽孢杆菌菌株,大部分均表现出促进大豆发芽的作用,其中促生作用最好的SN 10 E 1菌株为巨大芽孢杆菌[6]。也有报道表明促生作用并不仅仅局限在同种属的植物中。李青梅等发现,经胶质芽孢杆菌菌剂处理的黄瓜、番茄、茄子及辣椒等4种蔬菜种子的发芽率均高于对照,但对蔬菜幼苗根生长的影响因蔬菜种类不同而异[7]。而从油菜根际土壤中分离到的3株菌N-1、N-15、K-25可使桔梗种子的发芽率和幼苗茎长明显高于对照,也证明从一种植物根际能分离到对另一种植物有促生作用的功能菌,促生菌在不同植物根际的分布和作用存在交叉现象[8]。在前期研究中,本实验室从茶树根际分离筛选到4株细菌菌株,其中PseudomonashunanensisGD 3具有溶磷解钾能力,AgrobacteriumradiobacterKKS-6-N 1菌株具有固氮能力,2个菌株对白菜、空心菜及苋菜具有促生作用[9-10]。P 5菌株是采集茶树根际土壤、以阿须贝无氮培养基筛选获得的1株细菌,具有一定的固氮效能,而其余的促生特性还未知。本研究拟通过P 5菌株与2株具优良促生性能的GD 3和KKS-6-N 1进行浸种试验,比较不同菌株对辣椒和花生种子萌发的影响;并通过灌根处理盆栽花生幼苗,研究P 5菌株对花生根际土壤微生物功能的影响效应,从根本上解析P 5菌株的促生机制。
1 材 料
1.1 供试菌株及材料
3株茶树根际细菌菌株GD 3、KKS-6-N 1和P 5(未知种属)均为本实验室从不同环境茶树根际土壤中分离获得,保存于本实验室。
1.2 供试材料与土壤
供试花生及辣椒种子均为当地市售种子。土壤来源于贵州省贵阳市花溪区贵州大学南校区农田土壤(26°42′48″N,106°67′31″E),去除表面枯枝浮土后,采集距表面20~50 cm处土壤,过筛后用于盆栽试验。
1.3 培养基
LB培养基、高氏一号培养基及马丁培养基按《微生物学实验教程》[11],参考Mab等[12]的方法配制蛋白胨氨化培养基、吴翔等[13]的方法配制阿须贝氏无氮培养基,NBRIP培养基参考Nautiyal等[14]的方法,亚历山大硅酸盐培养基参考何琳燕等[15]的方法。
2 方 法
2.1 菌种的活化及菌液的制备
将保存于-80 ℃超低温冰箱中的GD 3、KKS-6-N 1和P 5菌液取出适量,接种于LB培养基中,于30 ℃恒温摇床中,150 r·min-1振荡培养过夜活化;翌日取活化的菌液再次转接于LB培养基中,相同条件下培养24 h,菌液测定OD600值,以无菌LB调整该值为1.0备用。
2.2 菌株浸种处理对辣椒种子萌发及盆栽试验的影响
