
中国东南部不同海拔亚热带森林中马尾松径向生长对气候的响应
2019-12-06 09:00黎敬业黄建国梁寒雪蒋少伟周鹏郭霞丽李军堂
热带亚热带植物学报 2019年6期
黎敬业, 黄建国, 梁寒雪, 蒋少伟, 周鹏, 郭霞丽, 李军堂
中国东南部不同海拔亚热带森林中马尾松径向生长对气候的响应
黎敬业1,2,3, 黄建国1,3*, 梁寒雪1,3, 蒋少伟1,2,3, 周鹏1,2,3, 郭霞丽1,2,3, 李军堂4
(1. 中国科学院华南植物园, 中国科学院退化生态系统植被恢复与管理重点实验室, 广东省应用植物学重点实验室, 广州 510650; 2. 中国科学院大学, 北京 100049; 3. 中国科学院核心植物园, 广州 510650; 4. 兴山县国有龙门河林场, 湖北 宜昌 443700)
为了解我国东南部亚热带森林不同海拔树木生长对气候响应的差异,建立了福建省武夷山脉东麓2个样点的4个马尾松()轮宽年表,对树木径向生长与气候因子进行了bootstrapped相关分析和线性混合模型(LME)拟合。结果表明,在高海拔地区马尾松径向生长对气候因子年际波动敏感性较强,主要表现为与生长季前冬季光温条件以及生长季内7月降水的正相关,生长-气候关系在不同样点间表现出较强的一致性。线性混合模型可以较好地拟合高海拔树木生长变化, 当使用前1年12月平均日最高温、当年1月日照时长和当年7月降雨量3个气候变量进行拟合时,模型解释量达到0.5,其中前1年12月最高温和当年1月日照时数在模型中起到主导作用,累积相对贡献率约占80%,说明生长季前冬季的光热条件是限制高海拔马尾松径向生长最主要的气候因子。因此,我国亚热带地区高海拔的树木径向生长可能对未来气候变化有更强的敏感性,相关森林管理政策的制定需要将此纳入考虑;同时我国亚热带地区高海拔森林中的树木有被用于树轮气候重建的潜力。
马尾松;径向生长;亚热带森林;海拔;气候;线性混合模型;中国
随着全球气候变化加剧,世界范围内的树木生长普遍受到影响[1-3]。而由于气候变化在不同区域的表现形式存在差异,且不同生境中的树木生长存在不同的气候限制因子,故树木生长对气候变化的响应也存在着多变的空间格局,探明这种生长-气候关系的空间异质性对于理解和预测森林生态系统在气候变化背景下的响应动态有着重要意义。目前该领域的研究主要沿两个梯度展开,即纬度梯度和海拔梯度。在纬度梯度方面,总体表现为高纬度地区树木生长受到生长季前期增温的显著促进[4-5], 同时受到生长季中后期高温干旱的抑制[6-9]。与之相比,生长-气候关系随海拔梯度变化的方式更为多样[10],有研究表明,欧洲堪萨斯地区和南美安第斯山脉的树木生长与温度关系表现为高海拔的正相关和低海拔的负相关[11-12],而波兰塔特拉山地区的挪威松()径向生长与温度的关系却表现为低海拔的正相关和高海拔的负相关[13]。这种海拔梯度上不均一的生长-气候关系有可能导致山地森林生态系统在气候变化中的响应模式表现出很高的空间异质性,进而可能导致森林群落结构产生垂直方向上的改变,对森林固碳潜力和物种多样性的影响意义深远。因此,我们迫切需要对此开展深入的研究,为相关地区森林管理和保育政策的制定提供参考。
