
人p54nrb基因真核表达载体的构建及其在HepG2细胞内的表达
2020-01-01 02:50:50陈春玲闫吕彬吴嫦丽张秀娟广东医科大学生理学教研室图书馆广东湛江5403
广东医科大学学报 2019年6期
陈春玲,闫吕彬,牛 伢,吴嫦丽,张秀娟*(广东医科大学 . 生理学教研室;. 图书馆,广东湛江 5403)
54 kDa核DNA、RNA结合蛋白(54kDa nuclear RNA-and DNA-binding protein,p54nrb)也称Non-POU domain containing octamer-binding protein(NONO),是一种主要分布于细胞核,且能与DNA、RNA结合的多功能核蛋白[1]。在细胞核内,p54nrb参与多种核内生物学过程,如初级mRNA的剪接[2]、DNA的损伤修复[3]、DNA解螺旋[4]、基因的转录调控[5]、缺陷RNA核滞留等[6],p54nrb基因突变或异常可引起广泛的基因mRNA谱表达异常。p54nrb是核蛋白体结构paraspeckle的核心组成蛋白[8-9],paraspeckle的功能是参与A-I超编辑RNA的核滞留过程。p54nrb核内功能的多样性决定了其生物功能的多样性,可参与生物节律的调节[10],并与神经系统发育[11]及肿瘤的发生、转移等密切相关[12-14]。为了进一步研究p54nrb基因的功能,我们构建了p54nrb基因真核表达载体,并观察了其在人肝癌细胞HepG2内表达,为后续研究p54nrb基因的生物功能提供载体。
1 材料和方法
1.1 材料
pcDNA3-HA及pcDNA3质粒、HepG2 细胞由广东医科大学生理学教研室保存。鼠抗HA单克隆抗体购自sigma公司;Alexa-Flour 488 标记的羊抗鼠IgG抗体购自中杉金桥。ReverTra Ace-α-™ kit、XhoI和KpnⅠ限制性核酸内切酶、Ligation High DNA连接酶、KOD-Plus-Neo酶购于TOYOBO公司;Lipofectamine 2000脂质体转染试剂购于Invitrogen公司;质粒提取试剂盒和凝胶回收试剂盒购于Aidlab 公司;总RNA提取试剂盒购于Axygen公司;胎牛血清及培养基购于Hyclone公司。
1.2 方法
1.2.1 HepG2细胞总RNA的提取 将对数生长期的HepG2细胞接种于细胞培养皿中,在细胞生长至80%~90%融合时,收取细胞,按Axygen总RNA制备试剂盒使用说明书提取总RNA。
1.2.2 RT-PCR扩增人p54nrb基因的蛋白编码序列提取总RNA,并以总RNA为模板反转录合成cDNA。取反转录生成的cDNA 2 μL,PCR扩增人p54nrb基因的蛋白编码序列,PCR反应的上游引物为:5'- CGG GGTACCATGCAGAGTAATAAAACTT TTAACTTG G-3';下游引物:5'- CCGCTCGAGGTATCG G CGA CGTTTGTTTGG -3';其中GGTACC为KpnI 酶切位点,CTCGAG为XhoI 酶切位点。PCR 反应条件设定为94 ℃、30 s,56 ℃、30 s,68 ℃、2 min,40个循环。用1%琼脂糖凝胶对PCR扩增产物进行电泳,使用天能凝胶图像分析系统对凝胶进行照相。使用Axygen DNA凝胶回收试剂盒切胶回收p54nrb PCR产物。
1.2.3 pcDNA3-p54nrb-HA载体的构建及鉴定 使用XhoI和KpnI限制性内切酶对p54nrb RT-PCR扩增产物及pcDNA3-HA质粒分别进行双酶切,并将酶切产物切胶回收,用DNA连接酶将酶切产物在16 ℃下连接16 h;然后将连接产物转化到DH5α感受态细菌,用氨苄霉素抗性的琼脂板进行阳性克隆的筛选。挑取阳性克隆,LB培养基中摇菌14 h,然后提取质粒。用XhoI和KpnI限制性内切酶对重组质粒进行双酶切初步鉴定,鉴定正确后再将该质粒送至华大基因公司测序。所构建的质粒命名为pcDNA3-p54nrb-HA。
1.2.4 细胞培养及质粒转染 HepG2细胞培养于含10%胎牛血清的RPMI 1640培养基。将1×104个HepG2细胞接种于Petri皿中,待细胞生长至融合度达到70%~80%时进行质粒转染,其过程如下:将0.2 μg pcDNA3-p54nrb-HA或pcDNA3-HA阴性对照质粒加入到23 μL OPTI-DMEM培养基,混匀后再加入1 μL Lipofectamine 2000脂质体转染试剂,室温环境中孵育10 min。将150 μL含10% FBS的1640培养液与质粒-脂质体混合物混匀后加入Petri皿中,转染完后将细胞置于5% CO2、37 ℃培养箱中培养。
