
Wnt/β-catenin信号通路促进小鼠胚胎干细胞向胰岛素分泌细胞分化
2020-01-01 09:07何巧娟张娜娜殷应传胡红琳
安徽医科大学学报 2019年12期
何巧娟,张娜娜,殷应传,胡红琳
糖尿病的患病率在我国逐渐升高,胰岛细胞功能破坏所导致的内源性胰岛素分泌不足是发生糖尿病的重要原因,但是外源性胰岛素由于需要多次注射且存在导致低血糖等风险而在临床使用时受到限制,迫切要求寻找新的治疗途径[1]。胚胎干细胞(embryonic stem cells,ESCs)可以无限地进行自我更新,保证了临床治疗时对细胞数量的要求[2]。同时通过体外定向诱导分化,ESCs还能够成功分化为胰岛素分泌细胞,将其移植到患糖尿病小鼠体内,能够降低血糖浓度,为糖尿病的细胞替代治疗提供了新的方向[3-5]。但是目前胰岛细胞分化还存在很多问题,如效率低、功能低下等。胰岛细胞源于内胚层细胞,Wnt/β-catenin信号通路在内胚层的定向分化过程中发挥着重要功能[6]。该研究将在小鼠ESCs向胰岛素分泌细胞分化时激活Wnt/β-catenin信号,检测调控此信号通路能否提高胰岛细胞的分化效率。研究结果将为未来利用干细胞治疗糖尿病提供理论基础和策略。
1 材料与方法
1.1 试剂与仪器E14小鼠胚胎干细胞(mouse ESCs,mESCs)和293T细胞购自中国科学院细胞库;细胞培养基和血清购买自以色列Biological Industries公司;Pfu Polymerase和荧光定量PCR(qPCR)试剂盒购买自北京天根生化有限公司;GAPDH、β-catenin、Pdx1和Insulin1 (Ins1) 等抗体购自美国ProteinTech和英国Abcam公司;BamHⅠ、XhoⅠ限制性内切酶和T4连接酶购自日本TAKARA公司; 琼脂糖凝胶DNA回收试剂盒、质粒抽提试剂盒购买自美国Axygen公司;白血病抑制因子购自美国Millipore公司;细胞因子Activin A、bFGF和FGF10等细胞因子均购自美国Peprotech公司;ITS、B27、Lipofectimine 3000脂质体和Puromycin等购买自美国Invitrogen公司;KAAD、Tetrabenazine和尼克酰胺等胰岛细胞分化所用的试剂购自美国Sigma公司;核酸分析仪Nanodrop、普通和定量PCR仪、生物安全柜和细胞培养箱等均购自美国Thermo Scientific公司;蛋白与核酸电泳设备购自北京六一生物科技有限公司。
