
不同成熟度树葡萄叶片中类黄酮合成转录组基因分析
2019-12-06 09:05张少平郑开斌洪佳敏林宝妹张帅邱珊莲
热带亚热带植物学报 2019年6期
张少平, 郑开斌, 洪佳敏, 林宝妹, 张帅, 邱珊莲*
不同成熟度树葡萄叶片中类黄酮合成转录组基因分析
张少平1,2, 郑开斌1,2, 洪佳敏1, 林宝妹1, 张帅1, 邱珊莲1*
(1. 福建省农业科学院亚热带农业研究所,福建 漳州 363005; 2. 福建省农业科学院作物研究所,福州 350013)
为了解树葡萄()类黄酮合成相关酶差异表达基因信息,对其幼叶和成熟叶进行全转录组测序并比较分析。结果表明,从树葡萄幼叶和成熟叶中获得59 321条单基因簇(Unigenes),在8大数据库共注释到32 912条Unigenes信息,其中类黄酮合成代谢相关酶基因77个,在成熟叶片中显著下调表达的基因6个,包括2个、1个、1个、1个2-羟基异黄酮脱水酶基因和1个。5个差异表达基因经qRT-PCR验证的结果与转录组测序结果相符合。因此,树葡萄叶片中含有大量不同种类黄酮合成代谢相关酶家族基因,成熟叶片中类黄酮含量显著减少是由于少量相关基因显著下调。
树葡萄;叶片;类黄酮;转录组基因
树葡萄()俗称珍宝果、嘉宝果、拟爱神木等,为桃金娘科(Myrtaceae)拟爱神木属常绿灌木或乔木,原产于巴西等南美地区[1]。我国对树葡萄引种栽培较早的地区始于台湾[2],近年来,随着人们对树葡萄的青睐,福建沿海等地也逐渐出现较大面积引种栽培[3]。树葡萄一年可多次开花,但果实盛产期集中在春秋两季,花果主要着生于树干或粗壮的枝条上。树葡萄不仅是集果实食用和观赏为一体的名贵优良树种[4],而且全株(包括根、茎、叶、花和果实)富含类黄酮等活性成分[5-6],具有抗氧化、抗炎、抗肿瘤及降低胆固醇和降血糖血脂等多种功效[7-9]。类黄酮是植物中多酚类次生代谢产物[10],主要成分包括查尔酮、黄烷酮、黄酮、异黄酮、二氢黄酮、黄酮醇及花青苷等[11-13]。
类黄酮合成代谢途径涉及的酶主要包括查尔酮合成酶(chalcone synthase, CHS)、查尔酮异构酶(chalcone isomerase, CHI)、黄烷酮3-羟化酶(flavanone 3-hydroxylase, F3H)(或黄酮醇合成酶 flavonol synthase, FLS)、类黄酮3ʹ-羟化酶(flavonoid 3-hydroxylase, F3ʹH)(或类黄酮3-单加氧酶flavonoid 3-monooxygenase)、类黄酮3ʹ,5ʹ-羟化酶(flavonoid 3,5-hydroxylase, F3ʹ,5ʹH)、二氢黄酮醇4-还原酶(dihydroflavonol/flavone 4-reductase, DFR)、黄酮合成酶(flavone synthase,FSI和FSII)、异黄酮还原酶(isoflavone reductase, IFR)、异黄酮合成酶(isoflavone synthase, IFS)(或异黄酮羟化酶isoflavone hydro- xylase)、花青素合成酶(anthocyanidin synthase, ANS)(或无色花色素双加氧酶leucoanthocyanidindioxygenase, LDOX)、花青素3--糖基转移酶(anthocyanidin 3--glucosyltransferase, 3GT)(或类黄酮3--糖基转移酶flavonoid 3--glucosyltrans-ferase; 或UDP-葡萄糖UDP-glucose)、无色花色素还原酶(leucoanthocyanidin reductase, LAR)、花青素还原酶(anthocyanidin reductase, ANR)等[14-16]。