将辣椒种子置于3%次氯酸钠溶液中进行表面消毒10 min,用蒸馏水反复清洗多次后,处理组的辣椒种子分别置于OD600值为1.0的3种菌液中浸种2 h,未处理组对照种子则置于LB培养基中;浸种完成后用无菌滤纸略微蘸干表面液体、置于铺有被无菌水润湿的无菌滤纸的培养皿中,于30 ℃恒温培养箱中放置15 d,期间保持滤纸湿润,统计辣椒种子的发芽率;之后移栽于装有300 g土壤的育苗盆中进行盆栽试验,整个试验期间保持土壤湿润,30 d后收获测定植株生长指标。试验设对照组、GD 3处理组、KKS-6-N 1处理组和P 5处理组,每组为25粒种子,重复3次;盆栽试验时每组6株幼苗,重复3次。
2.3 菌株浸种处理对花生种子萌发的影响
将花生种子置于20%过氧化氢溶液中进行表面消毒20 min,经蒸馏水反复清洗后浸泡6~12 h,以无菌滤纸略微蘸干后置于3种菌株的菌液中进行2 h浸种处理,对照种子则置于LB培养基中;待浸种完成后,置于铺有滤纸的培养皿中,30 ℃恒温培养箱中放置3 d,统计花生种子的发芽率。种子试验设计及处理同上。
表1 3株茶树根际细菌浸种对辣椒种子萌发及幼苗生长的影响
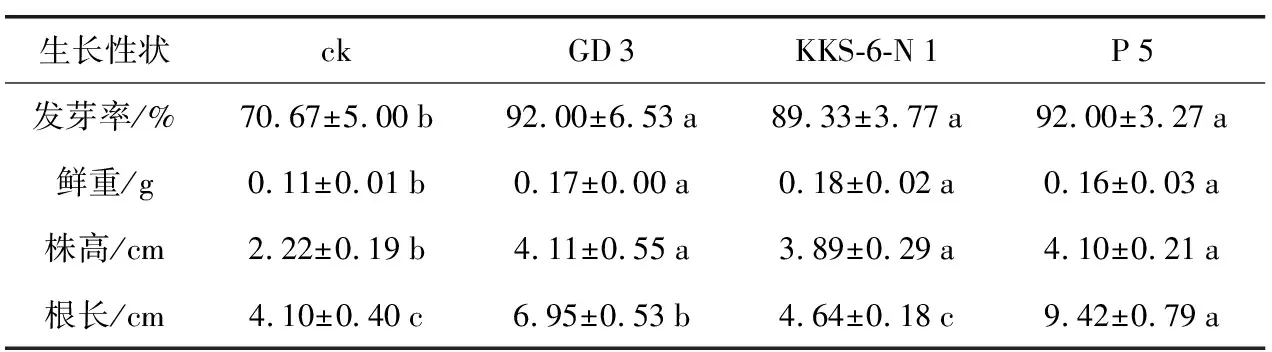
生长性状ckGD3KKS-6-N1P5发芽率/%70.67±5.00b92.00±6.53a89.33±3.77a92.00±3.27a鲜重/g0.11±0.01b0.17±0.00a0.18±0.02a0.16±0.03a株高/cm2.22±0.19b4.11±0.55a3.89±0.29a4.10±0.21a根长/cm4.10±0.40c6.95±0.53b4.64±0.18c9.42±0.79a
注:同一行不同小写字母代表不同处理之间差异显著(p<0.05)。下同。
2.4 菌株灌根处理对花生幼苗生长的影响
将花生种子经过氧化氢消毒,蒸馏水清洗浸泡后,置于铺有无菌滤纸的培养皿中,30 ℃条件下3 d待种子萌发后,选择长势相当的花生幼苗移栽于育苗盆中,以未接种菌株为对照,设置GD 3接种组、KKS-6-N 1接种组和P 5接种组,每2 d采用灌根法接种菌液,接种量为每次5 mL,对照组则用等量无菌LB培养基代替,每组均设6个重复,常规方式管理。30 d后测定花生株高、鲜重、根重及叶绿素含量等生长及生理指标,叶绿素含量采用Arnon法测定。
2.5 P 5菌株灌根对花生营养指标的影响
盆栽实验30 d后,测定P 5接种组及对照组花生植株的营养指标,植株全氮含量测定采用凯氏定氮法,全磷含量测定采用钼锑抗比色法,全钾含量测定采用火焰分光光度法[16]进行。
2.6 P 5菌株灌根对花生根际土壤三大菌群及功能菌群的影响