1.2 方法
1.2.1mESCs的培养与传代 培养皿用0.1%的明胶进行包被,加入mESCs及血清培养基。培养基的组成为:DMEM、10%胎牛血清;1×非必需氨基酸、0.1 mmol/L β-巯基乙醇以及1 000 U/ml LIF。当细胞生长密度达70%~80%时,用胰酶消化成单个细胞进行传代培养。
1.2.2mESCs向胰岛细胞分化 该实验主要基于前期研究人员发明的三步法[3]诱导mESCs向胰岛细胞分化。5 000个mESCs接种在96个孔细胞培养板中,依次经过3个不同配方的溶液进行处理,17 d后收集细胞进行分析。溶液1的配方:DMEM,4 500 mg/L葡萄糖,1×非必需氨基酸,2 mmol/L L-谷氨酰胺,1 mmol/L丙酮酸, 100 μmol/L β-巯基乙醇,ITS,0.25% Albmax,10 ng/ml Activin A和5 ng/ml bFGF。溶液2的配方:RPMI 1640,2 000 mg/L葡萄糖,1 μmol/L retinoic acid,50 ng/ml FGF10,2%B27和0.25 μmol/L Shh信号激动剂KAAD。溶液3的配方:DMEM,1 000 mg/L葡萄糖,L-谷氨酰胺,β-巯基乙醇,ITS,0.25% Albmax,10 nmol/L GLP-1,Tetrabenazine和10 mmol/L尼克酰胺。
1.2.3β-catenin基因重组质粒的构建、转染和筛选 针对克隆小鼠β-catenin基因编码区设计上下游引物,引物序列为Forward:5′-ATGGCTACTCAAG CTGACCTGATGG-3′; Reverse:5′-TTACAGGTCAGTATCAAACCAGGCC-3′。在上下游引物两端分别添加BamHⅠ和XhoⅠ的限制性内切酶位点,并连入PiggyBac(PB)-TET On系统。利用脂质体转染进mESCs,2 d后加入终浓度为2 μg/ml的Puromycin筛选阳性克隆。