在国内,大部分关于树木生长-气候关系海拔差异的研究集中在中部、西部和北部地区,东南地区未见相关报道[14-17],而中国的东南部又恰是多山地区,海拔异质性很高,南岭山脉、武夷山脉、江南丘陵和东南沿海丘陵都集中分布于此。大量山地森林生态系统孕育出了丰富的物种多样性,也是人口聚集的东南沿海城市重要的水源涵养区和生态屏障,多个国家级自然保护区和更多的省级自然保护区都划设在这一地区。因此,为了填补树木生长-气候关系海拔差异研究在我国东南部的空白,我们开展了树轮生态学试验。马尾松()作为在中国亚热带地区分布最广,且具有重要的生态和经济价值的针叶树种[18-19],对其气候响应特性进行研究有重要的理论和应用价值,又由于其树轮中早晚材间界限清晰,易于进行交叉定年工作,因此被选做该研究的目标树种。武夷山脉是我国东南部省份重要的生态屏障和水源涵养地,区内海拔落差大,气候受东亚季风和西南季风的强烈影响,适合进行树轮气候和树轮生态学试验,故而被选作研究区域。
本研究希望回答以下科学问题: (1) 研究地区内树木生长与气候要素的关系是怎样的?这一关系是否存在海拔差异?(2) 地区内同海拔相隔较远的样点间树木生长-气候关系是否存在相似性?这样的相似性是否存在海拔差异?(3) 用气候要素对树木生长进行模型拟合,拟合优度是否存在海拔差异?模型中体现出的不同海拔树木生长的主导气候因子是否随海拔变化?针对上述科学问题,我们提出以下假设: (1) 由于高海拔地区温度较低,树木径向生长可能主要受到冬春季温度的影响,表现为正相关,低海拔这一相关性不显著;(2) 相隔较远的样点间树木径向生长与气候的关系应该类似,且相似性在高海拔地区更强;(3) 用气候因子的年际波动对径向生长的年际波动进行拟合对高海拔树轮年表的拟合效果要优于低海拔年表。
1 样地概况
采样区域(图1)位于中国福建省境内的武夷山脉东麓(25.3°~26.6° N,116.8°~117.4° E),区内为亚热带季风气候,年最高月均温出现在7月,约23℃,年最低月均温出现在1月,约4℃。该区年均降水量约1 800 mm,约65%的降水量出现在1-6月(图2)。本试验设置的2个采样点WY1和WY2在武夷山脉东麓,与武夷山主脉呈平行分布,东北部的WY1样点与西南部的WY2样点相距约160 km。两样点内在高、低海拔各设置1样方(20 m×20 m),高海拔样方位于1 200~1 300 m,低海拔样方位于700~ 800 m,海拔高度差大约500 m。所有样方均在东南至西南方向的阳坡,坡度在20°以内,样方内土壤和植被类型(次生或原生常绿针阔混交林)均较为一致, 以最大化减少由于立地条件引入的系统误差。
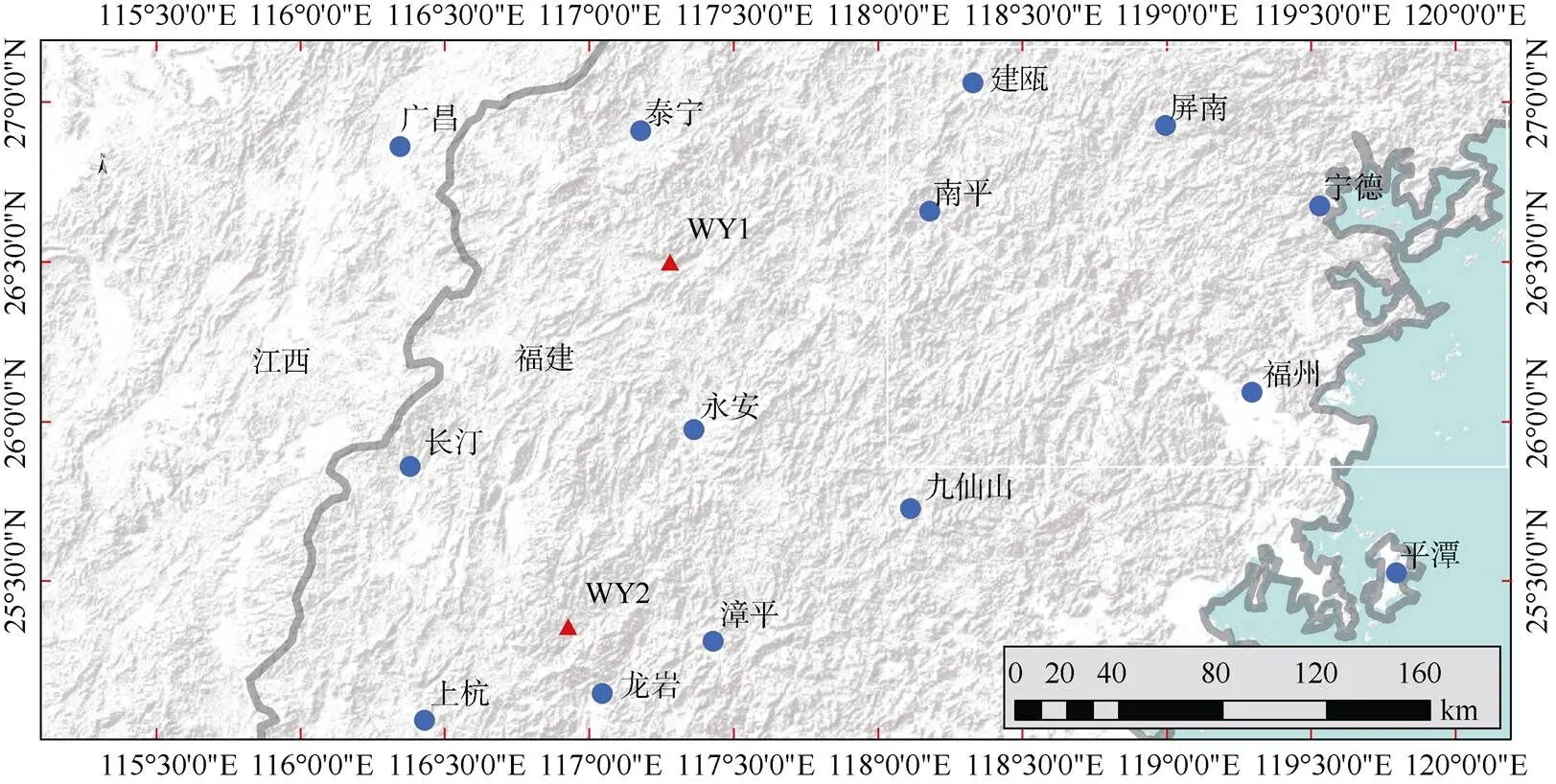
图1 采样区域和样点分布图。▲: 采样点; ●: 气象站。
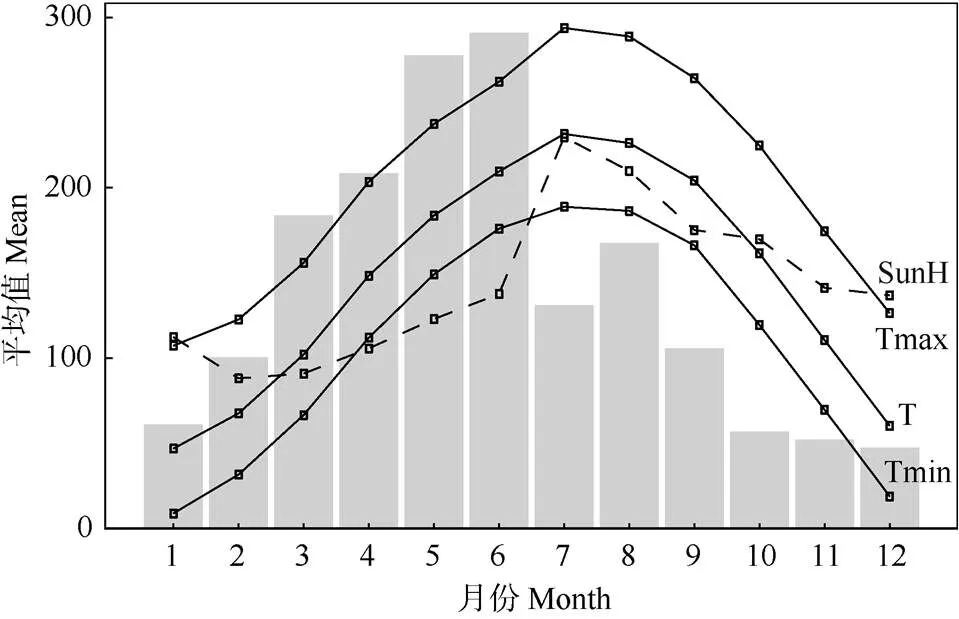
图2 采样区域气象要素1987-2014年的月均值。Tmax: 日最高温(0.1℃); T: 日均温(0.1℃); Tmin: 日最低温(0.1℃); SunH: 总日照时数(h); 柱子: 总降水量(mm)。
2 材料和方法
2.1 树轮样本处理
根据每个样方内马尾松的数量,随机选取10~20棵长势正常的马尾松()作为目标树,使用5.5 mm直径生长锥在胸径高度(约1.2~ 1.3 m)沿与坡向垂直90°方向取2根样芯。样芯随即被保存在两端封口的6 mm直径吸管中带回实验室。样芯在实验室被固定在木槽中,经过干燥、打磨等处理得到树轮清晰可辨的树轮样本。树轮样本经过交叉定年后,使用Lintab树轮测量平台和配套TSAP软件(Frank Rinntech, Heidelberg, Germany)对轮宽进行测量(精确到0.01 mm)。测量后的轮宽序列使用COFECHA软件[20]对定年质量进行检测和辅助校正。最后,每个样方内的所有轮宽序列使用ARSTAN软件[21]进行去趋势(使用50%序列长度的样条函数对轮宽序列进行标准化,从而尽可能去除树木本身年龄和其它低频信号造成的生长速度变化趋势,提高处理后轮宽年表内年际间气候波动信息的信噪比)并整合成1个标准年表(Standard chro- nology),作为后续数据分析中的轮宽指数(RWI)使用。