30 d盆栽实验后,利用抖根法收集P 5处理组的花生根际土壤,测定土壤三大菌群(细菌总数、放线菌总数及真菌总数),功能菌群数量的测定包括氨化细菌、固氮菌、溶磷菌及解钾菌等4种类型。细菌、放线菌及真菌总数分别以LB固体培养基、高氏一号培养基及马丁培养基测定,采用稀释平板涂布法进行计数;以蛋白胨氨化培养基培养氨化细菌,采用最大近似值法(MPN, most probable numbers)测定;固氮菌、溶磷菌及解钾菌分别以阿须贝无氮培养基、NBRIP培养基及亚历山大硅酸盐培养基培养,以稀释涂布平板法计数。
2.7 P 5菌株灌根对花生根际土壤酶活性的影响
采集P 5灌根处理30 d的花生根际土壤进行土壤酶活性测定。土壤蔗糖酶酶活的测定采用硫代硫酸钠滴定法,单位酶活是指每克干土所消耗的硫代硫酸钠(0.05 mol·L-1)体积(mL·g-1)[17];过氧化氢酶酶活采用高锰酸钾滴定法,单位酶活是指每克干土于20 min内所消耗的高锰酸钾(0.1 mol·L-1)体积(mL·g-1);脲酶酶活的测定采用苯酚钠-次氯酸钠比色法,单位酶活是指37 ℃条件下、24 h处理后每克风干土壤中的氨氮含量(mg·g-1)[18];中性磷酸酶酶活是指每克干土经24 h处理后释放酚的体积(mg·g-1),参照吴金水等[19]的方法进行测定。
2.8 P 5菌株的分子生物学鉴定及系统发育树构建
P 5菌株的初步鉴定采用分子生物学方法进行。利用细菌DNA提取试剂盒抽提菌株的DNA,以细菌16 S rRNA基因的通用引物对27 F/1492 R进行PCR扩增,上、下游引物序列分别为27 F(5′-AGAGTTTGATCMTGGCTCAG-3′),1492 R(5′-TACGGYTACCTTGTTACGACTT-3′)[20]。PCR扩增体系及扩增条件参考王欢等[10]的方法。扩增产物经琼脂糖凝胶电泳,检测到有既定大小的条带(约1 500 bp)送至上海英骏生物工程公司测序。获得的核苷酸序列经NCBI网站上BLAST进行在线同源性比较,选择相似性最高的结果进行分析。选取克雷伯氏菌属的不同种菌株用于系统发育树的构建,以PseudomonasmooreiRW 10 T(AM 293566)作外群。采用Clustal W 2进行多重序列比对、Mega 6.0构建系统发育树,进行1 000次自展检验[21]。
2.9 数据分析与处理
采用SPSS 20.0软件,以LSR多重比较对数据进行分析。
3 结果与分析
3.1 3株茶树根际细菌浸种对辣椒种子萌发及幼苗生长的影响