1.2.5 细胞免疫荧光检测 HepG2细胞转染pcDNA3-p54nrb-HA质粒24 h后进行细胞免疫荧光实验。用PBS洗涤细胞1次;用4 %多聚甲醛固定细胞15 min,用PBS洗涤细胞1次;向细胞加入含有0.1 % Triton X-100的PBS,室温下孵育5 min,用PBS漂洗细胞3次;1‰ 硼氢化钠孵育5 min;用含3 % BSA的PBS室温封闭细胞1 h,然后再用0.1 % Triton X-100/ PBS洗涤细胞1次;加入鼠源性抗HA一抗抗体(稀释比为1:100),4 ℃孵育过夜,0.1% Triton X-100/PBS 漂洗3次;加入Alexa-Flour 488 标记的羊抗鼠IgG荧光二抗,稀释比为1:1 000),室温避光孵育1 h,漂洗3 次; DAPI/PBS核染色10 min,洗涤细胞2 次,然后用PBS封片并在荧光显微镜下观察及照相。
2 结果
2.1 pcDNA3-p54nrb-HA载体的构建及鉴定
通过RT-PCR扩增p54nrb基因的蛋白编码区后,对扩增产物进行凝胶电泳,可见在1 400 bp左右处出现清晰条带,与1 461 bp p54nrb目的片段大小相符。对质粒pcDNA3- p54nrb-HA进行双酶及回收酶切产物凝胶电泳后,可见约1.4 kb和5.4 kb大小条带(图1)。pcDNA3-p54nrb-HA质粒测序结果与GenBank所提供的人p54nrb蛋白编码区基因cDNA 序列(GenBank: CR456761.1)一致(图2),说明该质粒构建成功。
2.2 细胞免疫荧光检测p54nrb在细胞内表达和定位
转染pcDNA3-p54nrb-HA质粒后,静息状态下绿色荧光只分布于细胞核,如图3所示。
3 讨论

图1 p54nrb蛋白编码区的PCR扩增及pcDNA3-p54nrb-HA质粒的酶切鉴定
NONO/p54nrb属于果蝇行为/人类剪接蛋白(drosophila behavior/human splicing,DBHS)家族中的一员。DBHS蛋白几乎参与基因调控的每一个步骤,包括转录调控,RNA加工和运输,以及DNA修复等。现在研究表明,DBHS蛋白是动态蛋白质家族,它介导了广泛的蛋白质-蛋白质和蛋白质-核酸相互作用,总体上可作为一个多用途的分子支架,该家族在细胞生物学中作用复杂而且重要[15]。p54nrb蛋白可以单独或与其他转录因子协同作用,与双链或单链DNA以及单链RNA作用,参与基因转录的激活或抑制,转录的起始、延伸或终止,以及转录后的加工和输出等多个步骤[1],如p54nrb/NONO与视紫红质增强子区结合,增强视紫红质启动子转录活性,促进视紫红质的表达[16]。p54nrb作为孕激素受体(Progesterone Receptor,PR)转录阻遏物抑制PR介导的孕激素反应元件的反式激活,引起下游RNA的降解可以促进转录的终止[6]。p54nrb基因敲除削弱了ERK激活,进而抑制RNA聚合酶在小鼠胚胎干细胞双线期基因靶点的定位,扰乱了靶基因的激活。NONO/p54nrb参与双链DNA的解螺旋,p54nrb与剪接因子PSF与形成异源二聚体,结合DNA拓扑异构酶I,增强DNA拓扑异构酶I活性。p54nrb还参与超编辑RNA核滞留和DNA损伤修复等。
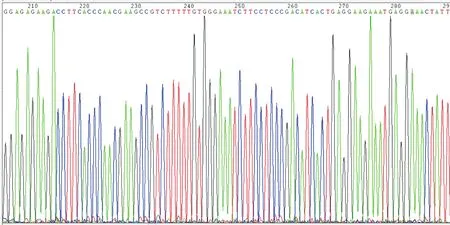
图2 pcDNA3-p54nrb-HA质粒的测序结果

图3 p54nrb在HepG2细胞的表达与定位(200×)
由于p54nrb参与了基因的复制、转录和转录后mRNA的剪切等多个过程,p54nrb基因突变或异常可引起广泛的基因mRNA谱表达异常,其表达或功能异常与多种生理以及病理疾病有关。有研究表明p54nrb基因可能参与胶原蛋白代谢过程。NONO 作为昼夜节律蛋白PERIOD(PER)的分子伴侣参与衰老和细胞周期的调控。NONO对于局部树突的形态和桥接蛋白的结构有重要作用;p54nrb/NONO可能是一个影响神经发育的重要基因,该基因突变或缺失可能与人智力低下的疾病有关,p54nrb缺陷小鼠有认知和情感缺陷[11]。现研究表明,NONO参与多种肿瘤的发生和发展,如p54nrb与乳腺癌、前列腺癌、结直肠癌、膀胱移行细胞癌、恶性黑色素瘤、非小细胞肺癌、红白血病等多种肿瘤的发生和发展密切相关[12-14]。p54nrb功能的多样性,促使学者们对其功能进行深入研究,本实验成功地克隆了人p54nrb基因真核表达载体,并通过转染实验证实p54nrb分布于细胞核。该载体的构建,将为后续进一步研究p54nrb功能提供重要工具。
猜你喜欢
特产研究(2024年1期)2024-03-12 05:40:56
天然产物研究与开发(2019年10期)2019-11-05 10:12:44
食品科学(2018年10期)2018-05-23 01:27:28
中成药(2017年9期)2017-12-19 13:34:27
CHINESE JOURNAL OF AERONAUTICS(2017年1期)2017-11-21 12:54:14
中成药(2017年5期)2017-06-13 13:01:12
西南医科大学学报(2015年1期)2015-08-22 13:01:46
中国当代医药(2015年9期)2015-03-01 02:01:59
西南军医(2015年6期)2015-01-23 01:25:50
河北医科大学学报(2011年1期)2011-03-25 10:15:30