近年来,有关树葡萄的研究大多集中在栽培管理、营养成分、药理功效及产品加工等方面[17-19],而对树葡萄类黄酮代谢合成相关基因的研究还未见报道。树葡萄叶片富含类黄酮,尤以嫩叶含量最高[20-21],因此,本研究将以树葡萄栽培种沙巴幼叶为对象,以成熟叶为对照,利用高通量测序技术进行转录组基因测序,通过不同数据库注释后再进行类黄酮合成代谢中所涉及关键酶基因检索,以期获得叶片中类黄酮合成代谢相关基因的种类、表达量及差异表达信息,为深入研究树葡萄或其他特色植物中类黄酮合成代谢奠定了基础。
1 材料和方法
1.1 材料
从福建省农业科学院亚热带农业研究所天然产物源种质资源圃中,取15 a生树葡萄栽培种沙巴(‘Shaba’)的嫩叶和成熟叶,嫩叶为春季新长出20 d内的叶片,红色;成熟叶为30~60 d的叶片,嫩绿色。两种叶片分别标号后送北京百迈克生物科技有限公司进行RNA提取、转录组测序、测序数据组装和差异表达基因的qRT- PCR验证等。
1.2 RNA提取和高通量测序
采用Trizol法分别提取树葡萄嫩叶和成熟叶的总RNA,各3份共6个样本,每个样本检测合格后取等量混匀组成1个RNA池,再分别进行带有Oligo(dT)磁珠富集叶片mRNA并进行随机打断、反转录成双链cDNA再进行纯化和末端修复、加A尾后连接测序接头、PCR富集制备测序文库及测序等。
应用Illumina HiSeq测序平台,构建树葡萄叶片转录组文库后,测序并获取相关数据。所得原始数据通过过滤去除接头序列及低质量读序以获取高质量的干净读序。将干净读序进行序列组装,最后获得树葡萄栽培种沙巴叶片的单基因簇(Unigenes)库。
1.3 功能注释及关键基因检索
转录组测序获取树葡萄叶片相关数据后,分别进行随机性检验和饱和度检验等测序文库质量评估,合格后的数据进行表达量分析。使用BLAST软件[22]对树葡萄叶片所有Unigenes在不同数据库进行功能注释,这些数据库有蛋白质直系同源数据库(cluster of orthologous groups, COG)、东京基因与基金组百科全书(Kyoto encyclopedia of genes and genomes, KEGG)、基因本体论数据库(gene ontology, GO)、蛋白质真核同源数据库(euKaryotic orthologous groups, KOG)、蛋白质序列数据库(swiss-prot protein database, Swiss-Prot)、蛋白质家族域数据库(protein families database, Pfam)、直系同源蛋白的功能描述和功能分类(evolutionary genealogy of genes: Non- supervised orthologous groups, eggNOG)和非冗余蛋白数据库(non-redundant protein database, NR)。经数据库注释后,进一步在Swiss-Prot和NR数据库进行类黄酮合成相关基因检索分析,以期获得树葡萄幼叶和成熟叶片中类黄酮合成基因种类和表达量。