利用从茶树根际分离、筛选到的3株细菌菌株GD 3、KKS-6-N 1及P 5对辣椒进行浸种试验,萌发后于盆栽条件下继续生长30 d后测定幼苗的相应生长指标。以3株细菌菌液进行了2 h的浸种处理后,辣椒种子的萌发率均在90%左右,较对照显著提高了近20%,但不同菌株处理之间并无显著性差异(p<0.05)。30 d的盆栽实验表明,与对照相比,GD 3、KKS-6-N 1、P 5浸种处理的辣椒幼苗鲜重分别增加54.55%、63.64%和45.46%,株高分别增长85.14%、75.23%和84.69%,根长分别增长了69.51%、13.17%和129.76%。显然,3株茶树根际细菌的浸种处理使辣椒种子的发芽率明显提高,且对辣椒的株高和鲜重具有显著促生作用;其中,P 5浸种处理的植株根长明显高于对照及其余2个菌株的处理。
3.2 3株茶树根际细菌浸种对花生种子萌发的影响
利用3株茶树根际细菌菌株对花生进行相同的浸种处理,3 d后统计种子的萌发率(图1),结果发现,菌剂浸种处理组的发芽率均在90%以上,较对照种子的发芽率提高了20%以上,3株细菌菌株对种子的萌发具有明显的促进效果,但不同菌株处理之间对种子萌发率并无显著差异(图1)。
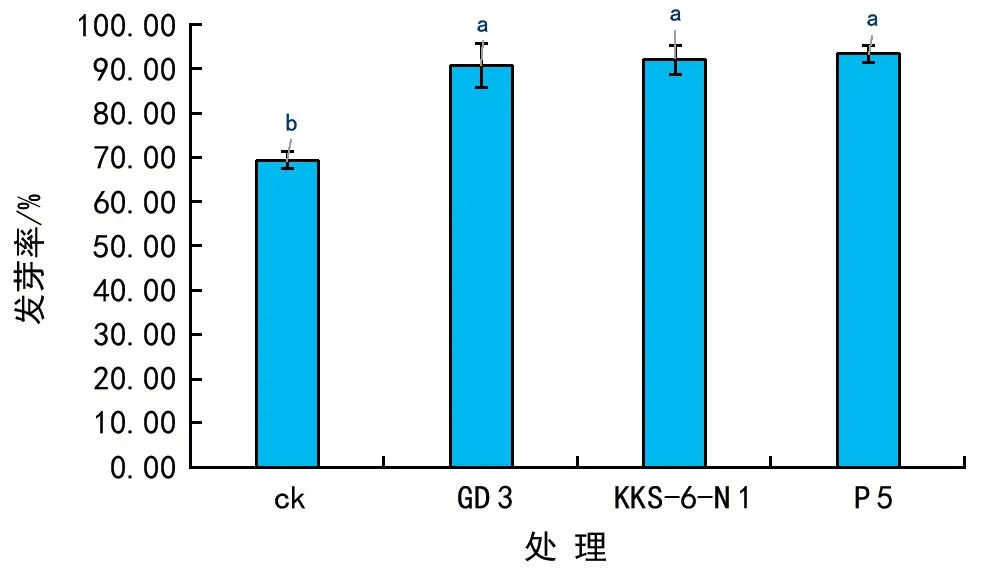
图1 3株茶树根际细菌对花生种子萌发的影响
3.3 3株茶树根际细菌灌根处理对花生幼苗生长的影响
为了进一步探讨根际细菌对花生幼苗生长的影响作用及内在机制,分别利用3株菌液进行灌根处理,培育30 d后测定花生的生长性状(表2),发现3株菌株对花生幼苗生长的影响呈现一定差异。与对照相比,GD 3、KKS-6-N 1、P 5灌根处理的花生幼苗鲜重分别增长4.63%、26.83%和35.12%,株高分别增加了5.99%、16.22%和32.73%,根重分别增长21.62%、29.73%和35.14%。生理指标的测定结果显示,GD 3和KKS-6-N 1接种组的叶绿素含量略低于对照,而P 5接种组的叶绿素含量较对照提高了13.47%。结果(图2)表明,P 5菌株的灌根处理对花生幼苗的促生效果显著,其次为KKS-6-N 1。
表2 3株茶树根际细菌对花生幼苗生长的影响
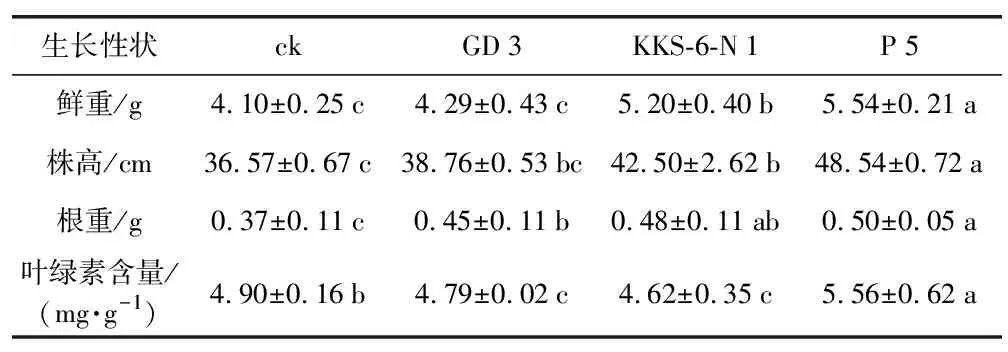
生长性状ckGD3KKS-6-N1P5鲜重/g4.10±0.25c4.29±0.43c5.20±0.40b5.54±0.21a株高/cm36.57±0.67c38.76±0.53bc42.50±2.62b48.54±0.72a根重/g0.37±0.11c0.45±0.11b0.48±0.11ab0.50±0.05a叶绿素含量/(mg·g-1)4.90±0.16b4.79±0.02c4.62±0.35c5.56±0.62a
3.4 P 5菌株灌根处理对花生植株营养元素的影响
由于P 5菌株可以显著促进花生幼苗鲜重和株高的增加,因而经P 5灌根处理的花生植株进行了营养指标测定(表3),发现接种组植株的全氮增高了13.56%,与对照呈现显著差异;全钾含量略有提升,但差异不明显,而全磷含量基本无改变(p<0.05)。