2.2 气候数据
本研究采用的温度、降水量和日照时数使用中国国家气候信息中心数据共享平台(http://data.cma. cn)下载的中国地面气候资料月值数据集,由于采样点距离附近气候站点远近不一,海拔差异大,且各气候站点在研究年份区间内有不同程度的数据缺失,因此本研究采用克里金插值法, 对已有站点的气候数据进行空间插值, 得到研究样点连贯和更为准确的气候数据。此外,由于水分条件不仅受到降水的影响,也与土壤性质和温度密切相关,为将上述因素纳入考虑,自适应帕尔默干旱指数(scPDSI, CRU self-calibrating PDSI 1901-2016 0.5° Global 3.25, http://climexp.knmi.nl)也被用于相关分析中(scPDSI数值没有量纲,其数值与所表征的湿润程度正相关)。
2.3 数据分析
首先,为了展现各样方内马尾松生长对气候条件年际间波动的响应模式,并对比其海拔差异, 采用R软件中的bootRes函数包计算bootstrapped相关分析[22](重采样次数=1 000),包括各样方年表与各月值气候要素(月均日最高温Tmax、月均日最低温Tmin、月总降水量PCP、月总日照时数SunH和月值自适应帕尔默干旱指数scPDSI)的相关系数。本研究中,用于计算bootstrapped相关系数。考虑到生长季前的气候条件有可能影响到生长季内的树木生长[4,23-25],本研究使用的气候数据覆盖了自生长季前一年10月至生长季当年12月的月份区间。
其次,为了评估气候条件对不同海拔马尾松径向生长速率年际波动的解释能力,采取线性混合模型分别对高低海拔年表分组(每组2个样方,高海拔为WY1-H和WY2-H,低海拔为WY1-L和WY2-L)进行拟合。相比普通多元回归,线性混合模型可以处理变量内各观测值之间不独立,不服从正态分布,或者响应变量观测值内的不同分组与对应的自变量线性关系不一致的数据。在本研究中,同一海拔分组内2个年表内的轮宽指数共同组成一个响应变量,其内部分属于不同年表的观测值与对应气候数据的线性关系(斜率)会存在差异,此时如果使用普通的线性模型进行处理,将会导致模型解释量偏低,低估气候变量对树木生长的解释能力[26-27]。线性混合模型的结构采用公式W=0+β+(0+μ)X+ε计算。
本研究构建的模型中,固定效应为气候因子,而随机效应为样方(年表)差异。W和X分别代表标准年表中样方和年份的轮宽指数以及对应样方和年份的气候数据;0和β分别代表固定效应和随机效应的截距;0和μ分别代表固定效应和随机效应的斜率;ε代表无法被固定效应和随机效应解释的随机误差。本研究采用R软件的函数包[26]进行线性混合模型的计算和分析。为了决定最终模型中应该包含哪些气候要素,我们先进行了自变量只包含单一气候要素的模型分析,然后选取解释量最高的若干变量组合纳入最终模型内。最后,为了比较不同气候要素对研究地区内不同海拔内树木生长影响的主次差异,我们对上最优模型中各变量的相对贡献率进行了分解估计[28]。
3 结果和分析
3.1 年表基本统计特征