1.4 差异表达基因的qRT-PCR分析验证
通过qRT-PCR检测2个树葡萄样本在转录组测序中差异表达的5个类黄酮合成相关基因(表1)的表达。首先,树葡萄幼叶和成熟叶各取1g RNA,使用TransScript II All-in-One First-Strand cDNA Synthesis SuperMix for qPCR (One-Step gDNA Removal) (货号:AH341)试剂盒,根据试剂盒说明书的方法进行反转录成cDNA。然后应用SYBRGreen染料进行qRT-PCR检测分析,均设置3次重复。
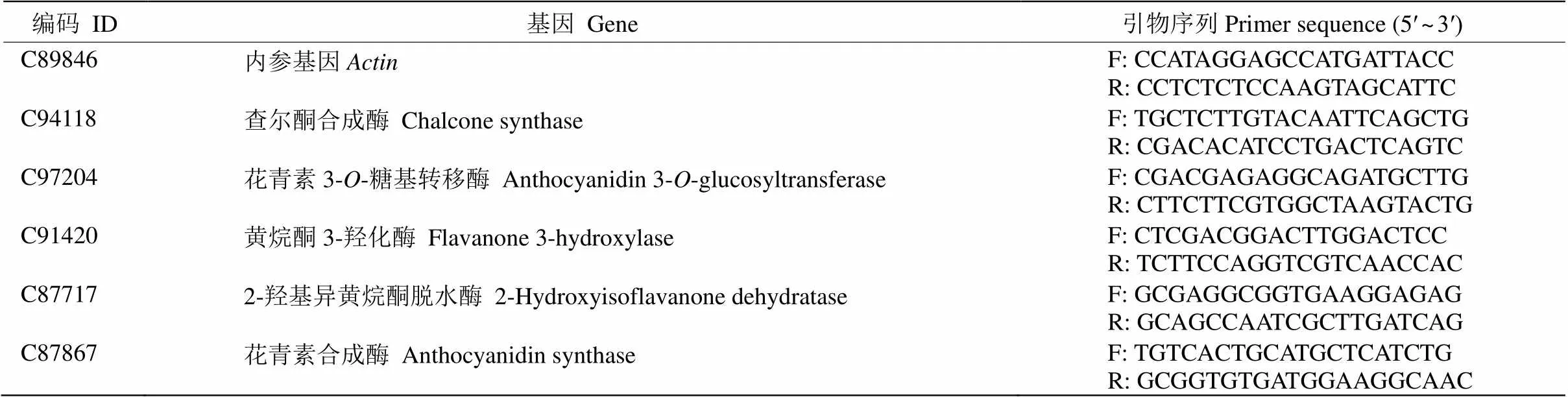
表1 qRT-PCR分析的5个基因和引物
2 结果和分析
2.1 测序分析
通过对树葡萄幼叶和成熟叶转录组进行测序和合并组装,共获得16.53 Gb的clean data,各样品的clean data均达到7.30 Gb,Q30碱基≥92.83%。组装后共获得59 321条Unigenes,其中长度在1 kb以上的有17 623条。所有Unigenes通过COG、KEGG、GO、KOG、Swiss-Prot、Pfam和NR等数据库进行注释比对,共有32 912条注释, 其中长度在300 bp~ 1 kb的Unigenes为17 592条,而长度大于1 kb的有15 320条(表2)。
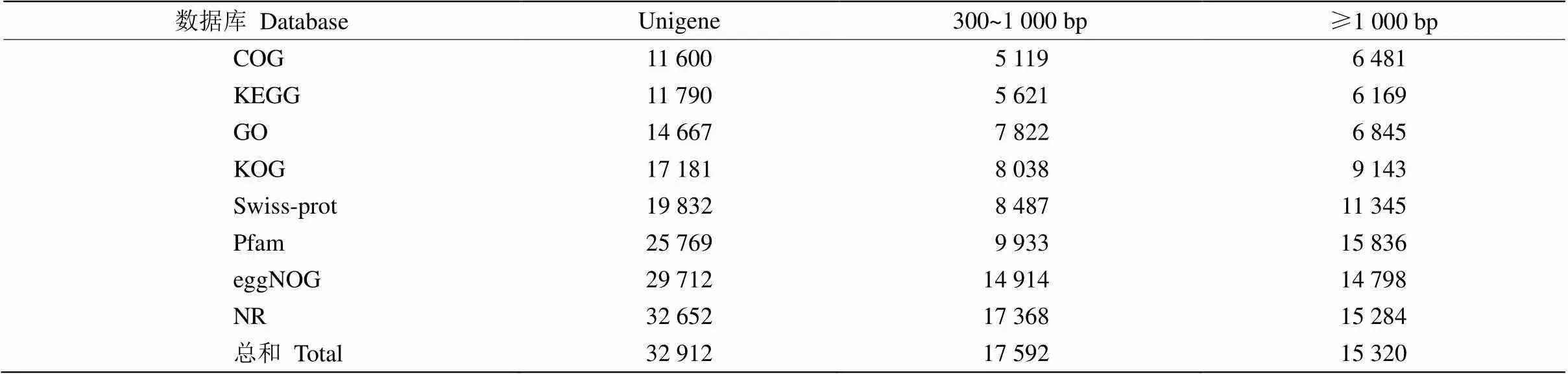
表2 数据库注释的Unigenes数量
2.2 类黄酮相关基因分析
2.2.1 查尔酮合成酶相关基因
所有32 912条Unigenes通过chalcone (查尔酮)关键词检索,依据NR和Swiss-Prot数据库分析, 共获得9个查尔酮合成酶相关基因(表3),包括3个和6个基因。差异表达极显著下调的基因有3个,包括2个(编码c93272和c86311)和1个基因(编码c94118),这3个查尔酮相关核心基因在树葡萄幼叶中的FPKM值明显高于成熟叶片;同时,上述9条Unigenes在NR数据库匹配的物种主要有巨桉(, 6条),而在Swiss-Prot数据库匹配的物种有拟南芥(, 2条)和茶树(, 2条)等7种植物。

表3 9个查尔酮合成酶相关基因
2.2.2 类黄酮相关基因
通过flavonoid(类黄酮)关键词检索,依据NR及Swiss-Prot数据库分析,共获得6个类黄酮相关基因(表4),包括3个和3个,基因, 这些基因在树葡萄幼叶和成熟叶中的表达差异不显著,因此,该两类家族基因在树葡萄叶片中无差异表达。6条Unigenes在NR数据库匹配的物种均是巨桉,而在Swiss-Prot数据库匹配的物种有矮牵牛(, 5条)和茄子(, 1条)。
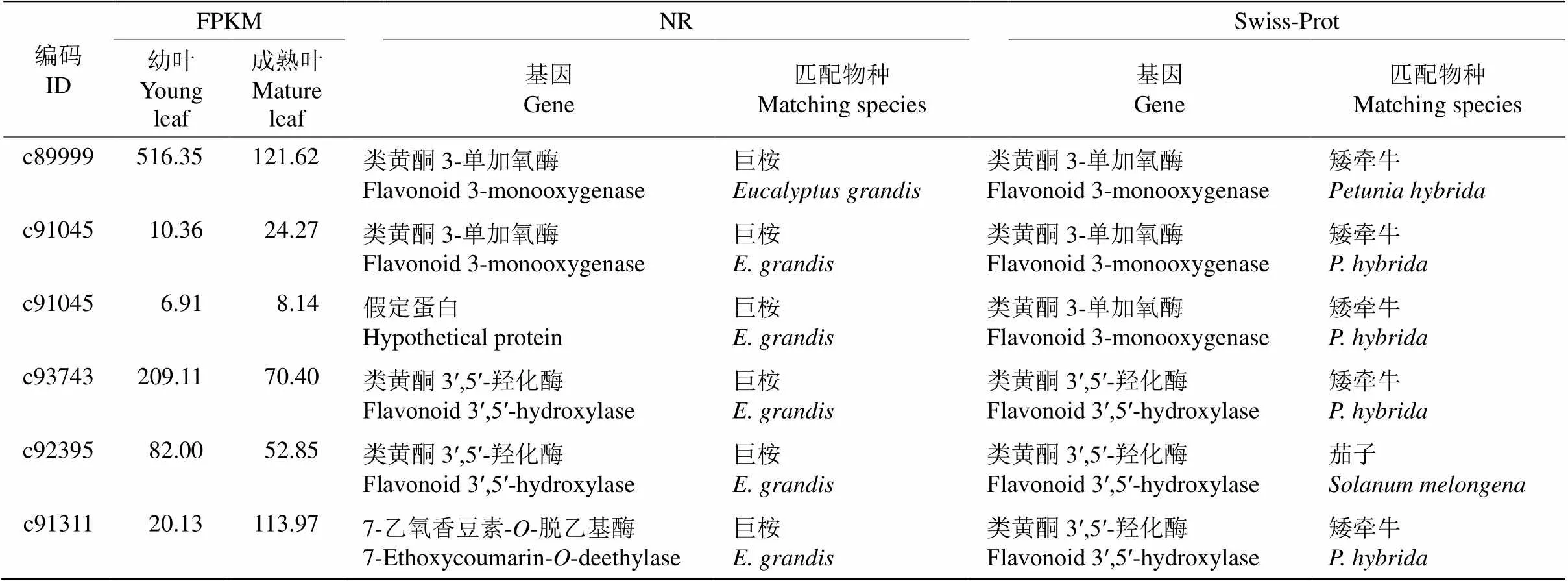
表4 6个类黄酮相关基因
2.2.3 黄酮相关基因