注:图左3株花生为P 5菌株处理,图右3株为ck。图2 根际促生菌菌株P 5对花生幼苗生长的影响
表3 根际促生菌菌株P 5对花生植株营养元素的影响
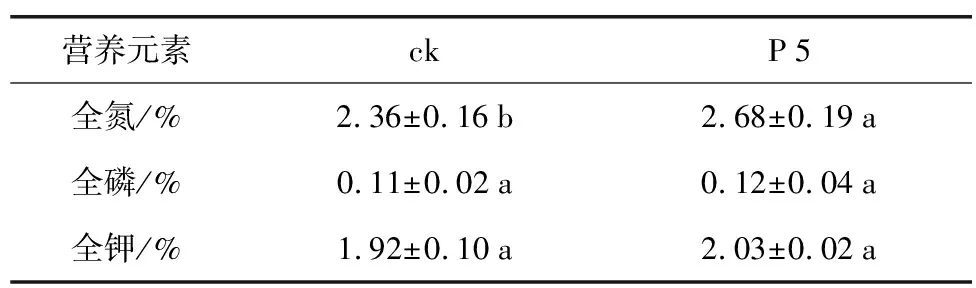
营养元素ckP5全氮/%2.36±0.16b2.68±0.19a全磷/%0.11±0.02a0.12±0.04a全钾/%1.92±0.10a2.03±0.02a
3.5 P 5菌株灌根处理对花生根际土壤三大菌群和功能菌群数量的影响
由于土壤微生物在土壤营养物质循环及维持土壤肥力方面起着重要作用,实验拟进一步从土壤菌群数量和土壤酶活入手,解析该菌株对花生的促生机制。对P 5菌株灌根处理的花生根际土壤及对照土壤进行三大菌群总数的测定(表4),结果发现,处理组根际土壤中的细菌总数较对照增长了2.63倍,真菌总数增长了2倍,细菌数/真菌数之比增加了1.32倍,菌剂处理使得细菌和真菌数量较对照呈现显著差异,而放线菌总数并无明显差异(p<0.05)。由于P 5处理后的花生根际土壤中细菌总数显著提高,因而对根际土壤中与促生相关的功能菌群数量进行了测定(表4),发现氨化细菌及固氮菌数分别较对照增高了2倍和2.44倍,溶磷菌数增高了4.07倍,与土壤氮磷含量直接相关的氨化细菌、固氮菌及溶磷菌数均明显高于对照,而解钾菌数略有下降,但差异不显著(p<0.05)。
3.6 P 5菌株灌根处理对花生根际土壤酶活性的影响
酶活性的测定结果(表5)显示,与对照相比,处理组根际土壤的蔗糖酶活性增加了1.41倍,过氧化氢酶活性增长了2.65倍,脲酶增长了2.33倍,中性磷酸酶活性基本无改变,表明P 5浸种处理的花生根际土壤中,蔗糖酶、过氧化氢酶及脲酶酶活性均较对照有显著提高(p<0.05)。
表6 P 5菌株的16 S rRNA基因相似性比对
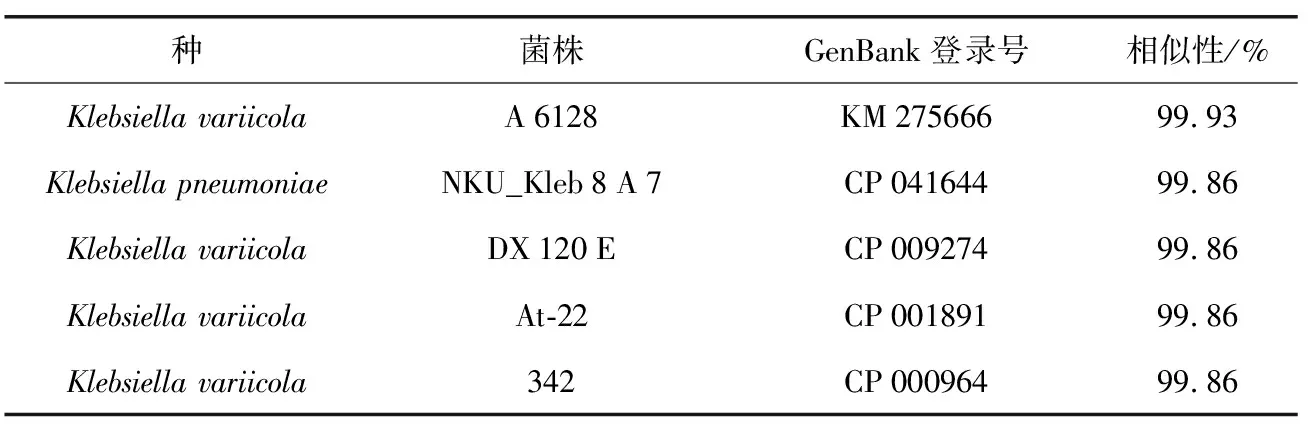
种菌株GenBank登录号相似性/%Klebsiella variicolaA6128KM27566699.93Klebsiella pneumoniaeNKU_Kleb8A7CP04164499.86Klebsiella variicolaDX120ECP00927499.86Klebsiella variicolaAt-22CP00189199.86Klebsiella variicola342CP00096499.86
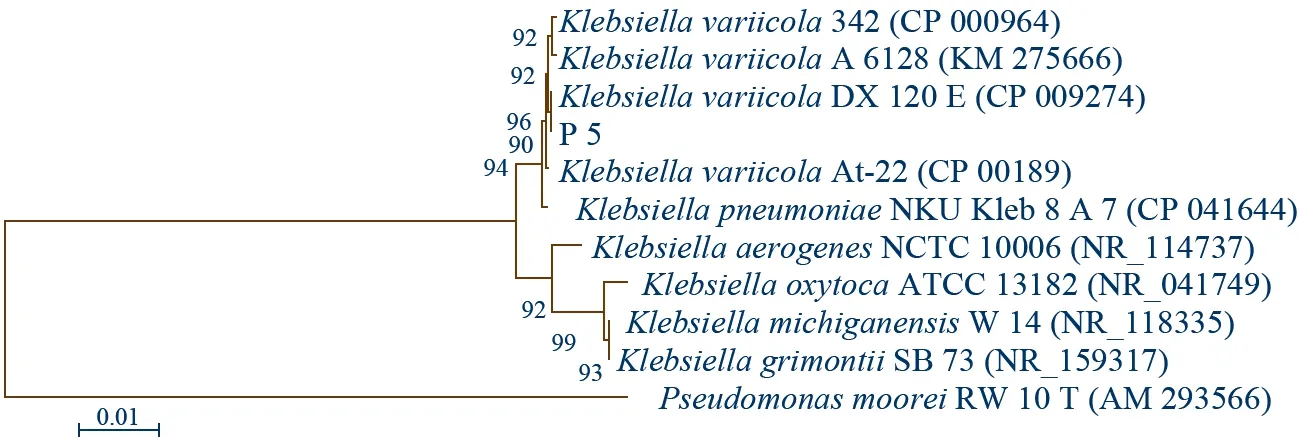
图3 P 5菌株的16 S rRNA基因系统发育树
