
海洋浮游介形类的多样性和生态学研究进展
2019-12-24 06:36廖彤晨尹健强李开枝谭烨辉
生态科学 2019年6期
廖彤晨, 尹健强, 李开枝,3,4*, 谭烨辉,3,4
海洋浮游介形类的多样性和生态学研究进展
廖彤晨1,2, 尹健强1, 李开枝1,3,4*, 谭烨辉1,2,3,4
1. 中国科学院热带海洋生物资源与生态重点实验室, 中国科学院南海海洋研究所, 广州 510301 2. 中国科学院大学, 北京 100049 3. 广东省应用海洋生物学重点实验室, 中国科学院南海海洋研究所, 广州 510301 4. 中国科学院南海生态环境工程创新研究院, 广州 510301
海洋浮游介形类(ostracods)是一类分布较广的小型甲壳动物, 大多数是典型的有机碎屑摄食者, 本身又是中深层鱼类的饵料, 在海洋碳循环中起重要作用。目前海洋浮游介形类研究主要集中在海洋表层, 大约还有200—400 种的海洋浮游介形类未被发现, 尤其是栖息于深海的种类。今后介形类分类学研究的重要趋势之一是深海种类的发现。我国介形类研究起步晚, 积累少, 应加强我国不同海域深海介形类及其与环境关系的深入研究。文章综述了海洋浮游介形类的多样性和生态学等方面的研究进展, 重点概述了海腺萤科分类的发展, 比较分析了不同海域介形类的物种多样性和分布特征, 总结了浮游介形类的生态作用及其影响因素, 概括了我国海洋浮游介形类的研究现状, 以期为海洋生物多样性、海洋生物地球化学循环以及深海生态学等方面的研究提供基础依据。
多样性; 动物地理学; 生态学; 海腺萤科; 介形纲
0 前言
介形类(ostracods)隶属于节肢动物门(Arthropoda)、甲壳动物亚门(Crustacea)、介形纲(Ostracoda)。介形类是一类小型甲壳动物, 从古生到今生、陆地到水体皆有分布。介形纲在海洋浮游生物学研究中常称作介形类。海洋浮游介形类体长范围在0.5—3 mm, 深海的大海萤()可达30 mm, 以海腺萤科(Halocyprididae)为主, 其在热带—亚热带海域的浮游动物中数量仅次于桡足类, 喜栖息于海洋中层(mesopelagic zone)和深层(bathypelagic zone), 昼夜垂直移动明显。全球海洋生物普查计划发现中深层鱼类资源产量是全球经济鱼类产量的2—3 倍, 而深海浮游动物如介形类和胶质类生物是其主要食物来源[1]。海洋浮游介形类是典型的有机碎屑摄食者, 摄食从海洋表层沉降到深海的“海雪”及其它有机碎屑, 而其本身又是中深层鱼类的饵料, 在海洋表层至底层的食物链和碳循环中充当“生物泵”作用[1-3]。海洋介形类因自身特性对水温和盐度变化敏感, 某些种类只分布于某些适应的海域, 可作为水团水系变化的指示种[4]。介形类在生物繁殖方式的进化、生态系统结构与功能和气候变化等研究中是非常热门的模式生物[5-6]。另外, 海腺萤介形类的一些种类能进行生物发光, 是研究生物发光的良好材料[7]。
由于海洋浮游介形类的种类个体小, 鉴定困难, 以及多栖息于海洋中、深层分布等原因, 其分类学和生态学研究至今仍未被充分重视。本文对海洋浮游介形类的分类学和生态学研究现状进行了综述, 以期为海洋生物多样性和海洋生物地球化学循环等研究提供基础依据。
1 海洋浮游介形类的多样性研究
1.1 海洋介形类的分类研究概况
海腺萤科种类是浮游介形类的主要组成[8], 现包括5 亚科54 属223 种[9], 见图1, 数据统计来源于World Register of Marine Specis网站(WoRMS), 截至2018 年11 月。终生营浮游生活的海洋介形类, 多数属于海腺萤科(Halocyprididae), 少数属于海萤科(Cypridinidae); 前者主要分布在深海, 后者主要生活在近海[8]。海腺萤科隶属于介形纲、壮肢亚纲(Myodocopa)、海腺萤目(Halocyprida)、海腺萤亚目(Halocypridina)。全球海洋浮游介形类仍约有200—400 种未被发现[10], 其中主要是一些栖息于特殊生境如深海的海腺萤科种类[8]。
海洋浮游介形类的身体完全被双瓣的甲壳(carapace)所覆盖, 甲壳一般不钙化, 成体具7 对附肢(limbs)和1 对尾叉(caudal furca)。甲壳和附肢的形态及其结构是重要的分类特征, 种类确定主要依据雌、雄成体的形态特征。海腺萤科种类根据分类学中的形态特征, 比如甲壳图案模式、甲壳腺体的位置和第一、第二触角的刚毛, 以及海腺萤科的自身特点, 被分为五个亚科: 原萤亚科(Archiconchoeciinae)、深海浮萤亚科(Bathyconchoeciinae)、浮萤亚科(Conchoeciinae)、真浮萤亚科(Euconchoeciinae)和海腺萤亚科(Halocypridinae)。发现并描述最多的是浮萤亚科, 共有37 属137 种[9]。