1.2.4RNA干扰慢病毒系统的构建与感染 针对小鼠β-catenin基因的mRNA序列,利用BLOCK-It RNAi Designer设计2个21 bp的小发卡RNA(short hairpin RNA,shRNA),分别命名为:shRNA#1(sh#1)和shRNA#2(sh#2),连入pLKO.1慢病毒载体,干扰序列分别为sh#1:5′-GGACCTACACTTA TGAGAAGC-3′ 和sh#2: 5′-GCACCATGCAGAATACAAATG-3′。pLKO.1载体、辅助质粒psPAX2和VSVG按4:2:1的质量比混合,利用脂质体将其转染到293T细胞中,2 d后收集慢病毒感染mESCs。48 h后,加入终浓度为2 μg/ml的Puromycin进行筛选。
1.2.5荧光实时定量PCR(qPCR) 用北京天根生化有限公司的RNA提取试剂盒抽提细胞总RNA,利用FastKing预混试剂进行第一链cDNA的合成,接着按照Talent荧光定量检测试剂盒的说明进行qPCR。所用的基因序列如下。β-actin-Forward:5′-TGTTACCAACTGGGACGACA-3′;β-actin-Reverse:5′-GGGGTGTTGAAGGTCTCAAA-3′;Pdx1-Forward: 5′-CCAAAACCGTCGCATGAAGTG-3′;Pdx1-Reverse:5′-CTCTCGTGCCCTCAAGAATTTTC-3′;Ngn3-Forward:5′-CTGCGCATAGCGGACCACAGCTTC-3′;Ngn3-Reverse:5′-CTTCACAAGAAGTCTGAGAACACCAG-3′;Pax4-Forward:5′-CCACCTCTCTGCCTGAAGAC-3′;Pax4-Reverse:5′-CCCACAGCATAGCTGACAGA-3′;Ins1-Forward:5′-CAGCCCTTAGTGACCAGCTA-3′;Ins1-Reverse:5′-ATGCTGGTGCAGCACTGATC-3′。
1.2.6Western blot检测 收集各组细胞,利用添加了蛋白酶抑制剂PMSF的RIPA裂解液裂解细胞,BCA法测定总蛋白浓度。用8%的SDS-PAGE胶分离各蛋白组分,并利用湿转系统将蛋白转移至PVDF 膜上,经过封闭、一抗孵育、TBST洗脱、HRP标记的二抗孵育、TBST再洗脱等步骤后,使用ECL发光液进行发光。使用Image J进行灰度值分析比较,实验重复3次。