表4 根际促生菌菌株P 5对花生根际土壤三大菌群数量(cfu·g-1)的影响
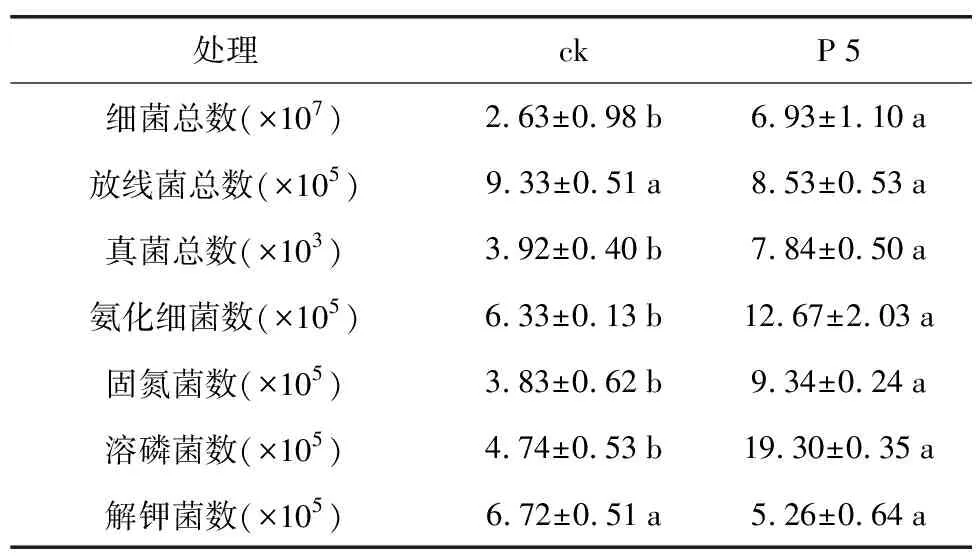
处理ckP5细菌总数(×107)2.63±0.98b6.93±1.10a放线菌总数(×105)9.33±0.51a8.53±0.53a真菌总数(×103)3.92±0.40b7.84±0.50a氨化细菌数(×105)6.33±0.13b12.67±2.03a固氮菌数(×105)3.83±0.62b9.34±0.24a溶磷菌数(×105)4.74±0.53b19.30±0.35a解钾菌数(×105)6.72±0.51a5.26±0.64a
表5 根际促生菌菌株P 5对花生根际土壤酶活性的影响
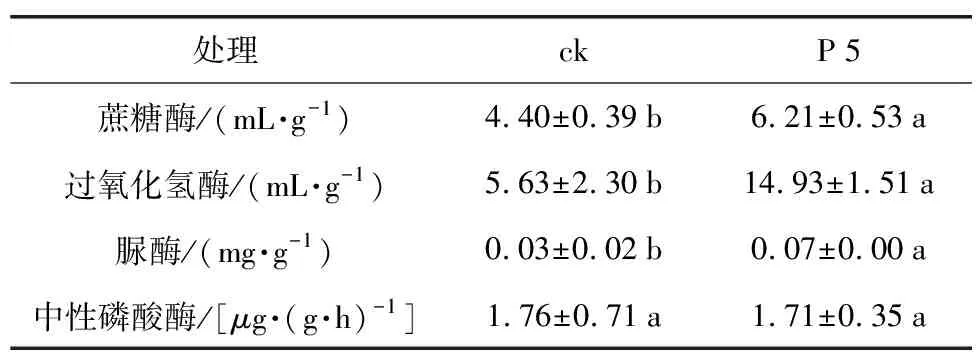
处理ckP5蔗糖酶/(mL·g-1)4.40±0.39b6.21±0.53a过氧化氢酶/(mL·g-1)5.63±2.30b14.93±1.51a脲酶/(mg·g-1)0.03±0.02b0.07±0.00a中性磷酸酶/[μg·(g·h)-1]1.76±0.71a1.71±0.35a
3.7 P 5菌株的分子鉴定及系统发育树的构建
利用细菌16 S rRNA基因的通用引物对27 F/1492 R对P 5细菌的总DNA进行PCR扩增,测序结果获得1 405 bp的核苷酸序列,经NCBI网站上进行BLAST比对,发现与其一致性最高的菌株为克雷伯氏菌属、变栖克雷伯氏菌(Klebsiellavariicola),与该种A 6128菌株的16 S rRNA基因相似性为99.93%,此外与多株变栖克雷伯氏菌菌株及肺炎克雷伯氏菌 NKU_Kleb 8 A 7菌株的相似性均为99.86%(表6)。与同属内的不同种构建系统发育树(图3),可见P 5菌株与Klebsiellavariicola的不同菌株聚于一大枝,且与该种DX 120 E菌株(CP 009274)的16 S rRNA基因在系统树上相邻,因而结合分子鉴定及系统发育树,可将P 5菌株鉴定为Klebsiellasp.。
4 结论与讨论
本研究中使用的3株细菌菌株GD 3、KKS-6-N 1和P 5均是从茶树根际分离筛选获得的,浸种试验表明,3株细菌均可使辣椒及花生种子的发芽率达到90%以上,较对照相比,发芽率可提高20%左右,对辣椒及花生种子的萌发均具有显著的促进作用。在农业生产上,目前的花生栽培通常是种子经拌种或包衣后进行播种,利用菌剂进行种子包衣处理不失为一个提高种子萌发的有效方式。而辣椒种子常因成熟度不够、采种技术不当或贮藏方法等原因造成发芽率降低[22],研究结果为利用3株细菌菌剂进行拌种或包衣处理奠定了基础。