海腺萤科分类始于1849 年, Dana描述了第一种。Sars、Claus和Müller等学者对海腺萤科进行了大量开创性和奠基性的描述工作(图2, 数据统计来源于WoRMS, 截至2018 年11 月)。20 世纪60 年代起, Deevey、Gooday、Chavtur和Angel等学者[11-12]对一些容易混淆的种类进行了澄清并陆续发现了一些新种, 特别是深海浮萤亚科的种类(图2)[13], 以上标本主要来源于挪威附近海域、马尾藻海、墨西哥湾和加勒比海等大西洋海域。Chavtur[14-16]对北太平洋的海腺萤分类区系进行了研究, 并发现了一些新种[17]; Graves[18-19]在印度洋阿曼湾发现了海腺萤科1 新属3 新种。19 世纪末20 世纪初, 英国学者Angel和俄罗斯学者Chavtur分别对大西洋和北太平洋的1000 m上层介形类进行了较深入的研究, 介形类多样性在这两个海域被研究得较为透彻。北冰洋、靠近南极洲和澳大利亚的太平洋海域、印度洋北部的孟加拉湾和阿拉伯海也有被研究, 而在太平洋中部及印度洋中部、南部深层海域的研究则相对缺乏。总体而言, 大西洋是海腺萤科介形类研究最全面的区域, 而在太平洋和印度洋的分类学研究则相对薄弱。
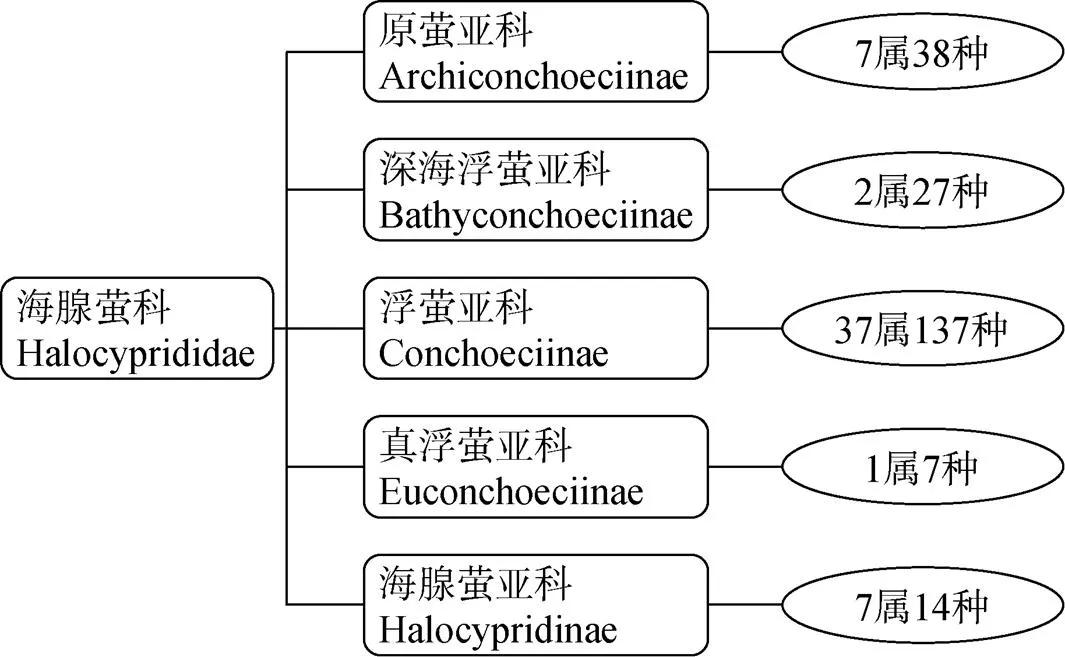
图1 海腺萤科的亚科、属和种数(数据统计来源于WoRMS网站, 截至2018 年11 月)[9]
Figure 1 Taxon of Halocyprididae (collected from WoRMS by November 2018)[9]

图2 海腺萤科分类研究历史简图(数据统计来源于WoRMS网站, 截至2018年11月)[9]
Figure 2 The history of taxonomy of Halocyprididae (collected from WoRMS by November 2018)[9]
Poulsen[20-21]对全球海腺萤的分类和分布进行了研究, 创建了海腺萤科新的分类系统。以甲壳、第一触角(first antenna)和第二触角(second antenna)的形态与结构为主要依据的分类系统逐渐发展到大颚(mandible)和其它附肢也成为建立新分类阶元的重要依据。近十几年一些学者陆续在Poulsen的分类系统基础上进行了修订, 如海腺萤科由4 亚科增加至5 亚科(图1), 原萤亚科由1 属增加至7 属[22], 浮萤亚科由17 属增加至37 属[13], 其中浮萤亚科后浮萤属()由1 属拆分成10 属等[23]。总之, 根据发现的新物种或新标本材料, 海腺萤科的分类系统在原有分类阶元的基础上不断地进行调整。
当前, 海洋浮游介形类分类学研究的重要趋势是继续重视新种的发现, 特别是深海环境的种类。自从Deevey[24]建立深海浮萤属()并发现6 新种以来, 已共有22 种被描述。然而, 许多被发现的深海新种属于其他属, 并有更多深海种类有待发现。深海浮游介形类具有较高的物种多样性, Angel在检查南大洋(Southern Ocean)和大西洋大于1000 m的浮游动物拖网样品中发现, 仍有至少20 种以上的新种待描述[11–13]。海洋上层(<200 m)介形种类可以随海流广泛分布, 而深海种类受海洋中、深层环流的影响, 其地理分布范围受限, 会存在一些地方种(endemic species)或未描述种类。
1.2 海洋浮游介形类物种多样性
大西洋出现的浮游介形类种类最多, 高达153 种, 其西北部海域记录118 种, 包括翼萤属、原萤属、浮萤属等海腺萤介形类29 属, 以及壮肢介形类2 属, 并且、刺喙葱萤()、胖海腺萤()和这四种在数量上占绝对优势[2]。在太平洋西北部海域, Chavtur对浮游介形类的原萤亚科进行了系统的分类, 并发现一些新种[22]。Deevey也对邻近南极的南太平洋海域的浮游介形类进行了研究, 描述了48 种的分布和形态特征, 其中数量最多的是浮萤属的种类[25]。在印度洋海域, George鉴定了浮游介形类的32 种, 其中2 种属于海萤科, 其余全都属于海腺萤科[26]。在北印度洋, 阿拉伯海是介形类大量聚集的海域, 优势种是齿形海萤(), 占介形类总数量的75%, 主要分布在印度西部近岸海域[27]。在南大洋的澳大利亚—新西兰海域, 100 m以上水层仅出现6 种介形类[28]。在南极海域, 二腺翼萤()从表层至深层都有分布深海上层水团600—900 m深(水团起源于北大西洋)发现12 种;, 细齿浮萤(),和这些种类只分布于亚南极; 南极深海下层水团600—2500 m 出现了介形类10 种[28]。而北冰洋的浮游介形类非常贫瘠, 仅记录12 种, 其中大部分来源于北大西洋以及完全独立于太平洋的种类, 数量最多的优势种是[29]。