本研究建立了4个标准年表(高海拔WY1-H, WY2-H; 低海拔WY1-L, WY2-L)。从各年表的历史折线图看,两高海拔年表间的同步性较好,存在较多重合的特征年,而两低海拔年表的同步性较差, 特征年基本不重合(图3)。对年表的公共区间(1987- 2014)分析,高海拔的年表相对低海拔表现出更高的平均敏感度(MS),说明高海拔样点内的马尾松径向生长速率(树轮宽度)年际波动中所含高频信号较多。此外,高海拔样方内各目标树轮宽序列的主成分1 (PC1)所解释的总体变异比例(平均58%)也显著高于低海拔样方(平均35%)(表1),说明高海拔样方内不同马尾松个体之间径向生长速度年际波动的同步性要显著高于低海拔样方。根据上述现象可以推论,研究区域内高海拔森林内马尾松径向生长速率可能对气候要素的年际间波动有更强的敏感性。对各年表1987-2014年的公共区间进行分析,EPS为0.84~0.89, 样本量偏少可能是EPS值偏低的原因, 大致符合树轮学年表建立质量要求(EPS>0.85)[29],是较为可信的年表数据,后续分析都将基于这一年份区间进行(表1)。
3.2 Bootstrapped相关分析
首先,对单年表与单气候要素进行bootstrapped相关分析(图4),显著相关性大部分出现在高海拔年表和气候要素的组合中,且在不同样点间呈现出良好的一致性。这验证了在研究区域内,高海拔马尾松径向生长对气候因子的敏感度强于低海拔样点, 且不同样点间马尾松径向生长在生长季内具有较为统一的气候性限制因子种类以及时空分布格局, 也侧面体现出研究区域内影响马尾松生长的生态因子在高海拔地区具有较低的空间异质性。
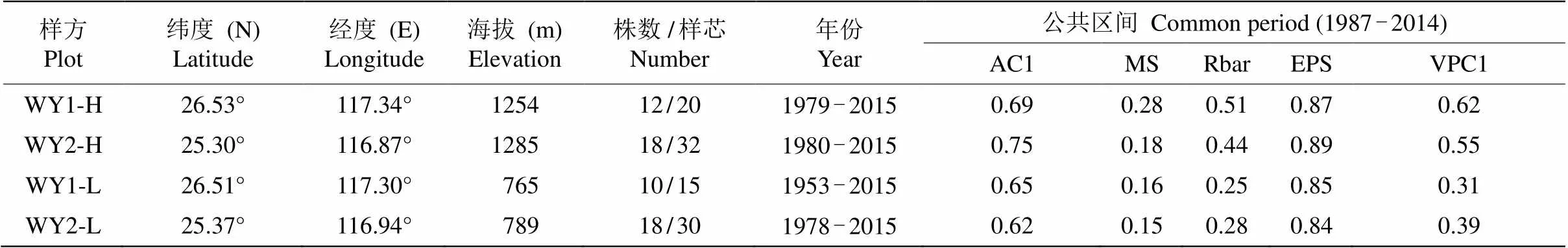
表1 样方信息和各样方STD年表统计量
AC1: 一阶自相关; MS: 平均敏感度; Rbar: 序列间平均相关系数; VPC1: 全体序列主成分1解释量。
AC1: 1st order autocorrelation; MS: Mean sensitivity; Rbar: All series correlation; EPS: Expressed population signal; VPC1: Variance in PC1 of all series.

图3 高低海拔样点的轮宽指数
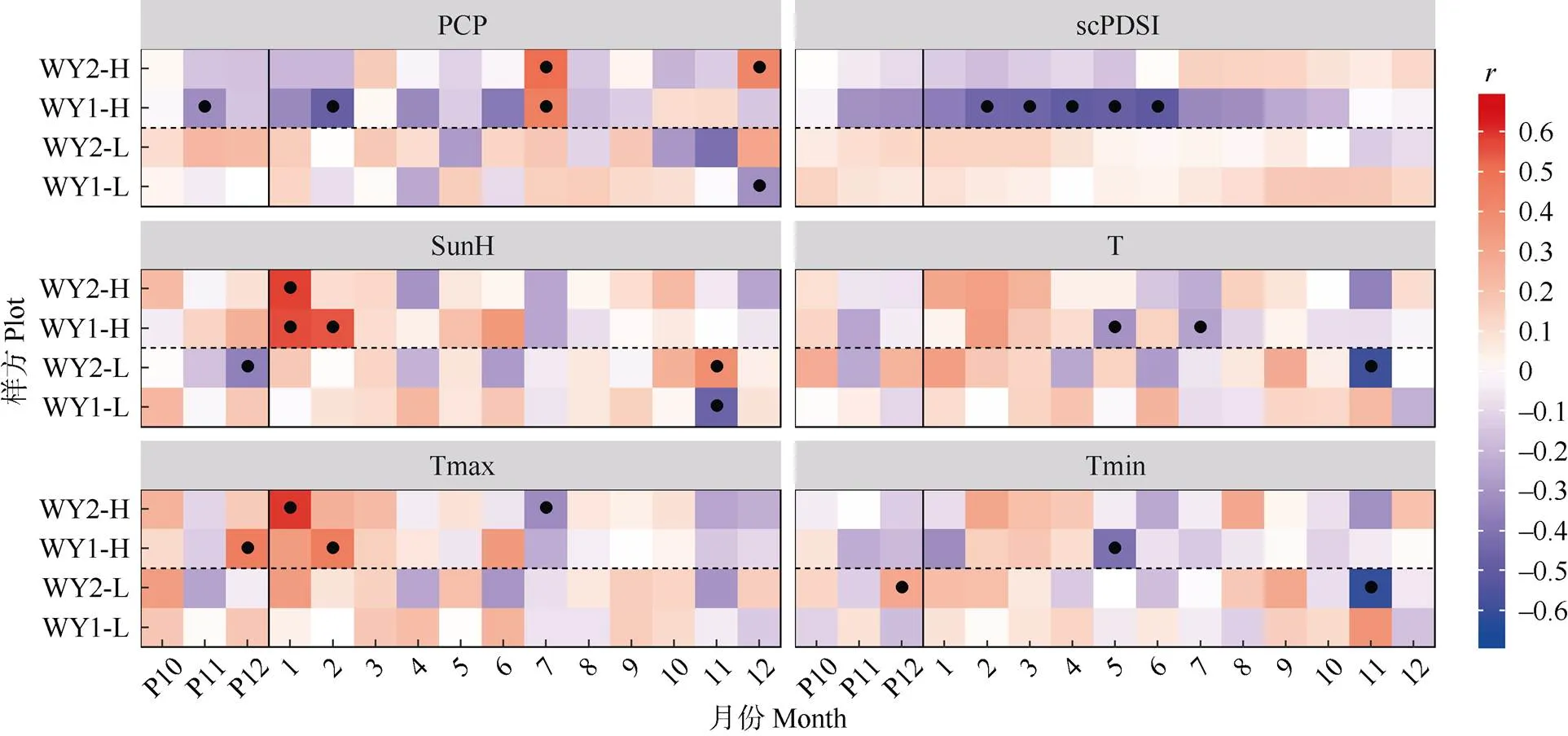
图4 标准年表与气象要素的bootstrapped相关系数。●: P<0.05。P10~P12: 前1年的10-12月。以下图表同。
具体地说,高海拔马尾松年表与生长季前1年12月至当年1-2月的温度和日照时长呈现显著的正相关(<0.05),说明生长季前冬季的光热条件对马尾松生长有着重要的影响。然而,此前有报道认为树木形成层活动起始主要由日最低温控制[30-33],而马尾松形成层开始活动的临界温度约为11℃[34]。在本研究区域内,12-2月的平均日最低温都远低于10℃,考虑到高海拔地区相对于低海拔地区由于对流层大气垂直温度直减率导致的温降,高海拔样方内的冬季温度更是远远无法满足马尾松形成层活动的要求。近年在本研究地区西南方向广东省低海拔地区开展的微树芯研究也证明,当地马尾松形成层活动在冬季进入半休眠状态,并不分裂产生木质部细胞[32]。综上可以推断,研究地区内的马尾松径向生长在冬季(12-2月)是停止的。因此,本研究中高海拔马尾松年表与冬季温度/光照呈现出的显著正相关更可能是由于较高的冬季温度和光照时长对休眠期马尾松叶片光合作用产生了促进作用。有研究表明,生长季前期树木光合作用的起始温度阈值要远低于木质部活动的起始温度阈值[35-37],樟子松()的形成层活动温度阈值大约为5℃~ 10℃, 而其光合作用在0℃以上就可以进行[38-39]。据此推测本研究地区内冬季温度很可能可以支持马尾松通过光合作用固定能量和碳,供给此后的生长季内消耗,那么在此期间的温度和日照水平的提升显然对树木生长是有利的。