但是,促进种子萌发的菌株不一定会对植株的生长有显著促进作用。张振建等报道,乌拉尔杆菌PN 133和土壤根瘤杆菌Y 42的浸种均可使苋菜和油麦菜的发芽率提高,而灌根处理则促生效果并不明显[23]。利用2株溶磷菌进行青稞种子的萌发试验中,菌株10 BN-11对增加青稞发芽率及促进生长效果明显,菌株12-BN-6对株高、根长和鲜重具有明显的促进作用,但可抑制种子的萌发[24]。王艳燕等的研究发现,在9种可促进辣椒发芽率提高的菌株中,后续的穴盘育苗有促生效果的仅有4株菌,其中对辣椒下胚轴和主根长度影响显著的GH 6-1和GH 5-3并未在苗期表现出显著的促生作用,说明有些菌虽能促进种子萌发,但未必能在后期幼苗生长过程中促进其对养分的吸收[25]。本研究的浸种实验虽显示3株细菌均可明显提高辣椒和花生种子的发芽率,且不同菌株的处理并无显著差异;但辣椒种子萌发后的生长过程中,3株细菌对其后续的影响略有不一致;而且,在利用菌液的灌根处理中,发现P 5菌株对花生幼苗的促生效果显著高于KKS-6-N 1,GD 3对幼苗株高及鲜重的影响则并不明显,这也与报道的相一致。因而对于根际细菌促生作用的研究中,有必要分别对种子萌发和幼苗生长进行研究,综合评判根际细菌菌株的促生作用。
研究显示,根际促生菌可提高种子的发芽率和芽长,进而提高植株的生物量,但植物对所分离的功能菌株是否具有根际效应,是菌株发挥功能的关键[26]。康贻军等在评价PseudomonaschlororaphisRA 6 和BacilluspumilusWP 8浸种和拌土对豇豆生长的作用时,发现菌液的浸种处理及108cfu·g-1剂量的菌株拌土处理均可显著提高豇豆的株高,认为浸种处理从原理上讲,应该不会对非根际土壤产生较大影响[27]。Probanza等研究表明,PGPR对植物土壤中土著微生物数量及群落结构的影响可能是其促生的另一个机制[28]。研究表明,土壤酶活性与土壤速效养分密切相关,而土壤微生物数量大、繁殖快、活性强,直接影响到植物根际有机质的分解和养分的转化[29]。因而,为了进一步解析和探究P 5菌株对花生幼苗生长的促进机制,本研究对接种P 5菌株的花生根际土壤进行了土壤微生物及酶活的测定。由于细菌、放线菌和真菌是土壤微生物的三大类群,构成了土壤微生物的主要生物量,它们的区系组成和数量变化能反映土壤生物活性水平[30]。研究结果显示,P 5处理组根际土壤中的细菌总数较对照增长了2.63倍,真菌总数增长了2倍,细菌总数/真菌总数之比增加了1.32倍,而放线菌总数并无明显差异(p<0.05)。多数报道表明,接种促生菌均可不同程度地提高土壤细菌总数和放线菌数,降低真菌数量[31-33]。王兴祥等研究认为,细菌型土壤是土壤肥力提高的一个生物指标,真菌型土壤是地力衰竭的标志[34]。