1.2.7免疫荧光 吸去培养液,加入4%多聚甲醛室温固定20 min,PBS洗两次,加入含有5% BSA的PBS封闭液在37 ℃温箱孵育1 h,0.2% Triton X-100溶液进行细胞膜打孔,加入含有一抗的稀释液(1 ∶100)覆盖细胞,4 ℃过夜,次日用PBS漂洗3次,加入含有绿色荧光基团488的二抗以及Hoechst染料的稀释液(稀释比例均为1 ∶5 000),37 ℃避光孵育1 h,用PBS漂洗后在荧光显微镜下进行观察拍照。
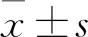
2 结果
2.1 胚胎干细胞向胰岛素分泌细胞定向分化mESCs向胰岛细胞分化主要分为3个阶段,见图1A。第一阶段为1~7 d,主要通过添加“溶液1”进行诱导;第二阶段为7~11 d,细胞培养液换成“溶液2”;最后阶段为11~17 d,细胞培养在“溶液3”中。每2 d换1次液。17 d后收集细胞检测胰岛素分泌细胞形成的情况。qPCR结果显示:随分化时间的增加,胰岛前体细胞标志基因Pdx1(pancreatic and duodenal homeobox factor 1),Ngn3(neurogenin 3)和Pax4 (paired-homeobox 4)的表达逐渐增加。见图1B。至17 d时Ngn3和Pax4的表达有所降低,但是胰岛素基因Ins1的表达达到最高值(62.110±12.457,F=298.4,P<0.01),差异有统计学意义,说明有大量的胰岛素分泌细胞生成。见图1B。本研究分别使用了免疫荧光和Western blot进行了检测,Pdx1与Ins1的蛋白在第17天时高水平表达,与qPCR检测结果一致,见图1C和1D。表明mESCs被成功地诱导分化成了胰岛素分泌细胞。
2.2 CHIR99021促进胰岛素分泌细胞的分化形成为了检测Wnt/β-catenin信号通路在胰岛细胞形成过程中的功能,本研究在mESCs向胰岛细胞分化的三个不同阶段分别添加3 μmol/L CHIR,然后检测胰岛素分泌细胞的形成情况。见图2A。结果显示在分化的第一阶段,即第1~7天,CHIR刺激后,第7天时,胰岛β细胞的标志基因Pdx1和胰岛素Ins1表达均上调(F=3.745、3.987,P<0.05),差异有统计学意义。同样在第三阶段,即第11~17天,CHIR刺激后,第17天时,胰岛β细胞的标志基因Pdx1和胰岛素Ins1表达均上调(F=9.770,P<0.01;F=5.819,P<0.05),差异有统计学意义。且相比于第一阶段,第三阶段第17天收集的细胞经CHIR处理后产生更多的Ins1,qPCR检测的结果分别为(82.121±15.422)和(99.218±19.803),见图2B、2D。而在第二阶段添加CHIR并不影响Pdx1和Ins1的表达,差异无统计学意义,见图2C。以上结果表明,在mESCs向胰岛细胞分化的早期和晚期,添加CHIR能够增加胰岛细胞的定向分化效率。