通过flavone (黄酮)关键词检索,依据NR及Swiss-Prot数据库分析,共获得10个黄酮相关基因(表5),包括5个、4个和1个黄酮3--酰基转移酶基因。这些基因在树葡萄幼叶和成熟叶的差异表达不显著,但这些黄酮相关基因在幼叶的FPKM值普遍低于成熟叶。这10条Unigenes在NR数据库匹配的物种主要为巨桉(7条);而在Swiss- Prot数据库中,3个基因匹配油橄榄(),基因分别匹配甘草(, 2)和苜蓿(, 2)。
2.2.4 黄烷酮相关基因
通过flavanone (黄烷酮)关键词检索,依据NR和Swiss-Prot数据库注释分析,共获得11个黄烷酮相关基因(表6),包括4个、6个2-羟基异黄烷酮脱水酶和1个黄烷酮鼠李糖转移酶基因。这些基因在树葡萄幼叶和成熟叶片的差异表达极显著下调的有2个,分别为(编码c91420)和2-羟基异黄烷酮脱水酶基因(编码c87717),其他基因差异表达不显著。这11条Unigenes在NR数据库匹配的物种主要为巨桉(9条),而在Swiss-Prot数据库匹配的物种有大豆(, 2条)、拟南芥(2条)和甘草(2条)等8种植物。
2.2.5 黄酮醇相关基因
通过flavonol (黄酮醇)关键词检索,依据NR及Swiss-Prot数据库注释分析,共获得9个基因信息(表7)。这些基因在树葡萄幼叶和成熟叶片的差异表达均不明显,除编码c82886的基因外,其他基因在幼叶的FPKM普遍低于成熟叶。同时,这9条Unigenes在NR数据库匹配的物种较多,有菌类、草本和木本植物等7种,而在Swiss-Prot数据库匹配的物种有拟南芥(4条)和水稻(, 2条)等4种植物。
2.2.6 花青素相关基因
通过anthocyanidin (花青素)和相关基因等关键词检索,依据NR及Swiss-Prot数据库注释分析, 共获得32个花青素相关基因信息(表8),包括18个花青素相关糖基转移酶、4个花青素相关酰基转移酶、5个和4个基因。其中花青素相关糖基转移酶基因包括13个和5个花青素5,3--糖基转移酶基因;花青素相关酰基转移酶包括3个花青素3--糖基6--酰基转移酶和1个花青素酰基转移酶基因。这些花青素相关基因在树葡萄幼叶和成熟叶片的差异表达极显著下调的只有编码c87867的基因。27个Unigenes在NR数据库匹配的物种为巨桉(只有4个和1个匹配为其他物种);而在Swiss-Prot数据库中,花青素相关糖基转移酶基因匹配的物种有木薯()、草莓()和拟南芥等7种植物,4个花青素相关酰基转移酶基因匹配的物种均为拟南芥, 5个匹配的物种均为葡萄(),5个匹配的物种有拟南芥(3)和银叶山蚂蝗(, 2)。

表5 10个黄酮相关基因
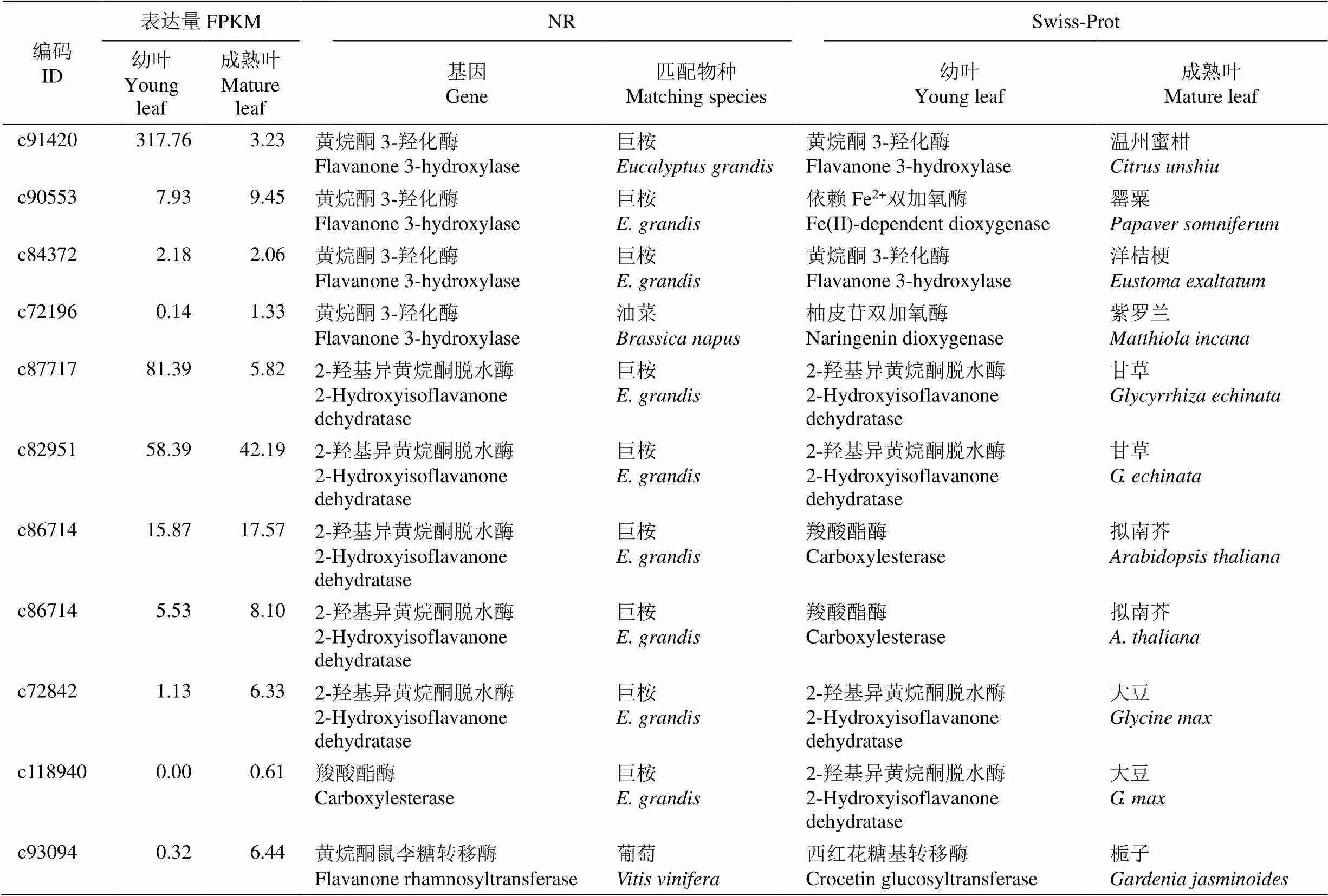
表6 11个黄烷酮相关基因

表7 9个黄酮醇相关基因
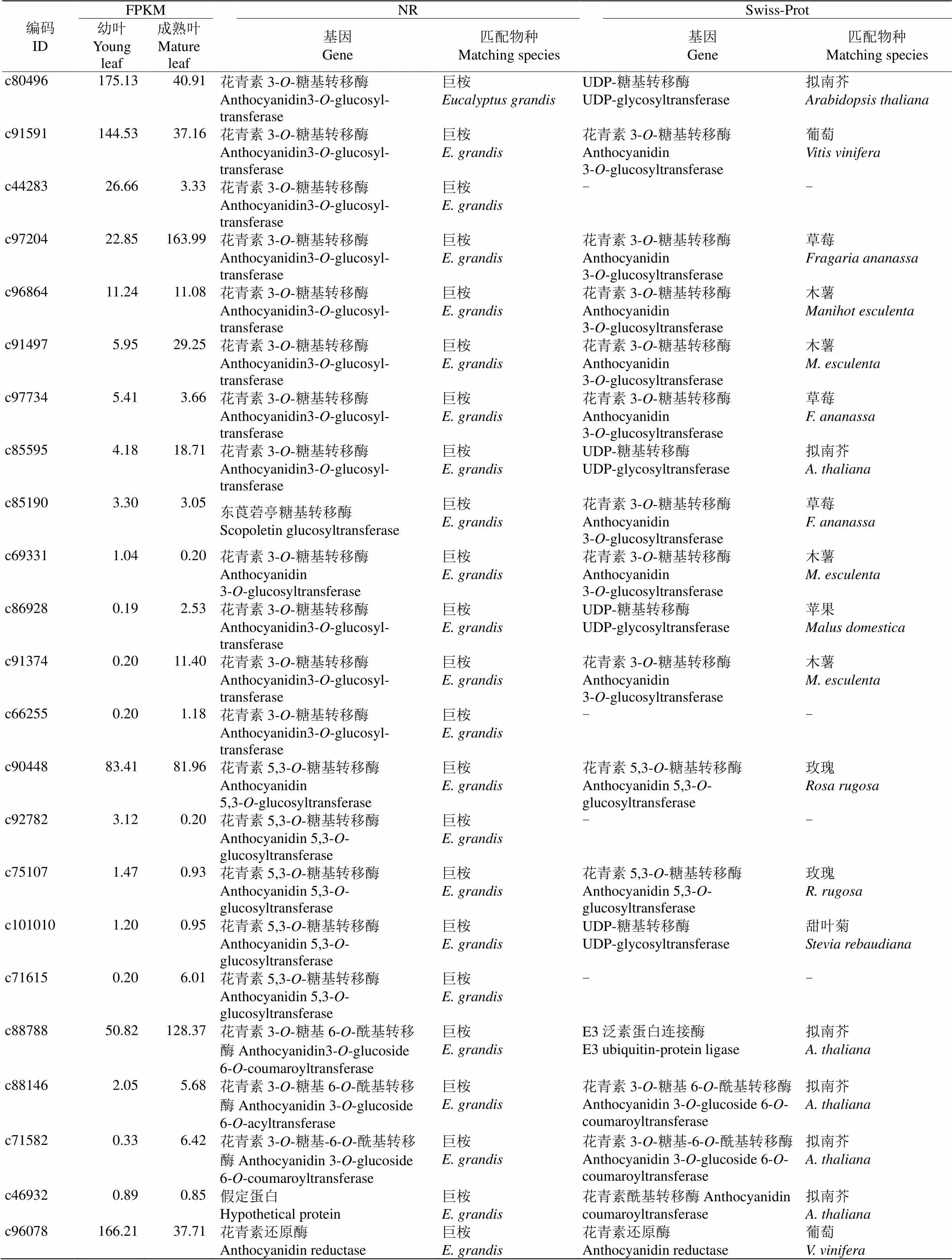
表8 32个花青素合成酶相关基因
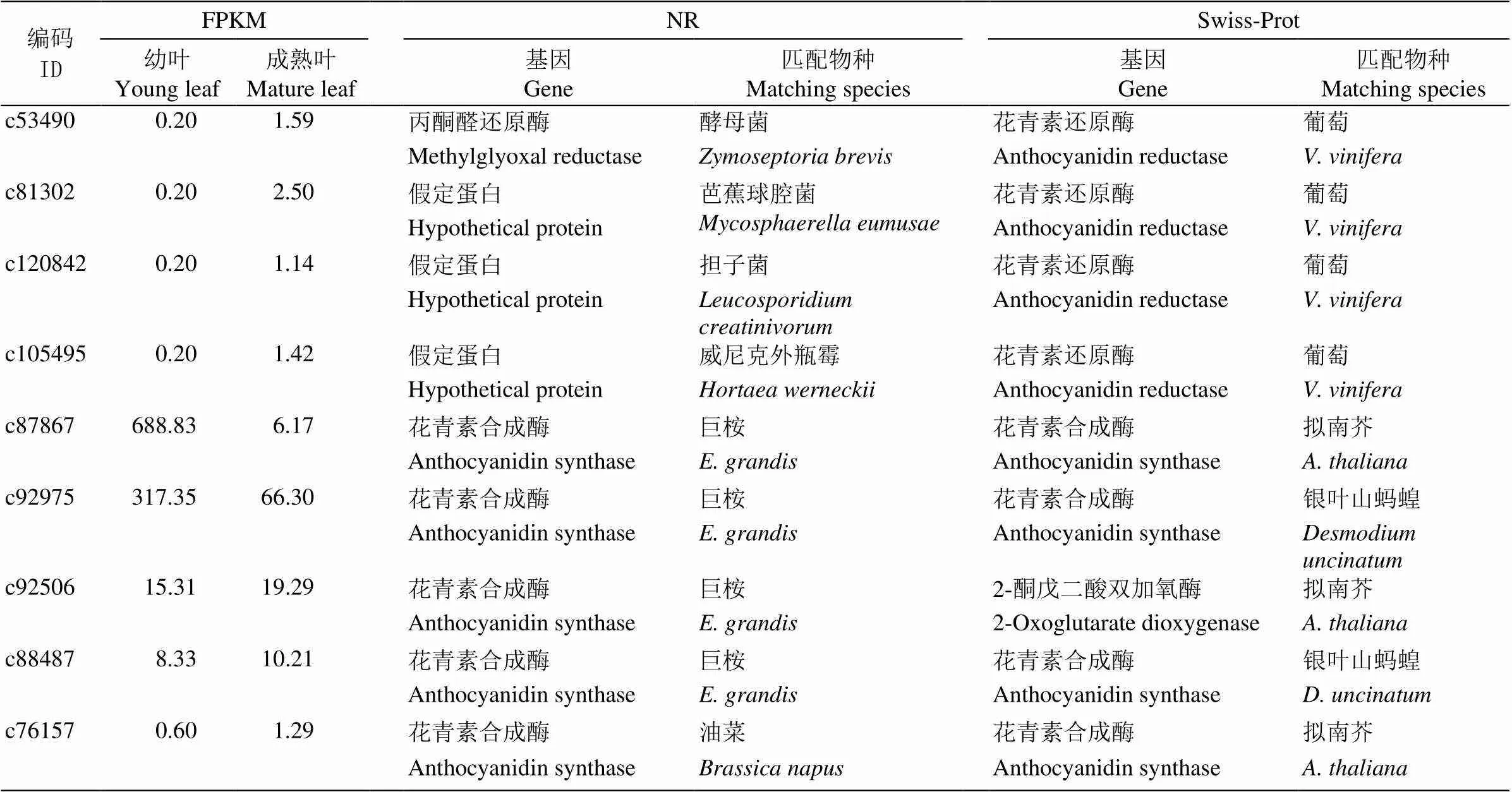
续表(Continued)
2.3 差异表达基因的qRT-PCR验证
选择差异表达显著的5个类黄酮合成酶相关基因进行qRT-PCR分析,这5个基因包括上调基因花青素3--糖基转移酶(编码C97204)以及4个下调基因,如查尔酮合成酶(编码C94118)、黄烷酮3-羟化酶(编码C91420)、2-羟基异黄酮脱水酶(编码C87717)和花青素合成酶基因(编码C87867)。结果表明(表9),上述基因在树葡萄幼叶及成熟叶片中的表达模式(上调或下调)与转录组测序结果完全吻合, 只是这两种检测结果的差异表达倍数略有不同。