此外,高海拔年表总体上在6月以前表现出对降水量和帕尔默干旱指数呈负相关,而在7月突然转变为显著的正相关。这说明,上半年内高海拔区域马尾松生长得到的水分供应是充足的。由图2可见,研究地区内年降水量的近65%都分布在1-6月。在此阶段,过多降水会减少光照,降低树木的净光合速率,且有可能导致土壤水分含量过高,造成根系缺氧,抑制树木生长。而由图2可见,研究地区内降水量从7月开始骤减超过50%,与此同时,气温在7月达到年中的最高值,马尾松生长较易面临水分亏缺和高温胁迫,从而表现出对7月降水量的正相关关系。
最后值得注意的是,低海拔年表除在生长季前1年12月以及当年11月之外,其与气候要素的相关性均不显著,且即使是在上述月份,不同样点间的生长-气候关系也无明显的一致性。这说明与高海拔地区相比,低海拔森林内的气候条件总体较为适合马尾松的生长,因此气候条件的年际间波动对低海拔树轮宽度影响不大。
3.3 线性混合模型分析
当使用单一气候因子对单一年表分组(高海拔两年表/低海拔两年表)进行拟合的情况下,对高海拔年表轮宽指数的年际波动解释量最高的气候因子是生长季当年1月的日照时长(2=0.30),其次是7月的降水量(2=0.20,<0.01)。对低海拔年表轮宽指数年际波动解释量最高的参数是生长季前一年10月的月均日最高温(2=0.06)和当年9月的月均最低温(2=0.04),但>0.05, 均不显著(表2)。当将上述解释量较高的气候要素纳入同一线性混合模型(LME)中并将变量个数限定为3个时,最优模型对高海拔年表的解释量达到了0.50,而对低海拔年表的最高解释量只有0.13 (图5)。这说明高海拔马尾松的径向生长年际变率中包含了大量的气候信息,与之相比低海拔马尾松的径向生长年际变率中的气候信息则很少。对模型内不同变量的相对贡献率分析结果表明(图6),低海拔气候-年表混合模型内3个变量的相对贡献率则没有显著差别。而在高海拔模型中,对马尾松径向生长解释量最高的气候要素是1月日照,相对贡献率超过了60%,而如果将前一年12月的月均日最高温一并考虑,则上述2个气候要素对模型的相对贡献率大约达到了80%。据此结合前述分析可以推断,研究区域内高海拔马尾松径向生长速率年际波动的主要驱动因子是生长季前冬季的光热条件,暖冬和长日照可以极大促进马尾松的生长。
4 结论
全球气候变化背景下,认识森林生态系统对气候变化响应模式的空间分布格局对更好地制定相关森林管理和生态保护政策有着重要意义。本研究首次对中国东南部亚热带森林中树木径向生长与气候条件年际波动关系的海拔差异进行了研究, 结果表明,武夷山脉东麓高海拔地区的马尾松径向生长对气候要素的年际波动有着较强的敏感性,主要受到生长季前冬季光热条件的影响,这与此前国内各地区的许多研究结论吻合[14-16,25],说明我国高海拔地区树木生长与气候关系在水平方向上异质性较低。与之相比,低海拔地区马尾松生长对气候条件年际波动的响应弱,空间差异大。本研究结果表明,武夷山脉东麓高海拔森林的马尾松生长可能会对全球变化响应更敏感,应该在未来的森林管理中给予更多关注。此外,亚热带高海拔森林内生长的树木对气候可能有较高的敏感度,具有树轮气候学研究的潜在价值。

