
文昌鸡白麻羽性状与SLC45A2多态性关系
2019-12-23 01:42:48何艺平高鑫凤许继国
广东农业科学 2019年10期
何艺平,陈 杰,高鑫凤,许继国
(1.广州市畜牧科学研究所,广东 广州 510550;2.南昌师范学院生物技术研究所/江西省地方鸡种遗传改良重点实验室/中国科学院家禽分子育种技术联合实验室,江西 南昌,330022;3.江丰实业股份有限公司,广东 广州 510450;4.华南农业大学动物科学学院,广东 广州 510642)
【研究意义】通过有意识的育种,家养动物群体聚集了丰富的具有性状效应(phenotypic effects)的变异[1]。作为全球生物多样性重要组成部分的家养动物,不仅为人类提供了稳定的动物蛋白来源及许多有用的动物产品,其性状多样性还是研究基因型性状之间关系的重要资源。实验动物的变异常常具有病理性性状,而家养动物中具有性状效应的变异大部分不具有病理性性状,是遗传学研究的重要组成部分[2-3]。家鸡的色素沉着性状作为家养动物性状多样性的代表,备受生物学家和育种工作者关注。研究色素性状的遗传对于探讨禽类起源、演化、生产以及开展实验动物研究等方面都具有重要意义。
【前人研究进展】目前,在家鸡中已经鉴定到了11个致因突变与羽色性状相关,涉及10个基因[4-14],其中2个为伴性遗传(银色羽和芦花羽基因座位)[11,14],10个为常染色体遗传的基因座位。白麻羽性状(亦称为珍珠白)是文昌鸡存在的一种较为典型的性状,与黄麻羽性状相比其主要特点是不能合成嗜黑素,并且不改变羽衣中真黑素和嗜黑素的相对分布(图1)。关于文昌鸡白麻羽的遗传规律以及其形成的分子机制未见报道。根据白麻羽的性状特征和遗传方式以及Gunnarsson等[11]研究基础,我们推测其是由银色羽位点突变所致。
【本研究切入点】本研究首先检测了已知的银色羽突变位点与白麻羽性状的关系,结果显示其并非致因突变。之后通过正反交试验探明了白麻羽性状的伴性隐性遗传规律。结合白麻羽性状不能合成嗜黑素、不改变色素相对分布的特点以及其伴性遗传的特征,我们首先猜测其很可能是位于银色羽基因座位上一个新的等位基因,因此重新筛查了银色羽对应的SLC45A2基因的多态性,并验证了多态位点与白麻羽性状之间的关系。从性状来看,与野生型相比,白麻羽个体发生的是嗜黑素(黄色-红色)合成障碍而并非嗜黑素和真黑素(褐色-黑色)的相对分布变化,因此我们猜测很可能是由于SLC45A2基因外显子区域的错义突变或者无义突变导致了嗜黑素合成障碍[11]。因此,本研究首先验证了外显子区域的错义或者无义突变与白麻羽性状的关系。【拟解决的关键问题】探索白麻羽性状的遗传规律,并找出对应的致因突变。研究结果可为白麻羽群体在育种中的利用以及相应性状的分子标记辅助选择提供依据。

图1 白麻羽文昌鸡(左)和黄麻羽文昌鸡(右)Fig.1 Wenchang chicken with white-partridge feathers(Left)and Wenchang chicken with yellow-partridge feathers(Right)
1 材料与方法
1.1 试验材料
供试鸡只来源于广州市江丰实业股份有限公司育种场,其中白麻羽和黄麻羽公鸡各1只,白麻羽和黄麻羽母鸡各15只。禽血DNA抽提试剂盒购买自广州飞扬生物科技有限公司。分子试验部分所用离心机、PCR等仪器设备来源于广东省农业动物基因组学与分子育种重点实验室。
1.2 试验方法
1.2.1 正反交试验 分别用1只白麻羽公鸡与15只黄麻羽母鸡杂交,以及1只黄麻羽公鸡与15只白麻羽母鸡杂交。记录每只种蛋后代对应的父母本,入孵出雏后佩戴翅号,并且记录其父母本信息,6周龄时统计公母鸡羽色分布。