3 结论和讨论
通过高通量测序分析, 从树葡萄幼叶和成熟叶片中共获得59 321条Unigenes,在8大数据库中共有32 912条Unigenes获得功能注释,NR数据库注释了32 652条Unigenes,几乎覆盖了所有其他数据库,我们选择NR数据库对树葡萄叶片类黄酮合成相关酶基因进行分析。由于NR数据库整合标准比较宽松,冗余度较高[23],而Swiss-Prot数据库中的蛋白质功能经过试验验证,注释的准确度高[23],因此,本文选择NR和Swiss-Prot数据库进行共同注释分析。这2个数据库注释到树葡萄中77个类黄酮合成酶相关基因信息,包括查尔酮、类黄酮、黄酮、黄烷酮、黄酮醇及花青素等6大类共有19种,其中查尔酮相关基因包括和;类黄酮相关基因包括和;黄酮相关基因包括、和黄酮3--酰基转移酶;黄烷酮相关基因包括、2-羟基异黄烷酮脱水酶和黄烷酮鼠李糖转移酶;黄酮醇相关基因为;花青素相关基因包括花青素相关糖基转移酶、花青素相关酰基转移酶、和。NR和Swiss-Prot数据库注释的类黄酮合成酶相关基因均为60个,NR数据库中有17条Unigenes没明确为类黄酮相关基因;Swiss-Prot数据库中有5条Unigenes未注释到信息,另有12条没明确为类黄酮相关基因。

表9 差异表达基因的qRT-PCR分析
在NR数据库注释的类黄酮合成酶相关基因中,除、黄酮3--酰基转移酶、黄烷酮鼠李糖转移酶和等基因外,其他类黄酮合成酶相关基因匹配的物种大多为巨桉,尤其是、、、2-羟基异黄烷酮脱水酶、花青素相关糖基转移酶和花青素相关酰基转移酶等家族基因所匹配的物种全是巨桉。因此,巨桉与树葡萄的大多数类黄酮合成酶相关基因亲缘关系较近。在Swiss-Prot数据库中,类黄酮合成酶相关基因匹配的物种较多,但家族成员匹配的均是矮牵牛,而家族成员匹配的均是葡萄,这表明矮牵牛的家族基因和葡萄的家族基因都进行了较为系统的研究。
通过比较树葡萄幼叶和成熟叶中类黄酮合成酶相关基因的FPKM值可知,在77个类黄酮合成代谢相关酶基因中,差异表达极显著的基因有6个,且在成熟叶中均显著下调,因此,树葡萄嫩叶中类黄酮含量显著高于成熟叶是因为嫩叶中存在高表达量的2个、1个、1个、1个2-羟基异黄酮脱水酶基因和1个。然而,类黄酮是植物重要的次生代谢产物之一[24-25],其含量除受合成代谢相关酶基因影响外[26-27],还与其转录因子调控密切相关[28]。目前,已报道关于类黄酮合成代谢的转录因子主要有MYB、bHLH和WD40等[29]。因此,本研究根据转录组测序数据,进一步分析了树葡萄叶中的上述转录因子,结果在获取的283个MYB、88个bHLH和246个WD40等三类家族基因转录因子中,差异表达极显著的分别有19、6和6个。与树葡萄叶片类黄酮合成代谢差异显著的合成酶皆为正向调控不同,这些差异表达极显著的三类转录因子中存在5个MYB、1个bHLH和1个WD40共7个负向调控因子,他们具体作用于哪个或哪些类黄酮合成酶还有待深入研究。