表2 单气象要素混合模型拟合结果
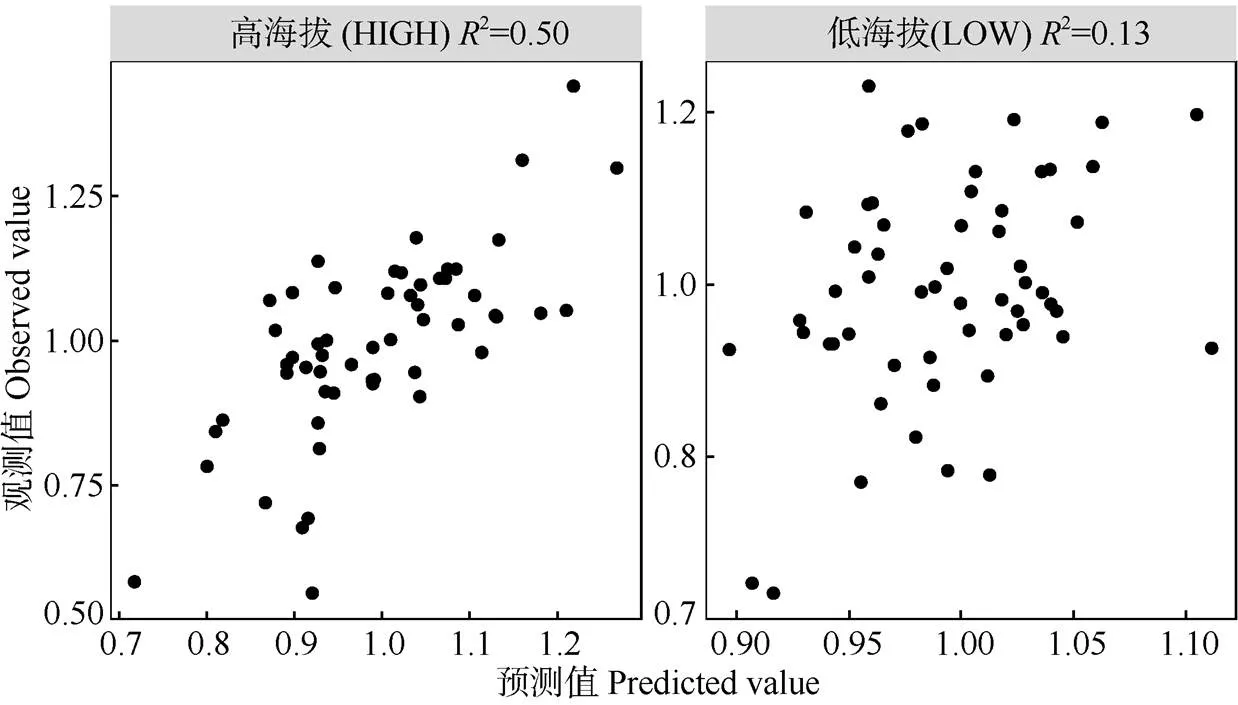
图5 对高低海拔年表进行线性混合效应模型拟合的预测值和观测值散点图
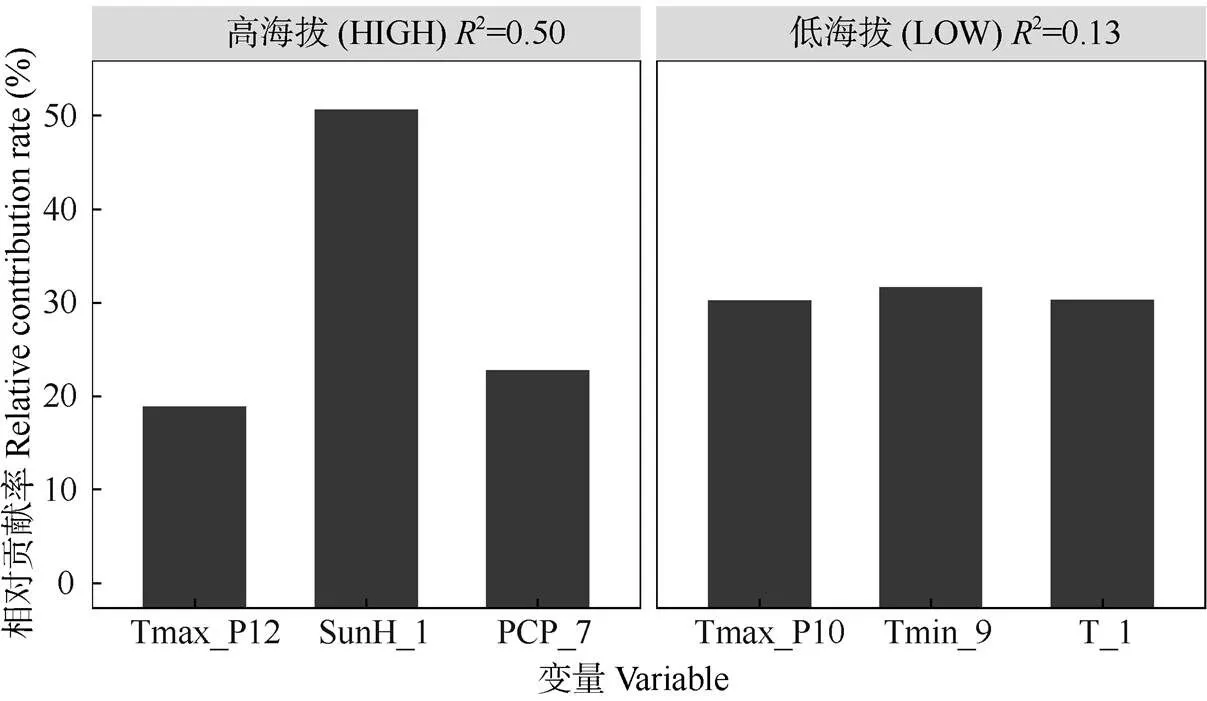
图6 线性混合效应模型LME内变量的相对贡献率
[1] YIN Y H, MA D Y, WU S H. Climate change risk to forests in China associated with warming [J]. Sci Rep, 2018, 8: 493. doi: 10.1038/s 41598-017-18798-6.
[2] PEROS M C, GAJEWSKI K, VIAU A E. Continental-scale tree popu- lation response to rapid climate change, competition and disturbance [J]. Glob Ecol Biogeogr, 2008, 17(5): 658-669. doi: 10.1111/j.1466- 8238.2008.00406.x.
[3] KHANDURI V P, SHARMA C M, SINGH S P. The effects of climate change on plant phenology [J]. Environmentalist, 2008, 28(2): 143-147. doi: 10.1007/s10669-007-9153-1.
[4] HUANG J G, TARDIF J C, BERGERON Y, et al. Radial growth response of four dominant boreal tree species to climate along a latitudinal gradient in the eastern Canadian boreal forest [J]. Glob Change Biol, 2010, 16 (2): 711-731. doi: 10.1111/j.1365-2486.2009.01990.x.
[5] JIANG X Y, HUANG J G, STADT K J, et al. Spatial climate- dependent growth response of boreal mixed wood forest in western Canada [J]. Glob Planet Change, 2016, 139: 141-150. doi: 10.1016/j. gloplacha. 2016.02.002.
[6] dos SANTOS V A H F, FERREIRA M J, RODRIGUES J V F C, et al. Causes of reduced leaf-level photosynthesis during strong El Niño drought in a Central Amazon forest [J]. Glob Change Biol, 2018, 24(9): 4266-4279. doi: 10.1111/gcb.14293.