1.2.2 DNA抽提 翅下静脉采血,放于装有抗凝剂的1.5 mL离心管中。根据OMEGA试剂盒说明书进行血样DNA抽提。
1.2.3 多态位点筛选 根据候选基因SLC45A2序列设计特异性引物(表1),选用3个白麻羽纯合子和6个黄麻羽纯合子作为模板,对候选基因SLC45A2外显子区域进行重测序,引物位置设计在外显子两侧100 bp左右。利用DNAStar中的SepMan软件读取测序结果,并进行SNP筛选。

表1 供试引物信息Table 1 Information of the primers for trials
1.2.4 候选位点验证 以白麻羽文昌鸡、黄麻羽文昌鸡、固始鸡、霸王山鸡、清远麻鸡、杏花鸡和红色原鸡DNA为模板,设计特异性引物(表1),利用测序的方法对候选变异位点进行分型。
2 结果与分析
2.1 正反交试验结果
白麻羽公鸡与黄麻羽母鸡杂交,后代公鸡全部能合成嗜黑素,表现为黄芦花性状(黄白相间);母鸡不能合成嗜黑素,全部为白麻羽性状。黄麻羽公鸡与白麻羽母鸡杂交,后代公鸡和母鸡均能够合成嗜黑素,公鸡表现为黄芦花性状,母鸡表现为黄麻羽性状(表2)。正反交试验结果表明,白麻羽性状呈现伴性隐性遗传。

表2 正反交试验结果统计Table 2 Result of the reciprocal cross experiment
2.2 SLC45A2多态性与白麻羽关系研究
2.2.1SLC45A2多态位点筛选 根据白麻羽性状不能合成嗜黑素且不改变色素相对分布的特征、伴性遗传的规律以及前人关于鸡银色羽性状和白虎毛色形成分子机制的研究结果[11,15-16],选取SLC45A2作为候选基因,筛选多态位点。以白麻羽和黄麻羽性状个体的DNA为模板,对SLC45A2外显子区域进行重测序,结果共筛选到5个SNP(表3),其中3个位于外显子上,2个为错义突变(SNP1、SNP4),1个为同义突变(SNP2)。
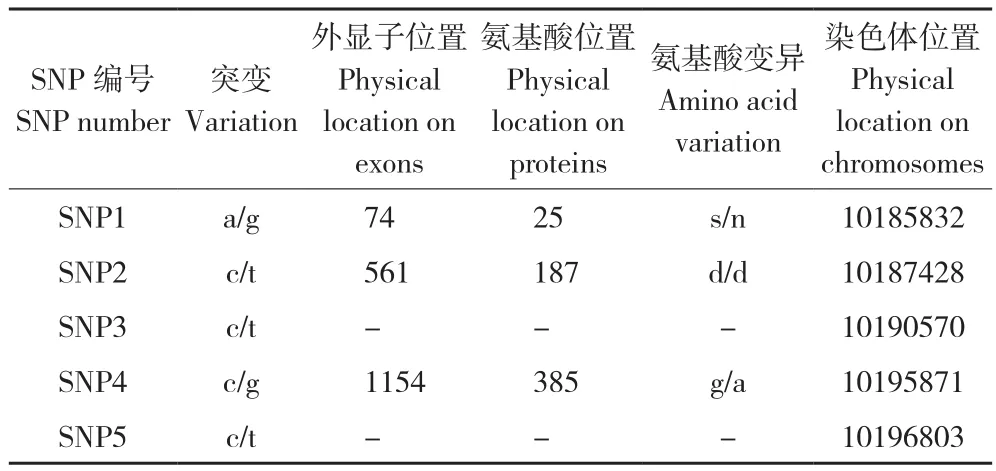
表3 SLC45A2基因SNPs位点信息Table 3 SNPs location information of SLC45A2 gene
2.2.2 候选位点验证 根据白麻羽性状不能合成嗜黑素以及不改变嗜黑素和真黑素相对分布的特点,推测与该性状相关的突变是错义突变,因此首先排除SNP2、SNP3和SNP5位点;又由于该性状为隐性性状,因此与该性状相关的SNP位点在白麻羽群体中应该呈现纯合子,因此排除SNP1位点(利用白麻羽性状和黄麻羽性状进行SNP筛选时,SNP1在白麻羽群体中出现了杂合子,数据未显示)。根据上述分析,我们选取SNP4作为候选位点,以不能合成嗜黑素的白麻羽文昌鸡与能合成嗜黑素的黄麻羽文昌鸡、固始鸡、霸王山鸡、清远麻鸡、杏花鸡和红色原鸡为材料,验证SNP4位点与白麻羽性状的关系。
采用Sanger测序的方法对被检测个体的SNP4位点进行分型,结果显示,所有能合成嗜黑素的品种均为GG基因型,而不能合成嗜黑素的白麻羽文昌鸡均为CC基因型(表4)。上述结果显示,SNP4位点(SLC45A2基因编码区1154位点)的C等位基因与白麻羽性状完全连锁,很可能是白麻羽性状的致因突变。

表4 SNP4位点在不同性状群体中的分布情况Table 4 Distribution of SNP4 in different populations
3 讨论