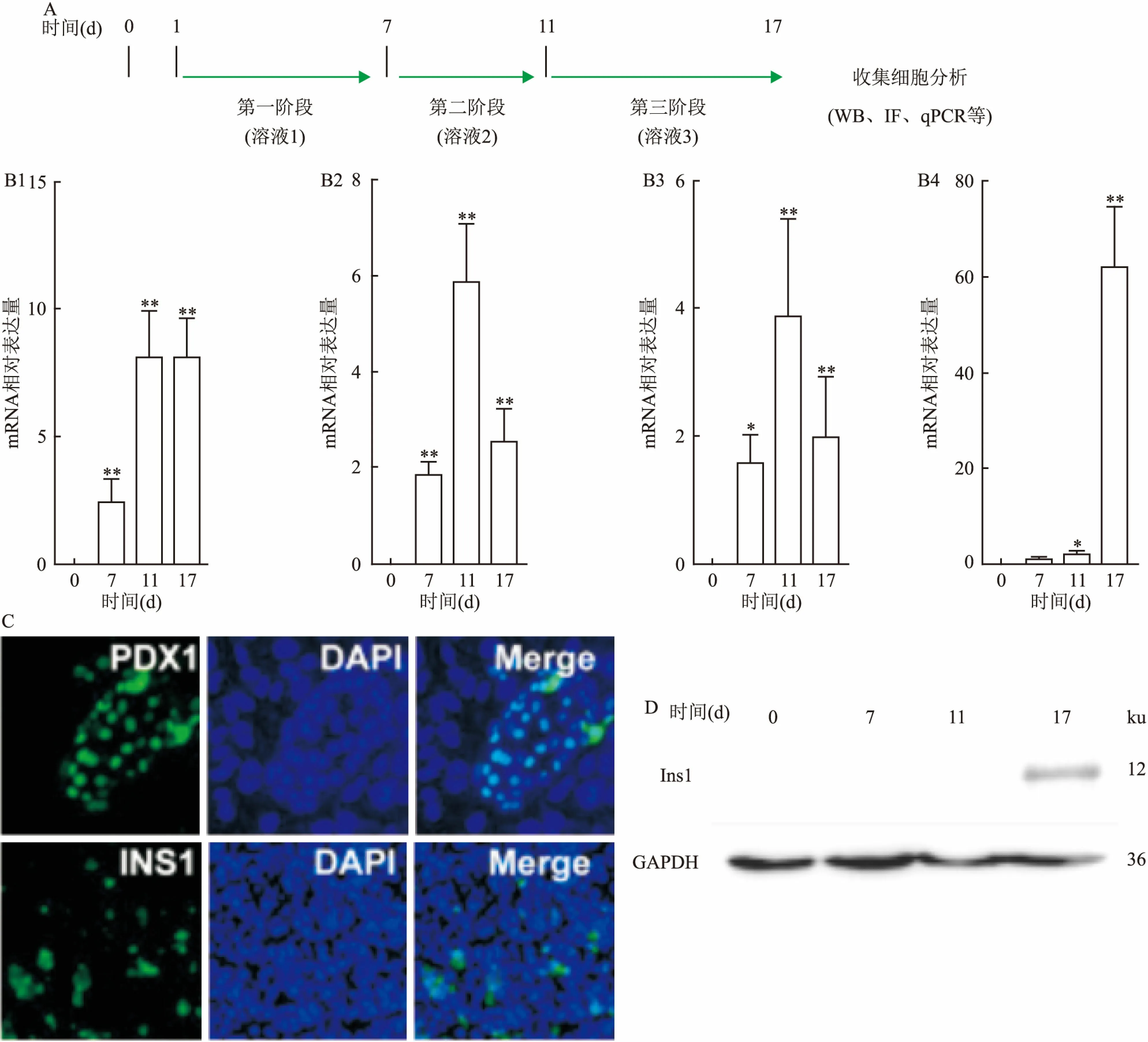
图1 mESCs分化为胰岛细胞
A:分化方案;B:qPCR检测Pdx1、Ngn3、Pax4和Ins1的表达;1:Pdx1;2:Ngn3;3:Pax4; 4:Ins1;与0 d比较:*P<0.05,**P<0.01;C:免疫荧光检测Pdx1和Ins1表达 ×100;D:Western blot检测Ins1和GAPDH的表达;GAPDH为内参蛋白
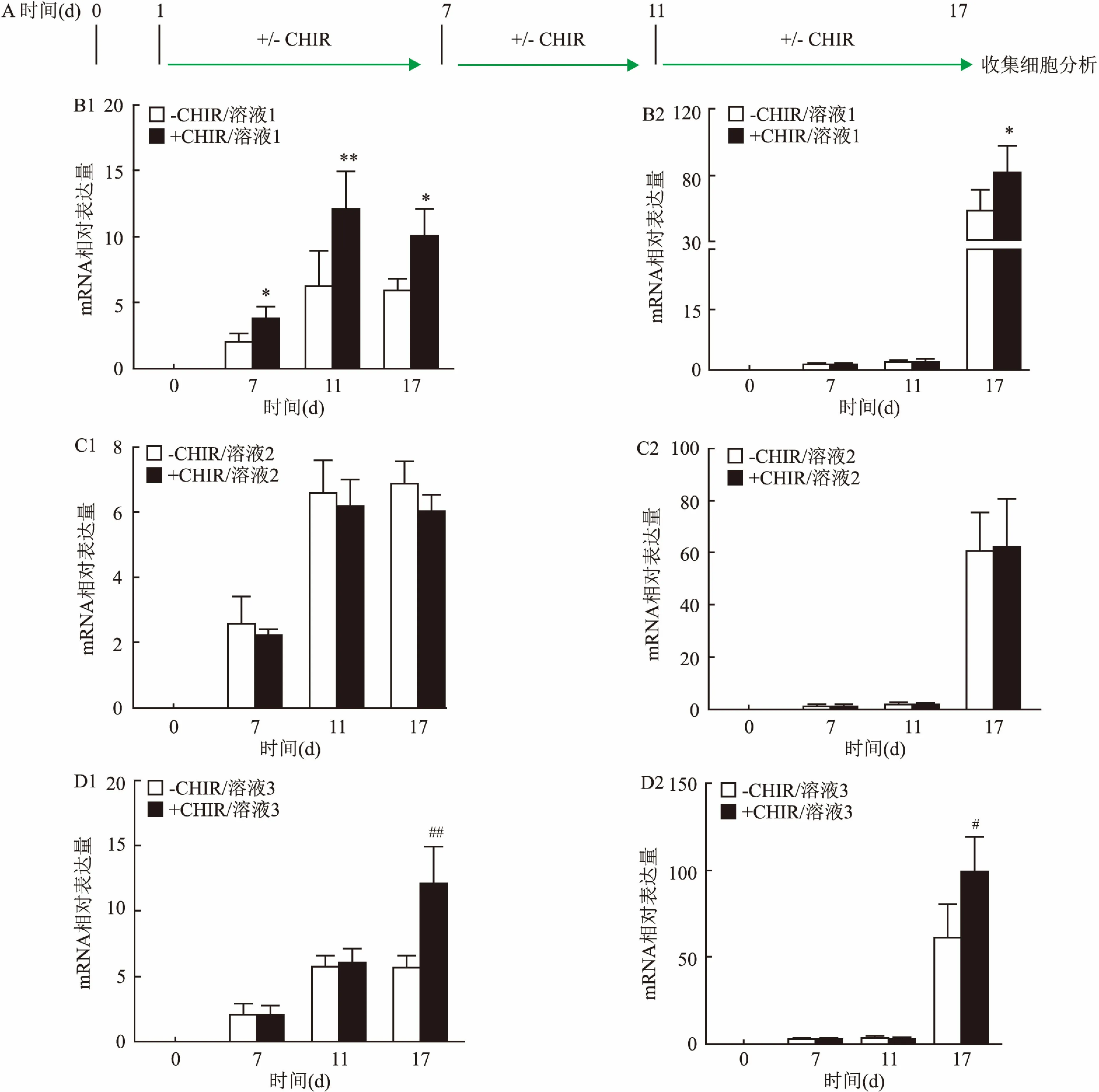
图2 CHIR在胰岛细胞定向分化中的作用
A:实验方案;B:溶液1中有无CHIR时,qPCR检测Pdx1和Ins1的表达;C:溶液2中有无CHIR时,qPCR检测Pdx1和Ins1的表达;D:溶液3中有无CHIR时,qPCR检测Pdx1和Ins1的表达;1:Pdx1;2:Ins1;与-CHIR/溶液1组比较:*P<0.05,**P<0.01;与-CHIR/溶液3组比较:#P<0.05,##P<0.01
2.3 诱导β-catenin基因表达能够促进胰岛细胞的诱导分化为了明确Wnt/β-catenin信号通路在mESCs向胰岛细胞分化过程中发挥功能,本研究首先使用Tet-on质粒构建了一个β-catenin可诱导表达系统,并导入mESCs里。Western blot结果以及蛋白条带灰度分析结果表明,添加终浓度为2 μg/ml的盐酸强力霉素(Doxcycline,Dox)可以有效地诱导β-catenin基因的表达(F=279.9,P<0.01)。见图3A、3B。接着,根据CHIR的实验结果,在mESCs向胰岛素分泌细胞分化的第一和第三阶段分别添加Dox,然后检测胰岛细胞的形成情况。qPCR结果显示:在第一阶段,与未处理组相比,Dox处理的细胞在第17天时表达更多的Pdx1和Ins1 (F=11.2、92.49,P<0.01),差异具有统计学意义,见图3C;同样,在第三阶段,第17天收集的细胞中,Dox诱导更高的Pdx1和Ins1表达(F=39.84、147.4,P<0.01),差异有统计学意义,见图3D。且溶液3和Dox诱导细胞产生的Ins1高于溶液1和Dox的功能,分别为(99.113±18.191)和(117.452±21.233),见图3C、3D。以上研究结果说明在胰岛细胞定向分化的早期和晚期阶段,激活Wnt/β-catenin信号可以促进mESCs向胰岛细胞的定向分化。