本研究从树葡萄类黄酮合成代谢相关酶中选取5个差异表达基因进行qRT-PCR验证,结果与转录组测序相符合。因此,本研究从树葡萄中获取的大量类黄酮合成相关不同家族基因及其在不同成熟度叶片中的差异表达情况,结合其他高等植物中存在大量家族基因[30],说明树葡萄在进化过程中经过不断复制以及串联复制等,获得了多拷贝不同家族的类黄酮合成相关基因,而这些多拷贝家族成员中起促进或抑制相关基因的表达可能集中在该家族基因中的少数基因上,大多数基因可能与树葡萄叶片类黄酮合成没有直接关系,而是否存在其他间接联系还有待进一步深入研究。本研究进行树葡萄不同成熟度叶片转录组测序,所获取的6个差异表达极显著下调基因,为进一步深入研究树葡萄或其他特色植物中类黄酮合成代谢奠定了基础。
[1] NERI-NUMA I A, SORIANO S R A, PEREIRA A P A, et al. Small Brazilian wild fruits: nutrients, bioactive compounds, health-promotion properties and commercial interest [J]. Food Res Int, 2018, 103: 345- 360. doi: 10.1016/j.foodres.2017.10.053.
[2] LIU Y J, FU J X, LIN Q H, et al. The introdution performance and cultivation techniques of jaboticaba “Sabara” [J]. S China Fruits, 2011, 40(4): 97-99. doi: 10.13938/j.issn.1007-1431.2011.04.022. 刘友接, 傅加兴, 林旗华, 等. “莎芭拉”嘉宝果引种表现及栽培技术要点 [J]. 中国南方果树, 2011, 40(4): 97-99. doi: 10.13938/j.issn. 1007-1431.2011.04.022.
[3] LIN B M, ZHENG K B, ZHANG S, et al. Studies on antioxidant and-glucosidase inhibitory activities of ethanol extracts from peels and seeds of jaboticaba at different maturities [J]. J Trop Subtrop Bot, 2018, 26(3): 233-240. doi: 10.11926/jtsb.3831.林宝妹, 郑开斌, 张帅, 等. 不同成熟度树葡萄果实醇提取物抗氧化和抑制-葡萄糖苷酶活性研究 [J]. 热带亚热带植物学报, 2018, 26 (3): 233-240. doi: 10.11926/jtsb.3831.
[4] TANG L, YUAN T T, ZHONG Q P. Analysis of nutritional components in[J]. Nonwood For Res, 2014, 32(2): 120-124. doi: 10.14067/j.cnki.1003-8981.2014.02.019.唐丽, 袁婷婷, 钟秋平. 嘉宝果营养成分分析 [J]. 经济林研究, 2014, 32(2): 120-124. doi: 10.14067/j.cnki.1003-8981.2014.02.019.