大西洋的海腺萤介形类种数最多[8,30–31], 太平洋次之[16,25, 32-33], 其次是印度洋[26-27]和南大洋[28, 34], 北冰洋最少[29](图3a)。太平洋和印度洋种类丰富, 五个亚科都有发现。海腺萤中记录最多的是浮萤亚科, 而深海浮萤亚科种数较少。

图3 海腺萤科分类和海域分布(文献来源:大西洋: [8]、[30]和[31], 太平洋: [16]、[33]、[42]和[43],印度洋: [34]和[35], 南大洋: [36]和[44], 中国海及邻近海域: [38]和[39])
Figure 3 Taxon and distribution of Halocyprididae (references: the Atlantic Ocean: [8], [30] and [31], the Pacific Ocean: [16], [33], [42] and [43], the Indian Ocean: [34] and [35], the Southern Ocean: [36] and [44], the Arctic Ocean: [37], the China’s Sea and adjacent water: [38] and [39])
海洋浮游介形类个体小、种类众多且复杂, 难以基于微观形态特征进行鉴定。近年有学者开拓了用DNA鉴定等分子学方法对介形类进行分类[35]。DNA条形码鉴定的方法为介形类研究提供了新思路, 结合经典分类学能增加对浮游动物样品进行常规分类分析的多样性、分布和丰度的有效估计[35]。Bucklin等人利用线粒体细胞色素C氧化酶(COI)基因扩增测序鉴定了大西洋的27 种浮游介形类[36]。我国也有学者用类似的方法对南海的16 种海洋浮游介形类进行了鉴定, 与形态学鉴定的结果相吻合[37]。
1.3 生物地理学研究
海洋浮游介形类从海洋上层、中层、下至5000 m的深层、从赤道到两极都有分布。根据其种类出现的规律和分布特点, 介形类常被分为几类, 如暖水种、冷水种和广布种等等。细长真浮萤()、后圆真浮萤()、尖头毛浮萤()、同心假浮萤()、贞女葱萤()等是常见的暖水种, 常分布于热带和亚热带海域[38]。
某些种类因自身特性及对环境的特殊要求, 只分布于某些区域, 属于区域分布种。例如暖水种中的贞女葱萤、尖头毛浮萤等具有适高温高盐特性, 主要分布于三大洋的暖水区[39]。属于典型的冷水种, 在整个北极海域和大西洋的冷水海域中均有分布。由于连接北冰洋和太平洋的白令海峡、北冰洋海域的楚科奇海和太平洋的白令海的深度太浅, 所以这一深海冷水种不会进入北太平洋, 是区域分布种[40]。而在同一海域出现的秀丽双浮萤()属于广布种, 在北冰洋、大西洋都有分布。常见的广布种还有宽短小浮萤()、秀丽双浮萤、兜甲萤()、圆形后浮萤()、刺额葱萤()、条状原萤(), 这些种类在大西洋、太平洋、印度洋和北冰洋都有分布[41]。、等额壮浮萤()、、和是南半球种类, 属于典型的区域布种, 仅见于南半球。其中等额壮浮萤发现于大西洋的0°S—55°S, 大西洋的33°S—68°S以及印度洋的33°S—54°S[25]。
如今, 海洋生物地理学的研究方法已经从单一考虑物种出现与否与海流水团的关系, 逐渐发展到种类个体大小、丰度以及优势种等与海洋物理过程和生物生产力等因素相互联系起来[42–44]。北大西洋中浮游介形类在海洋温跃层以下物种多样性增加, 至700 m物种最丰富, 然后逐渐降低至2000 m水层[30], 生物地理分布规律与Longhurst划分的生物地球化学分区较一致[41]。
2 海洋浮游介形类的生态学研究
2.1 海洋浮游介形类的分布特征
海洋介形类的分布在纬度、经度和深度上都有明显变化。以大西洋东北部海域为例, 垂直分布上, 浮游介形类种类数量随着水深增加而增加, 100 m以下水层开始急剧增加; 水平分布上, 符合赤道向两极递减的趋势, 可见大西洋大多数介形类适合生存于较暖水域[45]。在北大西洋20°W的不同纬度, 海腺萤介形类白天的丰度在50 m以上水层很低, 几乎不出现[45]。夜晚由于昼夜垂直迁移运动, 低纬度海腺萤丰度在25 m以上水层有所增加[45]。水平分布上, 在比18°N高的纬度, 介形类的丰度随着纬度的增加而减少, 在低纬度有较多种类[45]。在10 °N、30°N、32°N和40°N, 物种丰富度在1000 m上方一致随着深度增加而增加, 而在1000 m下方开始减少[45]。
南大洋的澳大利亚—新西兰海域和北冰洋的介形类种数都在1000 m下方开始减少, 种数最大值都出现在200—900 m之间[28-29]。在南极, 种类数量从北到南呈减少趋势[28]。在亚南极海区, 热带—亚热带种类在500 m水层出现, 导致这一水层介形类丰度最大[28]。由于温度降低, 500 m下的种类数开始减少[28]。大部分在夜晚开始于表面聚集, 早上生物量最高值出现在250—500 m处, 早上9 点到下午3 点, 大量介形类出现在250—500 m和500—750 m处[28]。在北印度洋, 介形类数量在近海远比远海高, 阿拉伯海是介形类大量聚集的海域[27]。