[7] SLIK J W F. El Niño droughts and their effects on tree species compo- sition and diversity in tropical rain forests [J]. Oecologia, 2004, 141(1): 114-120. doi: 10.1007/s00442-004-1635-y.
[8] WILLIAMSON G B, LAURANCE W F, OLIVEIRA A A, et al. Ama- zonian tree mortality during the 1997 El Niño drought [J]. Conserv Biol, 2000, 14(5): 1538-1542. doi: 10.1046/j.1523-1739.2000.99298.x.
[9] NAKAGAWA M, TANAKA K, NAKASHIZUKA T, et al. Impact of severe drought associated with the 1997-1998 El Niño in a tropical forest in Sarawak [J]. J Trop Ecol, 2000, 16(3): 355-367. doi: 10.1017/ S0266467400001450.
[10] FANG K Y, GOU X H, CHEN F H, et al. The advance of dendro- ecology [J]. J Glaciol Geocryol, 2008, 30(5): 825-834. 方克艳, 勾晓华, 陈发虎, 等. 树轮生态学研究进展 [J]. 冰川冻土, 2008, 30(5): 825-834.
[11] MARTIN-BENITO D, PEDERSON N, KÖSE N, et al. Pervasive effects of drought on tree growth across a wide climatic gradient in the temperate forests of the Caucasus [J]. Glob Ecol Biogeogr, 2018, 27 (11): 1314-1325. doi: 10.1111/geb.12799.
[12] CHRISTIE D A, LARA A, BARICHIVICH J, et al. El Niño-Southern Oscillation signal in the world’s highest-elevation tree-ring chrono- logies from the Altiplano, central Andes [J]. Palaeogeogr Palaeocl Palaeoecol, 2009, 281(3/4): 309-319. doi: 10.1016/j.palaeo.2007.11.013.
[13] SAVVA Y, OLEKSYN J, REICH P B, et al. Interannual growth response of Norway spruce to climate along an altitudinal gradient in the Tatra Mountains, Poland [J]. Trees, 2006, 20(6): 735-746. doi: 10.1007/s00 468-006-0088-9.
[14] ZHANG H, SHAO X M, ZHANG Y. Research progress on the response of radial growth to climatic factors at different altitudes [J]. J Earth Environ, 2012, 3(3): 845-854. 张慧, 邵雪梅, 张永. 不同海拔高度树木径向生长对气候要素响应的研究进展 [J]. 地球环境学报, 2012, 3(3): 845-854.
[15] KANG Y X, LIU J H, DAI S F, et al. Characteristics of ring-width chronologies ofand their responses to climate change at different elevations in Taibai Mountain [J]. J NW Agric For Univ (Nat Sci), 2010, 38(12): 141-147. 康永祥, 刘婧辉, 代拴发, 等. 太白山不同海拔太白红杉年轮生长对气候变化的响应 [J]. 西北农林科技大学学报(自然科学版), 2010, 38(12): 141-147.
[16] LEI J P, XIAO W F, HUANG Z L, et al. Responses of ring width ofto the climate change at different elevations in Zigui County, Three-Gorge Reservoir area [J]. Sci Silv Sin, 2009, 45(2): 33- 39. 雷静品, 肖文发, 黄志霖, 等. 三峡库区秭归县不同海拔马尾松径向生长对气候的响应 [J]. 林业科学, 2009, 45(2): 33-39.
[17] LYU L, DENG X, ZHANG Q B. Elevation pattern in growth cohe- rency on the southeastern Tibetan Plateau [J]. PLoS One, 2016, 11(9): e0163201. doi: 10.1371/journal.pone.0163201.
[18] ZHOU Z X. Masson Pine in China [M]. Beijing: China Forestry Press, 2001. 周政贤. 中国马尾松 [M]. 北京: 中国林业出版社, 2001.
[19] HOU H Y. Vegetation of China with reference to its geographical distribution [J]. Ann Miss Bot Gard, 1983, 70(3): 509-548. doi: 10. 2307/2992085.
[20] HOLMES R L, ADAMS R K, FRITTS H C. Tree-ring chronologies of western North America: California, eastern Oregon and northern Great Basin, with procedures used in the chronology development work, including users manuals for computer programs COFECHA and ARSTAN [R]. Tucson: Laboratory of Tree-Ring Research, University of Arizona, Tucson. Chronol Ser No VI, 1986.
[21] COOK E R, HOLMES R L. Users manual for program ARSTAN [R]. Tucson: Laboratory of Tree-Ring Research, University of Arizona, Tucson, USA, 1986.
[22] ZANG C. R: The bootRes package for response and correlation function analysis Dendrochronologia [CP]. Franco: Franco Biondi Dendroclimatic Calibration, 2012. doi:10.1016/j.dendro.2012.08.001.
[23] LI D W, FANG K Y, LI Y J, et al. Climate, intrinsic water-use efficiency and tree growth over the past 150 years in humid subtropical China [J]. PLoS One, 2017, 12(2): e0172045. doi: 10.1371/journal. pone.0172045.
[24] CHEN F, YUAN Y J, YU S L, et al. Influence of climate warming and resin collection on the growth of Masson pine () in a subtropical forest, southern China [J]. Trees, 2015, 29(5): 1423-1430. doi: 10.1007/s00468-015-1222-3.
[25] LI L L, SHI J F, HOU X Y, et al. High altitudeHayata growth response to climate in Jiulongshan and Guniujiang, southeastern China [J]. Chin J Appl Ecol, 2014, 25(7): 1849-1856. 李玲玲, 史江峰, 侯鑫源, 等. 中国东南高海拔黄山松生长对气候的响应——以浙江省九龙山和安徽省牯牛降为例 [J]. 应用生态学报, 2014, 25(7): 1849-1856.
[26] BATES D M, MÄCHLER M, BOLKER B M, et al. Fitting linear mixed-effects models using lme4 [J]. J Statis Software, 2015, 67(1): 1-48. doi:10.18637/jss.v067.i01.
[27] WINTER B. Linear models and linear mixed effects models in R with linguistic applications [M/OL]. arXiv:1308.5499, 2013.
[28] Grömping U. Relative importance for linear regression in R: The Package relaimpo [J]. J Stat Software, 2006, 17(1): 1-27.