2.4 干扰β-catenin基因表达削弱CHIR在胰岛细胞定向分化中的功能为了反向验证β-catenin是否可以介导CHIR的作用,本研究设计了两条针对β-catenin基因的靶向干扰序列,包装慢病毒并感染mESCs,药杀筛选后,在转录水平用qPCR检测,结果显示:相比较于Scramble对照组(1.000±0.333),在β-catenin shRNA两株细胞内,β-catenin的相对表达分别为(0.201±0.111)和(0.186±0.103),差异有统计学意义(P<0.01)。见图4A。同样,Western blot结果显示:相比较于对照组Scramble病毒感染的细胞,β-catenin shRNA病毒感染的细胞中β-catenin的蛋白水平被有效地下调。见图4B。Scramble和β-catenin shRNA细胞培养在含CHIR的培养条件下,17 d后,收集细胞进行检测。qPCR结果显示,在胰岛细胞分化的第一阶段,与Scramble对照组相比,干扰β-catenin基因表达能够减弱CHIR的功能,所以胰岛细胞形成的效率低于Scramble病毒感染的细胞,差异有统计学意义(F=13.92、31.50,P<0.01)。见图4C。同样,在第三阶段干扰β-catenin表达后,第17天收集细胞进行检测,也显示干扰β-catenin表达也会减弱CHIR的功能(F=9.036、11.27,P<0.01),差异有统计学意义。说明CHIR主要通过激活Wnt/β-catenin信号通路从而诱导mESCs向胰岛素分泌细胞,而干扰β-catenin基因则会抑制CHIR的作用。
3 讨论
糖尿病一直没有特效的治疗方法,寻找替代材料和疗法现已成为临床上亟待解决的重大问题之一。本项目研究建立并优化了目前胚胎干细胞向胰岛素分泌细胞的分化方法,表明在胚胎干细胞向胰岛细胞分化的早期和晚期,用小分子CHIR处理可以有效地诱导胰岛细胞的生成效率。因为GSK-3抑制很多蛋白的活性,从而调控多个信号通路[7],所以接下来通过调控β-catenin基因的表达来确定CHIR确实通过Wnt/β-catenin信号发挥功能。