不同海域的环境特征不同, 浮游介形类所受的影响有差异, 其分布特征也不同, 因此不同海域的浮游介形类分布规律仍需继续深入研究。
2.2 影响海洋浮游介形类分布的主要因素
浮游介形类的分布由其自身特征对环境的要求决定, 同时也受物理过程的影响, 如温度盐度的变化、含氧量、水团水系的盛衰、季风的转换、太阳辐射和降雨都影响着介形类的分布[45]。
南海海域夏季盛行西南季风, 南海中部西南部海域被爪哇海和巽他陆架的低盐水侵入, 可出现少数低盐暖水种[46]。同时, 南海暖流沿东北方向流动, 导致南海北部出现大量高温高盐种。冬季太阳辐射减弱, 加之东北季风影响, 导致南海暖流对南海北部的影响减弱, 高温高盐种数量从而降低[46]。我国东海东部主要受黑潮和大洋水控制, 黑潮能将太平洋的大洋暖水种带到此[47]。这两大水系都属高温高盐性质, 导致本海区的浮游介形类主要以大洋暖水种为主, 分布符合高多样性、低丰度等热带大洋的动物区系特征[39]。
研究表明, 在厄尔尼诺年的秘鲁寒流海域中, 厄尔尼诺影响的海域介形类多样性和丰度比上升流海域高, 占主导数量的是针刺真浮萤()尤其在近海海域[48]。受厄尔尼诺影响的海区, 介形类出现频率增高[48]。厄尔尼诺现象导致溶解氧浓度升高, 同时, 在上层水中更多有机碎屑产生, 使得昼夜迁徙的种类更容易存活[48]。因此, 厄尔尼诺现象有利于许多介形类从热带水区入侵表层水域[48]。厄尔尼诺现象导致秘鲁寒流流经的海域温度异常升高, 使原本秘鲁寒流生态系统中丰富的浮游生物摄食者大量剧减, 会导致超微型浮游生物的生产周期发生改变[48]。介形类对盐度温度非常敏感, 因此温盐的不同能造成介形类数量的变化[48]。1998 年秘鲁寒流生态系统受厄尔尼诺现象影响, 海区温度升高, 易受温度变化的针刺真浮萤和易受温盐变化的条状原萤两个暖水种急剧增加, 导致厄尔尼诺年该海区介形类数量比非厄尔尼诺年更多[48]。
2.3 浮游介形类在海洋生态系统中的作用
海洋浮游介形类在海洋生态系统能量流动和物质循环中扮演着重要角色。日本海域的介形类具有较高的代谢活性, 是该海域高浮游生物生物量的主要贡献者, 并且是日本海中层生态系统能量流和物质循环的重要成员[49]。大量介形类被发现于鱼类的消化道中, 因此它们可能在有机物质的快速循环中有重要作用, 这些有机物来自死亡有机体、粪粒, 甚至絮状物。这和它们聚集于其它水面浮游生物的下方即中层水的现象一致, 此处有中深层游泳动物, 白天会有大量排泄物聚集。介形类的昼夜迁徙能将有机物质带回海水表面[45]。
某些介形类因自身特性及对环境的特殊要求, 只分布于某些适应的水域, 并且它们对水温和盐度变化敏感[4], 因此对各水系具有很好的指示作用, 能作为水团水系变化的指示种, 以及能成为气候变化的潜在指标, 对研究海洋生态结构和气候变化有重要作用。例如在东海, 广盐暖水种后圆真浮萤的密集区可指示黑潮锋、沿岸锋和对马暖流的大体位置; 近海暖水种针刺真浮萤的分布可体现沿岸混合水的动态; 贞女葱萤、大浮萤、亚弓浮萤和尖头毛浮萤等高温高盐的狭布大洋暖水种四季均分布于200 m等深线东侧, 可在春季扩布于200 m等深线西侧, 正好与此时的黑潮暖水舌侵入相吻合, 黑潮是高温高盐性质, 因此这些大洋暖水种能作为黑潮流域西界的良好指示种[39]。是深海冷水种, 只生活在冷水, 在极夜避开大西洋中层暖水层, 故是北大西洋的冷水指示种。而大西洋暖水的主要指示生物是[29]。北方肉萤()是北大西洋—北极的中深层种类, 能作为亚南极水团的生物指示种, 能够在大西洋向南扩散至30 °N。浮游介形类可能是水层中动物群落变化的指示者。深层介形类的聚集与水层变化有关, 因此这些群体发生的变化也许能够反映海洋气候的转变[45]。
3 我国海洋浮游介形类研究现状
我国海洋浮游介形类研究与国外相比, 研究历史短, 积累资料少。上世纪六十年代, 台湾学者曾文阳[50]描述了台湾海峡2 新种。陈瑞祥和林景宏[51]在描述西太平洋热带海域和东海浮游介形类种类的基础上, 综合多方面材料编著了《中国海洋浮游介形类》, 并发现了3 新种。尹健强和陈清潮等[52]对南海浮游介形类的多样性和动物地理区系进行了研究, 并在南海发现2 新种: 刘氏深海浮萤()[53]和缺刻深海浮萤()[54]。国家海洋局第三海洋研究所向鹏等人在南海发现了深海新种[55]。黄宗国[56]主编的《中国海洋生物种类与分布》和刘瑞玉[57]主编的《中国海洋生物名录》记述了中国海海腺萤科近百种, 国内已记录的种类(包括一些还需要澄清的种类)不到全球种类的二分之一。