[29] FRITTS H C, SHATZ D J. Selecting and characterizing tree-ring chro- nologies for dendroclimatic analysis [J]. Tree-Ring Bull, 1975, 35: 31-40.
[30] ENSMINGER I, SCHMIDT L, LLOYD J. Soil temperature and intermittent frost modulate the rate of recovery of photosynthesis in Scots pine under simulated spring conditions [J]. New Phytol, 2008, 177(2): 428-442. doi: 10.1111/j.1469-8137.2007.02273.x.
[31] ROSSI S, ANFODILLO T, ČUFAR K, et al. Pattern of xylem phenology in conifers of cold ecosystems at the northern Hemisphere [J]. Glob Change Biol, 2016, 22(11): 3804-3813. doi: 10.1111/gcb. 13317.
[32] HUANG J G, GUO X L, ROSSI S. Intra-annual wood formation of subtropical Chinese red pine shows better growth in dry season than wet season [J]. Tree Physiol, 2018, 38(8): 1225-1236. doi: 10.1093/ treephys/tpy046.
[33] PELLERIN M, DELESTRADE A, MATHIEU G, et al. Spring tree phenology in the Alps: Effects of air temperature, altitude and local topography [J]. Eur J For Res, 2012, 131(6): 1957-1965. doi: 10.1007/ s10342-012-0646-1.
[34] ZHANG D P. A preliminary study of the relationships between the activity of stem cambium forming xylem and rainfall and temperature in[J]. J Fujian Coll For, 1994, 14(3): 215-219. 张大鹏. 马尾松茎木质部产生与水热关系的初报[J]. 福建林学院学报, 1994, 14(3): 215-219.
[35] TRANQUILLINI W, HAVRANEK W M, ECKER P. Effects of atmo- spheric humidity and acclimation temperature on the temperature response of photosynthesis in youngMill. [J]. Tree Physiol, 1986, 1(1): 37-45. doi: 10.1093/treephys/1.1.37.
[36] TANJA S, BERNINGER F, VESALA T, et al. Air temperature triggers the recovery of evergreen boreal forest photosynthesis in spring [J]. Glob Change Biol, 2003, 9(10): 1410-1426. doi: 10.1046/j.1365-2486. 2003.00597.x.
[37] OLEKSYN J, ZYTKOWIAK R, KAROLEWSKI P, et al. Genetic and environmental control of seasonal carbohydrate dynamics in trees of diversepopulations [J]. Tree Physiol, 2000, 20(12): 837-847. doi: 10.1093/treephys/20.12.837.
[38] ROSSI S, DESLAURIERS A, GRIÇAR J, et al. Critical temperatures for xylogenesis in conifers of cold climates [J]. Glob Ecol Biogeogr, 2008, 17(6): 696-707. doi: 10.1111/j.1466-8238.2008.00417.x.
[39] WU S H, JANSSON P E, KOLARI P. The role of air and soil tempe- rature in the seasonality of photosynthesis and transpiration in a boreal Scots pine ecosystem [J]. Agric For Meteorol, 2012, 156: 85-103. doi: 10.1016/j.agrformet.2012.01.006.
Elevational Heterogeneity in Radial Growth-climate Association ofin Southeastern China
LI Jing-ye1,2,3, HUANG Jian-guo1,3*, LIANG Han-xue1,3, JIANG shao-wei1,2,3, Zhou Peng1,2,3, GUO Xia-li1,2,3, LI Jun-tang4
(1. Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems,Key Laboratory of Applied Botany, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Core Botanical Gardens, Chinese Academy of Sciences,Guangzhou 510650, China; 4. State-owned Longmenghe Forestry Station of Xingshan County, Yichang 443700, Hubei, China)
To achieve a better understanding about the elevation heterogeneity in tree growth-climate association in southeastern China, four tree-ring width chronologies offrom eastern foot of Wuyi Mountains in Fujian were established, and the relationship between radial growth and climate factors along elevation were investigated by bootstrapped correlation analysis and linear mixed effect model (LME). The results showed that radial growth of trees at higher elevation had higher sensitivity to climate and higher consistency between sites, mainly reflected as the positive correlations with the winter-spring temperature/sunshine conditions and precipitation in July. Moreover, the linear mixed effect model using three monthly climate factors, such as monthly mean daily maximum temperature in last December, monthly total sunshine hours in current January, and monthly total precipitation in current July, can explain 50% of total variance of radial growth at higher elevation, and the both fronts play a leading role in the model with cumulative relative contribution rate of 80%. So, it was suggested that the pre-growing season temperature/sunshine condition are the main factor regulating tree radial growth at high elevation, and that these trees might have higher sensitivity to future climate changes and forest management policies should take this into account. Moreover, the results showed that the trees at high elevation in subtropical forest had the potential to be used for tree ring-climate reconstruction.
; Radial growth; Subtropical forest; Elevation; Climate, LME model; China
10.11926/jtsb.4038
2019-01-05
2019-02-18
国家自然科学基金项目(41861124001, 31570584, 41661144007); 中国科学院国际合作重点项目(GJHZ1752); 中国科学院百人计划项目; 广东自然科学基金项目(2016A030313152)资助
This work was supported by the National Natural Science Foundation of China (Grant No. 41861124001, 31570584, 41661144007); the International Collaborative Key Project of the Chinese Academy of Sciences (Grant No. GJHZ1752); the 100 Talents Program of the Chinese Academy of Sciences, and the Natural Science Foundation in Guangdong (Grant No. 2016A030313152).
黎敬业,硕士研究生,主要从事树木年轮学和森林生态学研究。E-mail: jingye_li@scbg.ac.cn
Corresponding author. E-mail: huangjg@scbg.ac.cn
猜你喜欢
农业灾害研究(2022年7期)2022-09-08
大气科学学报(2022年2期)2022-05-12
乡村科技(2021年17期)2021-10-20
华北水利水电大学学报(自然科学版)(2021年4期)2021-10-08
安顺学院学报(2021年4期)2021-09-16
景德镇陶瓷(2021年1期)2021-03-24
书香两岸(2020年1期)2020-07-04
延河(2018年10期)2018-10-23
中国科技纵横(2016年15期)2016-12-29
红领巾·成长(2016年4期)2016-11-19