结果表明过表达β-catenin能够模拟CHIR的作用,从而促进小鼠胚胎干细胞向胰岛细胞分化,而下调β-catenin基因的表达则会削弱CHIR的功能。因此,激活Wnt/β-catenin信号通路对于高质量胰岛素分泌细胞的高效产生具有促进作用。
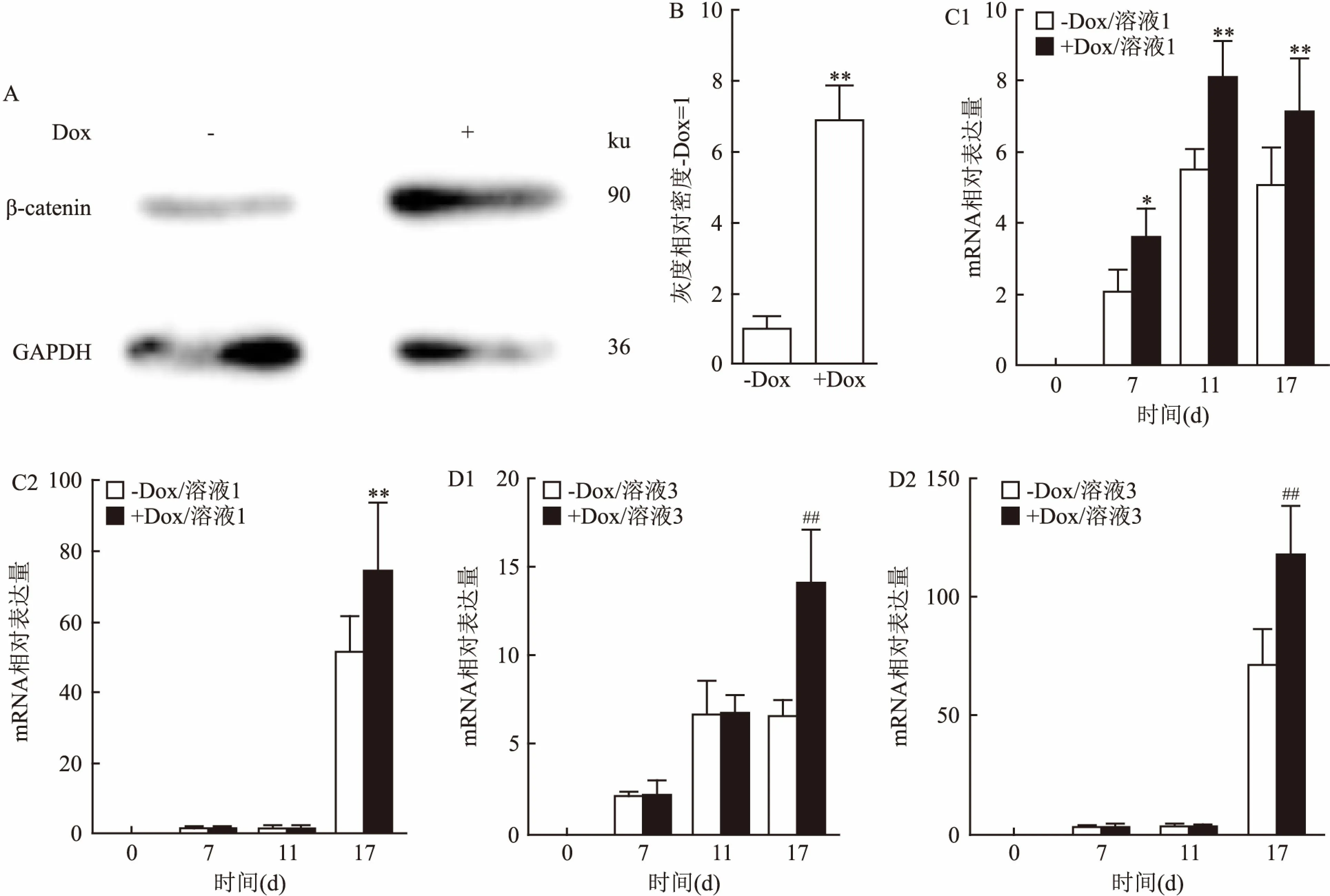
图3 β-catenin对胰岛细胞分化形成的影响
A:有无 Dox条件下,Western blot检测β-catenin和GAPDH的表达;B:根据A的结果进行条带灰度分析;C:溶液1中有无Dox时,qPCR检测Pdx1和Ins1的表达;D:溶液3中有无Dox时,qPCR检测Pdx1和Ins1的表达;1:Pdx1;2:Ins1;与-Dox/溶液1组比较:*P<0.05,**P<0.01;与-Dox/溶液3组比较:##P<0.01
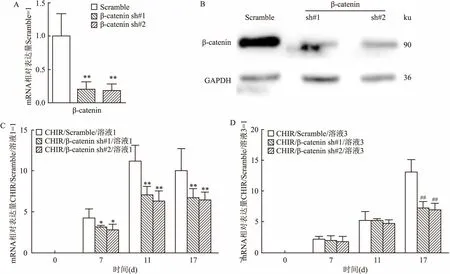
图4 β-catenin介导CHIR诱导mESCs向胰岛细胞分化的作用
A:qPCR检测β-catenin的表达;B:Western blot检测β-catenin和GAPDH的表达;C:溶液1中添加CHIR后,在有无干扰β-catenin基因表达时,qPCR检测Pdx1的表达;D:溶液3中添加CHIR后,在有无干扰β-catenin基因表达时,qPCR检测Pdx1的表达;与CHIR/Scramble/溶液1组比较:*P<0.05,**P<0.01;与CHIR/Scramble/溶液3组比较:##P<0.01
目前将胚胎干细胞诱导分化为胰岛素分泌细胞有多种手段,如:① 通过过表达胰腺组织特异性的转录因子Pdx1、Ngn3、Pax4、Nkx2.2和Nkx6.1等[8];② 通过组合不同生长因子进行诱导,如Activin A、Tgf-β、VEGF和bFGF等[9];③ 通过将胚胎干细胞形成拟胚体进行自发分化[10]。但是这些方法胰岛素分泌细胞分化的效率比较低,远不能满足糖尿病移植需要,而且降血糖的效果也不是十分理想。Wnt/β-catenin信号通路在胚胎早期发育过程中,对于中内胚层形成极其重要,同样在胚层干细胞中,激活此信号通路也能够诱导干细胞向中内胚层细胞的分化[6]。虽然前期也有报道:激活此信号通路能够促进胚胎干细胞向胰岛样细胞分化,但是得到的是胰岛前体细胞[11]。