但在巨尾桉接种固氮菌或解钾菌的土壤中,发现细菌及放线菌数量增多,而真菌数量均不同程度的增高[35]。在研究中,真菌数量虽有明显提高,但细菌数/真菌数之比仍高于对照,接种组土壤仍表现为细菌型土壤。土壤酶活性在一定程度上可以反映土壤肥力状况。土壤蔗糖酶活性强弱能反映土壤的熟化程度和肥力水平[36],过氧化氢酶活性与土壤呼吸作用及土壤微生物活动密切相关[37],脲酶活性可在一定水平上反映土壤的供氮水平与能力[38],磷酸酶在土壤有机磷向无机磷的分解转化方面起着重要作用[39]。研究显示,除中性磷酸酶外,P 5接种组的根际土壤蔗糖酶、脲酶及过氧化氢酶均显著高于对照,表明根际细菌P 5的处理活跃了土壤微生物;对功能菌群数量的进一步测定结果表明,氨化细菌数、固氮菌数及溶磷菌数显著增高(p<0.05)。氨化细菌及固氮菌数量的显著提高可以提供植物更多所需的氨氮,接种组植株全氮含量的明显提高与土壤脲酶等酶活性的增高、氨化细菌及固氮菌数的显著提升相一致,这显然是促进花生生长的原因之一。多项研究也已证实土壤酶活性与大豆、小麦及玉米产量呈显著正相关关系[40-41]。此外,P 5菌株经分子鉴定为克雷伯氏菌属,与KlebsiellavariicolaDX 120 E的同源性最高;而Klebsiellavariicola是2004年才发现的新种,作为内生菌,并由于其优良的固氮性能被报道[42-43];本研究中的P 5菌株是具有一定固氮能力的茶树根际细菌,因而接种了克雷伯氏菌属的P 5菌株后,显著提升了花生的全氮含量。值得注意的是,P 5菌株处理的花生全磷含量与对照基本无差异,这与根际土壤中性磷酸酶活无显著变化相一致,而溶磷菌数明显增高的原因还有待进一步研究。
综上所述,Klebsiellasp.P 5菌株的浸种处理可以显著促进辣椒和花生种子的萌发,灌根处理可以显著促进花生幼苗的生长,植株的全氮含量明显提高,这与根际土壤氨化细菌及固氮菌数量的显著增多,土壤蔗糖酶、脲酶及过氧化氢酶活性的显著增高直接相关,研究结果为下一步利用P 5菌株进行拌种或田间施用奠定了理论基础。
猜你喜欢
浙江农业科学(2022年7期)2022-07-15
中国土壤与肥料(2021年5期)2021-12-02
四川农业科技(2021年7期)2021-12-01
中国瓜菜(2021年4期)2021-06-02
江苏农业科学(2021年5期)2021-04-22
中国果业信息(2020年10期)2020-12-15
农业知识(2020年22期)2020-11-27
种子科技(2020年8期)2020-05-26
上海农业科技(2020年2期)2020-04-02
山西农业科学(2020年1期)2020-02-27