在我国海域及附近海域, 统计海域主要包括黄海南部、东海、台湾海峡西部及邻近海域、巴士海峡、南海和北部湾东部, 浮游介形类共183 种, 包括2 亚目、4 科、8 亚科、52 属, 其中南海海域种类最多, 达122 种[58]。优势种是后圆真浮萤, 在四个季节中密度最高[38]。我国不同海区介形类海腺萤科的种类组成趋势与全球海域一致[38], 浮萤亚科记录的种类较多, 而深海浮萤亚科发现的种类较少, 主要分布在南海, 仍未深入研究。南海记录的海腺萤种类最多, 其次是东海(包括台湾海峡)(图3b)。基于现有数据和资料, 黄海记录的海腺萤种数较少, 有待进一步完善。
介形类在南海和东海的分布并不符合上述其它世界海域的分布规律。在南海中部, 浮游介形类的总丰度在垂直方向上具有明显的不同量级的分层现象[59]。0—100 m处最多达0.75 个·m-3, 随着深度增加而迅速减少, 100—200 m尚有0.61 个·m-3, 200—500 m层只有0.18 个·m-3, 到了500—1000 m水层仅有0.05 个·m-3, 1000—4000 m层极少, 为0.01 个·m-3[59]。南沙群岛海域50—500 m水层的介形类最丰富, 50 m以上的表层种类相对贫乏, 500 m以下数量随着温度下降而递减[60]。水平分布上, 介形类主要集中分布在南沙群岛海域西部、西北部和东南部, 而在其中部海域分布较少[61]。海洋浮游介形类在东海北部、济州岛以南海域常年高丰度, 在长江口及其邻近舟山海域数量相对较低, 在东海南部冬春季的高丰度区出现在外海, 夏秋季近海[62]。
在我国海域及附近海域, 统计海域主要包括黄海南部、东海、台湾海峡西部及邻近海域、巴士海峡、南海和北部湾东部, 浮游介形类共183 种, 包括2 亚目、4 科、8 亚科、52 属, 其中南海海域种类最多, 达122 种[58]。优势种是后圆真浮萤, 在四个季节中密度最高[38]。
国内海洋浮游介形类研究主要存在以下几方面的问题: (1)采样方面, 海洋浮游动物采样常使用505 μm网目孔径的浮游生物网具, 导致个体较小的介形类漏掉了, 并且大多浮游动物拖网深度一般为200 m至表层, 一些深层分布的种类未能采集到; (2)鉴定方面, 海腺萤个体细小、身体又完全被甲壳包被, 在鉴定过程中常作为一类、或仅鉴定常见种、或又多种误认为一种; (3)形态图方面, 国内有关海腺萤种类分类文献所提供的形态图有一定比例不是根据我国海域采集的标本所绘制, 而是引自其他作者及别的海区, 有些种类的形态特征因在不同海域的环境影响下会有所变化; (4)分类系统方面, 目前国内有关介形类分类系统的最新、最权威的工具书[56-57]仍使用Poulsen所建立的分类系统[20-21], 但如今介形类分类系统在原来基础上经多次修订已经发生了很大变化; (5)动物地理学方面, 研究方法上需要结合物理海洋学和生态学的研究结果, 才能更能清晰地阐明不同尺度上浮游介形类的地理分布。
针对以上存在的问题, 亟需加强我国深海浮游介形类的系统研究, 以便与国际研究水平接轨。南海通过马六甲海峡和吕宋海峡分别与印度洋和太平洋相通, 是海洋生物多样性的热点区域之一[10]。南海还是能量流、物流和基因流中心, 是开展深海研究的良好区域[63]。那么, 因其特殊的地理位置和复杂的水深地形, 南海深海环境中是否会存在一些未知海腺萤种类?其物种多样性的空间分布格局如何?跟其它海区比较, 南海深海浮游介形类的分类地位和地理分布规律有何不同?这些都是急需解决的科学问题。今后开展南海深海浮游介形类的研究, 将丰富我国深海浮游介形类的分类学研究, 为南海深海生物资源和生态过程等方面的研究提供科学依据, 在维护我国的海洋权益方面也具有一定的现实意义。
4 展望
海洋浮游介形类对海洋生态系统的研究具有不可替代的重要作用。今后海洋浮游介形类分类学研究的重要趋势是继续重视新种的发现, 特殊是深海环境的种类。太平洋和印度洋深海环境的介形类研究几乎没有开展, 应该会有更多的种类待发现, 兴起的DNA条形码是帮助介形类鉴定的良好手段。另外, 引入新的方法和研究海洋浮游介形类的碳含量和量化在碳循环中的作用, 这对介形类在海洋食物链中的角色以及与深海生态系统的研究有着重要的研究意义。
[1] SUTTON T T. Vertical ecology of the pelagic ocean: classical patterns and new perspectives[J]. Journal of Fish Biology, 2013, 83(6): 1508–1527.
[2] ANGEL M V. The diel migrations and distributions within a mesopelagic community in the north east Atlantic. 3. Planktonic ostracods, a stable component in the community [J]. Progress in Oceanography, 1984, 13(3): 245–268.