SLC45A2基因编码区1154位点与文昌鸡白麻羽性状完全连锁,可能是银色羽基因座上的一个新的等位基因。Gunnarsson等研究发现,具有相似性状的国外品种是由于SLC45A2基因上的突变位点〔C.C1039A(L347M)〕导致的[11]。在白麻羽文昌鸡中虽然不存在该突变,但SLC45A2基因编码区1154位点突变却与白麻羽性状呈完全连锁的关系,可能该突变是一个新的银色羽等位基因,而且这一突变很可能是致因突变,其原因有3个方面:(1)试验结果显示,该突变与文昌鸡白麻羽性状完全连锁,并未在能够合成嗜黑素的品种中发现该突变;(2)白麻羽性状的特点是不能够合成嗜黑素,可能是由于基因编码区内发生错义突变导致的,本研究中鉴定到的突变符合上述条件;(3)本试验结果与白虎(不能合成嗜黑素但是可以合成真黑素)中的研究相似,白虎的形成也是由于SLC45A2基因上的一个错义突变(A477V)导致的[15]。
由于动物的毛色是由真黑素和嗜黑素两种色素在毛发中的相对分布导致的[16-18],因此我们有理由相信大部分羽毛性状是由于调控区的调节突变影响了参与色素合成的基因表达而导致的。大部分羽色多样性都是由嗜黑素和真黑素组成,其不同只是表现为嗜黑素和真黑素的相对分布,嗜黑素和真黑素合成能力并没有差别[19]。本研究中白麻羽性状相对于黄麻羽性状,其典型特征是不能合成嗜黑素,并且不改变色素的相对分布。因此,我们首先推测其很可能是编码区的错义或者无义突变导致基因无法行使正常的功能而导致的。本试验结果较好地支持了这一猜测。
本研究在候选位点验证时,选取了能合成嗜黑素的品种为相对性状,在被测的群体中显示SLC45A2基因编码区1154位点在能合成嗜黑素的群体中不携带C等位基因。囿于条件限制,被用于验证的6个群体5个在我国的广东省和海南省,这降低了验证的效力。未来可扩大验证样本的取样范围,进一步验证SLC45A2基因编码区1154位点与白麻羽性状的关系。另外,还需要在细胞层面以及通过基因编辑的方式对SLC45A2基因编码区1154位点进行功能验证以及分析其对于白麻羽性状形成的机制。SLC45A2基因突变导致白麻羽性状的分子机理尚不清楚,因此还需要进一步研究。目前有关SLC45A2只影响嗜黑素的合成而不影响真黑素的合成的原因并没有十分系统的试验数据,但众多研究结果提供了一些启示,很可能是SLC45A2基因的错义突变减少了机体全身黑色素的生物合成,但是高表达的EDN3维持了某些区域真黑素的合成[15]。
4 结论
本研究以分离出的白麻羽个体与黄麻羽个体进行正反交试验,结果显示白麻羽突变表型相对于野生型的黄麻羽为伴性隐性遗传。根据白麻羽表型伴性隐性遗传方式、白麻羽表型不能合成嗜黑素的表型特征(一般为基因功能缺失造成)和前人研究结果,我们选取SLC45A2基因外显子区域的错义突变作为候选致因突变进行了验证。结果显示,SLC45A2基因编码区1154多态位点能将白麻羽群体与非白麻羽群体分开。综合上述结果,得出初步结论,SLC45A2基因编码区1154多态位点很可能是导致白麻表型的致因突变。该突变与白麻表型形成之间的分子机制有待进一步研究。
猜你喜欢
广东药科大学学报(2023年5期)2023-12-30 00:08:39
农产品市场周刊(2022年14期)2022-08-10 07:06:54
时代报告·奔流(2022年1期)2022-04-29 04:10:56
昆明医科大学学报(2022年3期)2022-04-19 13:59:42
纺织科学与工程学报(2020年1期)2020-06-12 09:14:54
雨花(2018年10期)2018-11-15 11:59:26
农产品市场周刊(2018年26期)2018-07-13 03:24:48
军营文化天地(2016年10期)2016-06-15 20:28:32
浙江医学(2014年17期)2014-04-13 10:13:16
汉语世界(2012年2期)2012-03-25 13:01:29