Sakano et al[3]通过使用1 100个小分子化合物处理胰岛前体细胞,发现vesicular monoamine transporter 2(VMAT2) 的抑制剂Reserpine和Tetrabenazine可以增加胰岛细胞分泌胰岛素,从而发明了一种胚胎干细胞分化成成熟胰岛β细胞的方法,培养出的胰岛β细胞媲美正常实验小鼠内的胰腺细胞[3]。但是在这个方案中他们未尝试调控Wnt/β-catenin信号通路。基于此方法,本研究在胚胎干细胞向胰岛细胞分化的早期和晚期激活此信号能够促进胰岛细胞的产生,见图2、3,不仅体现在分化效率上,并且也能够诱导更加成熟的胰岛细胞形成,因为本研究优化的方案得到的胰岛细胞能够分泌更多的胰岛素。小分子CHIR是GSK3的选择性抑制剂,能够有效地激活Wnt/β-catenin信号通路,现已被广泛用于干细胞的培养和扩增[12],所以它的运用可以避免使用动物源性来源的Wnt蛋白,有利于未来的临床治疗应用。
间充质干细胞具有免疫原性低和多能性等优点,尽管目前已有报道被应用于临床上治疗糖尿病,有能够有效地降低血糖、修复胰岛细胞、缓解晚期患者的糖尿病足等[13]。但是间充质干细胞的治疗效果因人而异,而且其直接进入人体存在很多不确定性,如分化的效率、分化形成的细胞类型、在体内存活的时间、归巢的位置以及发挥的其他功能等,增加了其应用的潜在风险性[14]。而应用胚胎干细胞分化而来的胰岛素分泌细胞能够有效地解决以上问题。加之当前诱导多能性干细胞的出现,患者可以定制自己的“胚胎干细胞”,因为诱导性多能干细胞的生物学特性与胚胎干细胞类似[15],未来可以分离患者自身的成体细胞进行重编程而获得,进而分化为胰岛细胞,为实现糖尿病的个性化治疗提供新的策略。
猜你喜欢
医学信息(2022年9期)2022-11-27
今日农业(2022年13期)2022-09-15
中国生殖健康(2020年5期)2021-01-18
生物学通报(2020年10期)2020-08-13
中国生殖健康(2018年5期)2018-11-06
知识经济·中国直销(2017年10期)2017-11-07
新疆医科大学学报(2015年10期)2015-12-26
中国病理生理杂志(2015年8期)2015-12-21
中国卫生(2014年11期)2014-11-12
中国卫生(2014年2期)2014-11-12