[3] ROBINSON C, STEINBERG D K, ANDERSON T R, et al. Mesopelagic zone ecology and biogeochemistry – a synthesis[J]. Deep Sea Research Part II Topical Studies in Oceanography, 2010, 57(16): 1504–1518.
[4] FASHAM M J R, ANGEL M V. The relationship of the zoogeographic distributions of the planktonic ostracods in the North–east Atlantic to the water masses[J]. Journal of the Marine Biological Association of the United Kingdom, 2009, 55(3): 739–757.
[5] HORNE D J, LORD A R, ROBINSON J E, et al. Ostracods as climatic indicators in interglacial deposits or, on some new and little–known British Quaternary Ostracoda[J]. Cfs Courier Forschungsinstitut Senckenberg, 1990, 123: 129–140.
[6] HOLMES J A, CHIVAS A R. The Ostracoda: Applications in Quaternary Research[M]. Washington, D. C: American Geophysical Union, 2002.
[7] HORNE D J, COHEN A, MARTENS K. Taxonomy, morphology and biology of quaternary and living Ostracoda [M]// HOLMES J A, CHIVAS A R. The Ostracoda: Applications in Quaternary Research. Washington, D. C: American Geophysical Union, 2002, 131: 5–36.
[8] ANGEL M V. Towards a full inventory of planktonic Ostracoda (Crustacea) for the subtropical Northwestern Atlantic Ocean[J]. Deep Sea Research Part II: Topical Studies in Oceanography, 2010, 57(24/26): 2173–2188.
[9] HORNE D J. Halocyprididae Dana, 1853[EB/OL]. (2004- 12-21)[2018-11-06]. http: //www. marinespecies. org/aphia. php? p=taxdetails&id=127482.
[10] CMARZ. Science Plan for the Census of Marine Zooplankton[R]. 2004.
[11] ANGEL M V.– a new genus of halocyprid ostracod, with the description ofnov. sp. and the redescription of(Scott, 1909) [J]. Zootaxa, 2012, 3254: 32–54.
[12] ANGEL M V. Planktonic ostracods (Myodocopa: Halocyprididae) from abyssopelagic depths in the Atlantic, North Pacific and Gulf of Oman:(n. gen. , n. sp. ),(n. sp. ),Chavtur and Stovbun, 2008 and(Müller, 1906) [J]. Zootaxa, 2013, 3709: 401–431.
[13] ANGEL M V, GRAVES C. Bathyconchoeciinae, a new subfamily of deep oceanic planktonic halocyprid Ostracod (Myodocopa, Ostracoda) [J]. Zootaxa, 2013, 3630: 243– 269.
[14] CHAVTUR V G, STOVBUN G O. A new genus and a new species of pelagic ostracods of the Subfamily Halocypridinae from the North Pacific[J]. Publications of the Seto Marine Biological Laboratory, 2008, 40(5/6): 47–54.
[15] CHAVTUR V G. Redescription ofChavtur, 1977 (Ostracoda, Halocyprididae) from the North Pacific[J]. Publications of the Seto Marine Biological Laboratory, 2014, 42: 1–13.
[16] CHAVTUR V G. Morphology and systematic of the ostracod(Myodocopa: Halocyprididae) from the North–Western Pacific[J]. Publications of the Seto MarineBiological Laboratory, 2014, 42: 14–41.
[17] CHAVTUR V G, BASHMANOV A G.sp. nov. , a new deep–sea species of halocypric (Ostracoda, Myodocopa) from the northern Pacific[J]. Crustaceana, 2018, 91(8): 1001–1011.
[18] GRAVES C. Redescription of(Müller, 1890) and(Scott, 1894) (Halocyprididae: Ostracoda) from the Atlantic, and descriptions of two novel species ofMüller 1890, from the Gulf of Oman[J]. Journal of Natural History, 2011, 45(31/32): 1937–1981.
[19] GRAVES C.(Halocyprididae: Ostracoda) a new genus and species from the Northwest Indian Ocean[J]. Journal of Natural History, 2012, 46(9/10): 517– 556.
[20] POULSEN E M. Ostracoda–Myodocopa. Part III A. Halocypriformes–Thaumatocypridae and Halocypridae[J]. Dana Report, 1969, 75: 1–99.
[21] POULSEN E M. Ostracoda–Myodocopa. Part III B. Halocypriformes–Halocypridae Conchoecinae[J]. Dana Report, 1973, 84: 1–224.
[22] CHAVTUR V G, STOVBUN G O. Rivision of pelagic ostracods of the subfamily Archiconchoecinae[J]. Publications of the Seto Marine Biological Laboratory, 2003, 39 (4/6): 139–219.
[23] CHAVTUR V G, ANGEL M V. Revision of(Ostracoda: Halocyprididae) and the designation of two new tribes Conchoeciini and Metaconchoeciini[J]. Zootaxa, 2011, 2857: 1–87.
[24] DEEVEY G B., a new genus of pelagic ostracods (Myodocopa, Halocyprididae) with six new species from the deeper waters of the Gulf of Mexico[J]. Proceedings of the Biological Society of Washington, 1968, 81: 539–570.
[25] DEEVEY G B. A taxonomic and distributional study of the planktonic ostracods collected on three cruises of the “Eltanin” in the south Pacific and the Antarctic Region of the South Pacific[J]. Biology of the Antarctic Seas, VIII Antarctic Research, 1978, 28 (3): 43–70.
[26] GEORGE J, NAIR V R. Planktonic ostracods of the northern Indian Ocean[J]. Mahasagar–Bulletin of the National Institute of Oceanography, 1980, 13(1): 29–44.