[5] QIU S L, LIN B M, HONG J M, et al. Comparative study of the anti- oxidant activity and the-glucosidase inhibitory activity of the ethanol extracts from different parts of jaboticaba plant [J]. J Fruit Sci, 2018, 35(3): 311-318. doi: 10.13925/j.cnki.gsxb.20170316.邱珊莲, 林宝妹, 洪佳敏, 等. 树葡萄植株不同部位醇提物抗氧化及抑制-葡萄糖苷酶活性的比较研究 [J]. 果树学报, 2018, 35(3) 311-318. doi: 10.13925/j.cnki.gsxb.20170316.
[6] MORALES P, BARROS L, DIAS M I, et al. Non-fermented and fermented jabuticaba (Mart.) pomaces as valuable sources of functional ingredients [J]. Food Chem, 2016, 208: 220-227. doi: 10.1016/j.foodchem.2016.04.011.
[7] PEREIRA L D, BARBOSA J M G, RIBEIRO da S A J, et al.Polyphenol and ellagitannin constituents of jabuticaba() and chemical variability at different stages of fruit development [J]. J Agric Food Chem, 2017, 65(6): 1209-1219. doi: 10.1021/acs.jafc.6b02929.
[8] SILVA R M, PEREIRA L D, VÉRAS J H, et al. Protective effect and induction of DNA repair byseed extract and pedunculagin on cyclophosphamide-induced genotoxicity [J]. Mutat Res, 2016, 810: 40-47. doi: 10.1016/j.mrgentox.2016.10.001.
[9] HACKE A C M, GRANATO D, MACIEL L G, et al. Jabuticaba () seeds: Chemical characterization and extraction of antioxidant and antimicrobial compounds [J]. J Food Sci, 2016, 81 (9): C2206-C2217. doi: 10.1111/1750-3841.13405.
[10] NIE J Y, LÜ D G, LI J, et al. A preliminary study on the flavonoids in fruits of 22 apple germplasm resources [J]. Sci Agric Sin, 2010, 43(21): 4455-4462. doi: 10.3864/j.issn.0578-1752.2010.21.015.聂继云, 吕德国, 李静, 等. 22种苹果种质资源果实类黄酮分析 [J]. 中国农业科学, 2010, 43(21): 4455-4462. doi: 10.3864/j.issn.0578- 1752.2010.21.015.
[11] CHEN J J, PENG Z X, SHI M Y, et al. Advances in on flavonoid composition and metabolism in citrus [J]. Acta Hort Sin, 2016, 43(2): 384-400. doi: 10.16420/j.issn.0513-353x.2015-0689.陈嘉景, 彭昭欣, 石梅艳, 等. 柑橘中类黄酮的组成与代谢研究进展 [J]. 园艺学报, 2016, 43(2): 384-400. doi: 10.16420/j.issn.0513- 353x.2015-0689.
[12] ZHANG K, LEI J, HE Y, et al. A flavonoids compound inhibits osteoclast differentiation by attenuating RANKL induced NFATc-1/ c-Fos induction [J]. Int Immunopharmacol, 2018, 61: 150-155. doi: 10.1016/j.intimp.2018.05.030.
[13] XU H F, WANG N, JIANG S H, et al. Content and analysis of biosynthesis-related genes of flavonoid among four strains off.F1population [J]. Sci Agric Sin, 2016, 49 (16): 3174-3187. doi: 10.3864/j.issn.0578-1752.2016.16.011.许海峰, 王楠, 姜生辉, 等. 新疆红肉苹果杂种一代4个株系类黄酮含量及其合成相关基因表达分析 [J]. 中国农业科学, 2016, 49 (16): 3174-3187. doi: 10.3864/j.issn.0578-1752.2016.16.011.
[14] Lu Q, Peng Y, Zhu C H, et al. Effect of thermal treatment on carotenoids, flavonoids and ascorbic acid in juice of orange cv. Cara [J]. Food Chem, 2018, 265: 39-48. doi: 10.1016/j.foodchem.2018.05.072.
[15] YIN L, GUAN E S, ZHANG Y B, et al. Chemical profile and anti- inflammatory activity of total flavonoids fromFisch [J]. Iran J Pharm Res, 2018, 17(2): 726-734. doi: 10.22037/IJPR. 2018.2197.
[16] SEMAAN D G, IGOLI J O, YOUNG L, et al. Dataset on the kinetics of the inhibition of PTP1B by the flavonoids and pheophytin A from[J]. Data Brief, 2018, 17: 401-406. doi: 10.1016/ j.dib.2018.01.057.
[17] BALDIN J C, MUNEKATA P E S, MICHELIN E C, et al. Effect of microencapsulated jabuticaba () extract on quality and storage stability of mortadella sausage [J]. Food Res Int, 2018, 108: 551-557. doi: 10.1016/j.foodres.2018.03.076.
[18] HSU J D, WU C C, HUNG C N, et al.extract improves diabetic nephropathy via suppression of oxidative stress and inflammation in streptozotocin-nicotinamide mice [J]. J Food Drug Anal, 2016, 24(4): 730-737. doi: 10.1016/j.jfda.2016.03.009.
[19] SERAGLIO S K T, SCHULZ M, NEHRING P, et al. Nutritional and bioactive potential of myrtaceae fruits during ripening [J]. Food Chem, 2018, 239: 649-656. doi: 10.1016/j.foodchem.2017.06.118.
[20] QIU S L, LIN B M, ZHENG K B, et al. Antioxidative activity and- glucosidase inhibitory activity of ethanol extracts of leaves of different cultivars of jaboticaba [J]. J Fruit Sci, 2017, 34(11): 1450-1457. doi: 10.13925/j.cnki.gsxb.20170141.邱珊莲, 林宝妹, 郑开斌, 等. 不同品种树葡萄叶片醇提物抗氧化及抑制-葡萄糖苷酶活性研究 [J]. 果树学报, 2017, 34(11): 1450- 1457. doi: 10.13925/j.cnki.gsxb.20170141.