[27] GEORGE J. A preliminary report on the distribution and abundance of planktonic ostracods in the Indian Ocean[J]. Bulletin of the National Institute of Sciences of India, 1969, 38: 641–648.
[28] CHAVTUR V G, KRUK N V. Vertical distribution of pelagic ostracods (Ostracoda, Halocyprinidae) in the Australian–New Zealand sector of the Southern Ocean[J]. Russian Journal of Marine Biology, 2003, 29(2): 90–99.
[29] BASHMANOV A G, CHAVTUR V G. Structure and distribution of pelagic ostracods (Ostracoda: Myodocopa) in the Arctic Ocean[J]. Russian Journal of Marine Biology, 2009, 35(5): 359–373.
[30] ANGEL M V. Ostracoda[M]//BOLTOVSKOY D. South Atlantic Zooplankton Volume 1. Netherland: Bachhuys, 1999: 815–868.
[31] DEEVEY G B, and BROOKS A L. The planktonic ostracods of the Sargasso Sea off Bermuda: species composition and vertical and seasonal distribution between the surface and 2000 m[J]. Biological Science, 1980, 26 (2): 37–124.
[32] KAPLUN N L, MAZDYGAN E R, CHAVTUR V G, et al. Changes in the frequency of the occurrence and abundance of ostracods (Ostracoda: Halocyprididae) in the epipelagic zone of the North Pacific[J]. Russian Journal of Marine Biology, 2015, 41(3): 157–166.
[33] CHAVTUR V G. Fauna of the pelagic ostracods of Halocypridae family from the north western Pacific[J]. Biologiya Morya Vladivostok, 1977, 1: 28–38.
[34] CHAVTUR V G, MAZDYGAN E R. Composition and distribution of pelagic ostracods (Ostracoda: Myodocopa) in the Somov and Ross seas and adjacent waters of the Southern Ocean[J]. Russian Journal of Marine Biology, 2011, 37(3): 165–176.
[35] NIGRO L M, ANGEL M V, BLACHOWIAK–SAMOLYK K, et al. Identification, discrimination, and discovery of species of marine planktonic ostracods using DNA barcodes[J]. Plos One, 2016, 11(1): e0146327.
[36] BUCKLIN A, ORTMAN B D, JENNINGS R M, et al. A “Rosetta Stone” for metazoan zooplankton: DNA barcode analysis of species diversity of the Sargasso Sea (Northwest Atlantic Ocean)[J]. Deep–Sea Research Part II, 2010, 57(24): 2234–2247.
[37] XU Lei, WANG Lianggen, NING Jiajia, et al. Diversity of marine planktonic ostracods in South China Sea: a DNA taxonomy approach[J]. Mitochondrial Dna Part A Dna Mapping Sequencing & Analysis, 2018.
[38] 陈瑞祥, 林景宏. 中国海及其邻近海域浮游介形类大尺度生态研究Ⅲ. 浮游介形类的物种与生态类群多样性[J]. 生物多样性, 1997, 5(4): 257–262.
[39] 林景宏, 陈明达, 陈瑞祥. 东海浮游介形类对海流、水团的指示作用[J]. 海洋学报, 1996, 18(3): 86–91.
[40] CHAVTUR V G, BASHMANOV A G. Distribution and seasonal dynamics of Boroecia maxima, (Ostracoda: Halocypridinae) in the Arctic basin and adjacent Atlantic waters[J]. Russian Journal of Marine Biology, 2007, 33(2): 75–83.
[41] ANGEL M V. Planktonic oceanic ostracods—historical, present and future[J]. Proceedings of the Royal Society of Edinburgh Section B (biology), 1972, 73(22): 213–228.
[42] LONGHURST A R. Relationship between diversity and the vertical structure of the upper ocean[J]. Deep Sea Research Part A Oceanographic Research Papers, 1985, 32(12): 1535–1570.
[43] GIBBONS M J, RICHARDSON A J, ANGEL M V, et al. What determines the likelihood of species discovery in marine holozooplankton: is size, range or depth important? [J]. Oikos, 2005, 109(3): 567–576.
[44] APPELTANS W, AHYONG S T, ANDERSON G, et al. The magnitude of global marine species diversity[J]. Current Biology, 2012, 22(23): 2189–2202.
[45] ANGEL M V, BLACHOWIAK–SAMOLYK K, DRAPUN I. , et al. Changes in the composition of planktonic ostracod populations across a range of latitudes in the North–east Atlantic[J]. Progress in Oceanography, 2007, 73(1): 60–78.
[46]陈瑞祥, 林景宏. 中国近海浮游介形类大尺度生态研究II. 浮游介形类与水系的相关研究[J]. 海洋学报, 1998, 20(2): 96–101.
[47] 徐兆礼. 东海浮游介形类生态适应分析[J]. 海洋学报, 2007, 29(5): 123–131.
[48] CASTILLO R, ANTEZANA T, AYON P. The influence of El Niño 1997–98 on pelagic ostracods in the Humboldt Current Ecosystem off Peru[J]. Hydrobiologia, 2007, 585(1): 29–41.
[49] IKEDA T. Ecological and biological features of a mesopelagic ostracod,, in the Japan Sea[J]. Marine Biology, 1990, 107(3): 453–461.
[50] TSENG Wenyang.(Ostracoda) from Taiwan Straits[J]. Reports of the Taiwan Laboratory of Fish Biology, 1969, 19: 1–26.
[51] 陈瑞祥, 林景宏. 中国海洋浮游介形类[M]. 北京: 海洋出版社, 1995: 1–134.