[21] LIN B M, QIU S L, ZHANG S P, et al. Antioxidant capacities of different leaf extracts of Jaboticaba cultivars [J]. J NW Agric For Univ (Nat Sci), 2018, 46(9): 121-130. doi: 10.13207/j.cnki.jnwafu.2018. 09.016.林宝妹, 邱珊莲, 张少平, 等. 不同品种嘉宝果叶片提取物的抗氧化活性 [J]. 西北农林科技大学学报(自然科学版), 2018, 46(9): 121- 130. doi: 10.13207/j.cnki.jnwafu.2018.09.016.
[22] ALTSCHUL S F, MADDEN T L, SCHÄFFER A A, et al. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs [J]. Nucl Acids Res, 1997, 25(17): 3389-3402. doi: 10.1093/ nar/25.17.3389.
[23] ZHANG S P, HONG J J, QIU S L, et al. Sequencing and analysis of the transcriptome of[J]. Acta Hort Sin, 2016, 43(5): 935- 946. doi: 10.16420/j.issn.0513-353x.2016-0140.张少平, 洪建基, 邱珊莲, 等. 紫背天葵高通量转录组测序分析 [J].园艺学报, 2016, 43(5): 935-946. doi: 10.16420/j.issn.0513-353x.2016- 0140.
[24] YAO Y F, ZHANG S P, LIAN D M, et al. Transcriptome sequencing and differential expression analysis of flavonoid metabolism in flowers and fruits of okra [J]. Acta Bot Boreali-Occid Sin, 2018, 38(11): 2000- 2009. doi: 10.7606/j.issn.1000-4025.2018.11.2000.姚运法, 张少平, 练冬梅, 等. 黄秋葵花和果荚转录组测序及类黄酮代谢差异表达分析 [J]. 西北植物学报, 2018, 38(11): 2000-2009. doi: 10.7606/j.issn.1000-4025.2018.11.2000.
[25] WANG S W, FANG Y J, YU X F, et al. The flavonoid-rich fraction from rhizomes ofRoxb. ameliorates renal oxidative stress and inflammation in uric acid nephropathy rats through promoting uric acid excretion[J]. Biomed Pharmac, 2019, 111: 162-168. doi: 10. 1016/j.biopha.2018.12.050.
[26] PETRUSSA E, BRAIDOT E, ZANCANI M, et al. Plant flavonoids: Biosynthesis, transport and involvement in stress responses[J]. Int J Mol Sci, 2013, 14(7): 14950-14973. doi:10.3390/ijms140714950.
[27] DESHMUKH A B, DATIR S S, BHONDE Y, et al.root transcriptome of a medicinally important rare treefor characterization of the flavonoid biosynthesis pathway [J]. Phyto- chemistry, 2018, 156: 201-213. doi: 10.1016/j.phytochem.2018.09.013.
[28] MIERZIAK J, KOSTYN K, KULMA A. Flavonoids as important molecules of plant interactions with the environment [J]. Molecules, 2014, 19(10): 16240-16265. doi:10.3390/molecules191016240.
[29] LLOYD A, BROCKMAN A, AGUIRRE L, et al. Advances in the MYB- bHLH-WD repeat (MBW) pigment regulatory model: Addition of a WRKY factor and co-option of an anthocyanin MYB for betalain regulation [J]. Plant Cell Physiol, 2017, 58(9): 1431-1441. doi: 10.1093/ pcp/pcx075.
[30] ZHANG S P, ZHANG S H, QIU S L, et al. Thetrans- criptome as a source for anthocyanidin gene sequence analysis [J]. J Nucl Agric Sci, 2018, 32(4): 639-645. doi: 10.11869/j.issn.100-8551. 2018.04.0639.张少平, 张少华, 邱珊莲, 等. 基于转录组测序的紫背天葵花青素相关基因分析 [J]. 核农学报, 2018, 32(4): 639-645. doi: 10.11869/j. issn.100-8551.2018.04.0639.
Transcriptome Analysis of Flavonoid Synthesis Related Genes in Different Maturity Leaves of
ZHANG Shao-ping1,2, ZHENG Kai-bin1,2, HONG Jia-min1, LIN Bao-mei1, ZHANG Shuai1, QIU Shan-lian1*
(1. Subtropical Agriculture Research Institute, Fujian Academy of Agricultural Sciences,Zhangzhou 363005, Fujian, China; 2. Crop Sciences Institute, Fujian Academy of Agricultural Sciences,Fuzhou 350013, China)
The flavonoid content in immature leaves was significantly higher than that in mature leaves of. In order to understand the information of differentially expressed genes related flavonoid synthesis in, the transcriptome of 2 kinds of leaves were sequenced by Illumina HiSeq method. The results showed that there were 59 321 Unigenes obtained from young and mature leaves, among which 32 912 unigenes were annotated by 8 databases. There were 77 Unigenes related flavonoid anabolism, and 6 genes were down-regulated significantly in mature leaves, including two, one, one, one 2-hydroxyiso- flavanone dehydratase gene and one. The RT-PCR verified results of five differentially expressed genes were agreed with the transcriptome sequencing. So, it was suggested that there were a large number of family genes related flavonoid anabolism in the leaves of, but the decrease of flavonoid content in mature leaves was due to several down-regulated genes.
; Leaf;Flavonoid; Transcriptomegene
10.11926/jtsb.4040
2019-01-16
2019-03-16
福建省农业科学院创新团队项目(STIT2017-2-11); 福建省农业科学院青年创新团队项目(STIT2017-3-14); 福建省公益类科研院所专项(2017R1024-1)资助
This work was supported by the Project for Innovation Team of Fujian Academy of Agricultural Sciences (FAAS) (Grant No. STIT2017-2-11), the Project for Youth Innovation Team of FAAS (Grant No. STIT2017-3-14), and the special Project for Public Welfare Research Institutes in Fujian (Grant No.2017R1024-1)
张少平(1975~ ),男,硕士,高级农艺师,主要从事特色植物功能成分相关研究。E-mail: zspnc@163.com
Corresponding author. E-mail: slqiu79@163.com
猜你喜欢
热带作物学报(2022年7期)2022-08-06
中国农业科学(2022年14期)2022-07-26
保健与生活(2022年11期)2022-06-09
家庭医药(2021年18期)2021-12-03
保健与生活(2021年13期)2021-07-14
保健与生活(2021年5期)2021-04-12
恋爱婚姻家庭(2019年30期)2019-03-23
北方农业学报(2019年6期)2019-02-21
热带作物学报(2018年12期)2018-05-14
现代家庭·生活版(2017年10期)2017-10-20