[52] 尹健强, 陈清潮. 南沙群岛及其邻近海区浮游介形类的种类、动物区系和动物地理[M]// 中国科学院南沙综合科学考察队, 南沙群岛海区海洋动物区系和动物地理研究专集. 北京: 海洋出版社. 1991: 64–139.
[53] YIN Jianqiang, CHEN Qingchao, LI Kaizhi.n. sp., a new species of ostracod (Myodocopa, Halocyprididae) from the South China Sea[J]. Crustaceana, 2014, 87(8/9): 1027–1035.
[54] YIN Jianqiang, LI Kaizhi, TAN Yehui.sp. nov. (Myodocopa, Halocyprididae), a new species of ostracod from the neritic zone of the South China Sea[J]. Crustaceana, 2017, 90(1): 35–48.
[55] XIANG Peng, WANG Yu, CHEN Ruixiang, et al. A bathypelagic ostracodsp. nov. (Myodocopa, Halocyprididae) from the South China Sea[J]. Peerj, 2018: 6.
[56] 黄宗国. 中国海洋生物种类与分布[M]. 北京: 海洋出版社, 2008: 541–548.
[57] 刘瑞玉. 中国海洋生物名录[M]. 北京: 科学出版社, 2008: 635–641.
[58] 陈瑞祥, 林景宏. 中国近海浮游介形类大尺度生态研究: Ⅰ. 浮游介形类的丰度及其变动规律[J]. 海洋学报, 1998, 20(1): 94–100.
[59] 陈瑞祥, 林景宏. 南海中部海域浮游介形类的生态研究[J]. 海洋学报, 1993, 15(6): 91–98.
[60] 尹健强, 陈清潮. 南沙群岛海区的浮游介形类(1984–1988)[M]// 中国科学院南沙综合科学考察队, 南沙群岛及其邻近海区海洋生物研究论文集(二). 北京: 海洋出版社, 1991: 64–139.
[61] 郭伟龙, 王亮根, 杜飞雁, 等. 南沙群岛海域春、秋季浮游介形类生态学特征[J]. 水产学报, 2015, 39(6): 836– 845.
[62] 徐兆礼. 东海浮游介形类(Ostracods)分布特征[J]. 海洋学报, 2006, 28(2): 101–108.
[63] 汪品先. 南海—我国深海研究的突破口[J]. 热带海洋学报, 2009, 28(3): 1–4
Advances on species diversity and ecology of marine pelagic ostracods
LIAO Tongchen1,2, YIN Jianqiang1, LI Kaizhi1,3,4*, TAN Yehui1,2,3,4
1. Key Laboratory of Tropical Marine Bio–resources and Ecology, South China Sea Institute of Oceanology, ChineseAcademy of Sciences, Guangzhou510301,China 2. University of Chinese Academy of Sciences, Beijing 100049, China 3. Guangdong Provincial Key Laboratory of Applied Marine Biology,South China Sea Institute of Oceanology, Chinese Academy of Sciences, Guangzhou 510301,China 4. Innovation Academy of South China Sea Ecology and Environmental Engineering, Chinese Academy of Sciences, Guangzhou 510301, China
Planktonic ostracods are a group of small crustaceans with a wide distribution all over the world’s oceans. Most of the pelagic ostracods feeding on a variety of detritus are important food sources for deep–living fishes, and therefore they are vital to the vertical carbon flux in the oceans. The vast majority of references about marine pelagic ostracods focused on the upper ocean, and hence 200-400 species might not be found, especially the bathypelagic species. Report on the new bathypelagic species may be the key trends of taxonomy of Halocyprididae. Because of the few focus on the bathypelagic ostracods in our country, we have to go into more details in bathypelagic ostracods in the future. In this paper, species diversity and ecology of marine pelagic ostracods and taxonomy of Halocyprididae are summarized. Species diversity and distribution of ostracods in different oceans are compared and analyzed. The ecological functions of marine pelagic ostracods in the ocean as well as the status in our country of study in marine pelagic ostracods are summarized, which will provide a scientific baseline for marine species diversity, marine biogeochemical cycles and abyssopelagic ecology.
biodiversity; zoogeography; ecology; Halocyprididae; Ostracoda
10.14108/j.cnki.1008-8873.2019.06.028
S963.21+4
A
1008-8873(2019)06-190-09
2018-11-05;
2018-12-22
国家自然科学基金项目(41576125); 国家科技基础资源调查专项(2017FY201404); 中国科学院南海生态环境工程创新研究院自主部署项目(ISEE2018PY01)
廖彤晨(1995—), 女, 广东佛山人, 硕士研究生, 主要从事海洋浮游动物生态学研究, E-mail: liaotongchen@foxmail.com
李开枝, 女, 博士, 研究员, 主要从事海洋浮游动物多样性和生态学研究, E-mail: likaizhi@scsio.ac.cn
廖彤晨, 尹健强, 李开枝, 等. 海洋浮游介形类的多样性和生态学研究进展[J]. 生态科学, 2019, 38(6): 190-198.
LIAO Tongchen, YIN Jianqiang, LI Kaizhi,et al. Advances on species diversity and ecology of marine pelagic ostracods[J]. Ecological Science, 2019, 38(6): 190-198.
猜你喜欢
潍坊学院学报(2020年2期)2021-01-18
海洋通报(2020年5期)2021-01-14
流行色(2019年10期)2019-12-06
小哥白尼(趣味科学)(2019年7期)2019-11-16
收藏界(2018年1期)2018-10-10
创新作文(小学版)(2018年31期)2018-05-16
摄影之友(影像视觉)(2017年1期)2017-07-18
学苑创造·A版(2017年4期)2017-05-13
学苑创造·A版(2017年5期)2017-05-09
小学阅读指南·低年级版(2016年6期)2016-05-14
