
九香虫水提液对锰中毒大鼠睾丸损伤的抗凋亡作用
2019-11-29 03:55付惠惠王凤月邓源林侯晓晖
遵义医科大学学报 2019年5期
姜 璘,付惠惠,王凤月,邓源林,高 飞,侯晓晖
(1.遵义医科大学 细胞生物学教研室,贵州 遵义 563099;2.中国科学院动物研究所 干细胞与生殖生物学国家重点实验室,北京 100020)
世界卫生组织(WHO)规定,不育是至少有12个月以上的不避孕性生活史而仍未受孕[1]。男性不育不是一种独立的疾病,而是由某一种或多种疾病与因素导致的后果[2]。除与男性不育直接相关的病理原因外,环境因素亦是主要原因之一,如环境污染物的暴露[3]。随着现代工业的发展,金属锰的产品越来越多地进入到我们生活中,同时采矿、精炼、干电池生产、焊接等与锰相关的行业领域均可增加人类的锰暴露水平[4]。研究表明,锰具有雄性生殖毒性,表现为损害睾丸组织,降低精子数量及活力,诱导生殖细胞凋亡,改变酶活性,干扰生殖激素水平等[5]。早期组织病理学发现,锰可致睾丸间质水肿充血,曲细精管明显受损,若染毒超过150d可见曲细精管变性,生精上皮脱离,损害随时间推移而加重,且不可逆转[6]。此外,染锰小鼠精子畸形率增高,以精子头部畸形为主,精子活动能力逐渐降低,即使低水平锰对精子仍有毒性损害作用[7]。目前,临床上应用依地酸二钠钙(CaNa2EDTA)和对氨基水杨酸钠(PAS-Na)治疗环境污染物锰所带来的生殖损伤,但长期使用EDTA可造成体内微量元素丢失等副作用[8]。因此,需要寻找更安全有效的天然产物来保护男性生殖健康。
九香虫是我国一种传统的药用昆虫,中医临床上广泛用于各种疼痛、阳痿等症状。《本草纲目》中记载:“九香虫气味咸温、无毒,主治膈脘滞气、脾肾亏损、壮元阳”。近年研究发现,九香虫提取物具有明显的抗疲劳和补肾壮阳作用[9],如何志全等利用九香虫干预锰暴露雄性大鼠的生殖器官损伤,其性行为、精子数、睾丸和附睾的脏器系数等指标均有明显改善,表明九香虫对生殖器官损伤雄性大鼠的生殖功能具有一定的保护作用[10]。虽然九香虫可用于改善环境污染物锰所导致的生殖损伤,但其保护机制尚不明确,需要进一步研究。本研究通过锰暴露建立雄性SD大鼠生殖器官损伤模型,同时利用九香虫进行干预,通过检测睾丸组织凋亡相关基因Bcl-2、Bax、CytC和C-Cas3的变化,探讨九香虫生殖保护作用的可能机制。
1 材料与方法
1.1 材料与试剂
1.1.1 实验动物 8周龄雄性SD大鼠50只,购于第三军医大学动物学实验室[许可证号:SCXK(渝)2012-0005]。大鼠给予标准饲料,自由饮水,12 h光照12 h黑暗交替,动物饲养环境温度在22~24℃,并且动物实验严格按照动物伦理委员会的规定进行。
1.1.2 实验试剂 九香虫购于贵州省遵义市百颐医药公司。兔源Bcl-2单克隆抗体购于Abcam公司;兔源Bax、CytC和C-Cas3单克隆抗体购于CST公司;HRP标记山羊抗兔试剂盒购于Proteintech公司;其它试剂均为国产分析纯。
1.1.3 九香虫水提液 首先将干燥的九香虫粉碎成末,再取0.2 kg九香虫粉末加入1L蒸馏水煮沸30 min成水煎液,后者经过滤、旋转蒸发及冷冻干燥后,最后用灭菌的双蒸水配制成相应剂量(50、100、200 mg/kg)的九香虫水提液。
1.2 实验方法
1.2.1 动物模型制备及九香虫干预 按照随机分组法,将50只8周龄雄性SD大鼠随机分成5组(n=10),即空白组(Control)、模型组(Mn)、九香虫低(50CcE+Mn)、中(100CcE+Mn)、高(200CcE+Mn)剂量干预组。除了空白组大鼠腹腔注射1ml生理盐水外,其余各组按照30 mg/kg体重腹腔注射7.5 mg/mL氯化锰溶液,连续2周、5天/周、1次/天建立生殖损伤模型;同时,空白组和模型组灌胃1 mL蒸馏水,而九香虫低、中、高剂量干预组分别灌胃不同浓度(50、100、200 mg/kg)的九香虫水提液,连续4周、7天/周、1次/天进行九香虫干预。大鼠末次给药后颈椎脱臼处死,立即取出睾丸,并检视其是否有损伤,若无损伤,将一侧睾丸保存于4%多聚甲醛溶液中固定后嵌入石蜡中,用于免疫组化及HE染色;另一侧睾丸收集于EP管中保存于液氮中,用于WB检测。
1.2.2 免疫组化检测 按常规方法将石蜡切片进行脱蜡、脱水等步骤,并参照免疫组化试剂盒说明书进行操作,一抗为Bcl-2(1∶100)、Bax(1∶50)、CytC(1∶50)和C-Cas3(1∶50),二抗为山羊抗兔IgG(1∶200)。
1.2.3 Western blot检测 按常规方法提取睾丸组织总蛋白,再进行电泳、转膜、封闭、显色等步骤,一抗为Bcl-2(1∶1 000)、Bax(1∶2 000)、CytC(1∶2 000)和C-Cas3(1∶2 000)单克隆抗体及内参GAPDH,二抗为HRP标记山羊抗兔抗体。采用 Image-Pro软件分析,扫描出灰度值。
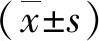
2 结果
2.1 睾丸组织HE染色 空白组睾丸曲细精管管腔大小基本一致、各级生精细胞有序排列于管壁上,着色均匀,支持细胞分布于生精细胞之间,细胞成群分布(见图1A,F)。模型组与空白组比较:生精小管管腔变大,管壁变薄,生精小管之间间隙增大,基膜不完整,各级生精细胞排列紊乱、细胞界限模糊,生精上皮的细胞层次减少(见图1B,G)。而经过九香虫干预后,睾丸组织结构得到了明显改善,生精小管管腔逐渐变小,基膜逐渐完整,生精上皮细胞层次逐渐增多(见图1C-E,H-J)。

图1 睾丸组织HE染色结果
2.2 九香虫干预影响凋亡相关蛋白表达 免疫组化检测凋亡相关蛋白BAX、BCL-2、C-CAS3和CYTC在睾丸的表达情况,如图2所示,上述蛋白在空白组大鼠各级生精细胞中均有表达(见图2A,F,K,P),而模型组较空白组的表达有所变化(图2B,G,L,Q),但九香虫干预组较模型组中各凋亡相关蛋白的表达量更接近于空白组,且随着浓度升高而变化(见图2C-E,H-J,M-O,R-T)。

图2 九香虫水提液对大鼠睾丸组织中BAX、BCL-2、C-CAS3和CYTC蛋白定位及表达的影响
Western blot检测凋亡相关蛋白BAX、BCL-2、C-CAS3和CYTC在各组大鼠睾丸中的变化情况。与空白组相比,模型组的BCL-2蛋白表达量减少,差异具有统计学意义(P<0.01);而九香虫干预组与模型组相比有一定程度的升高,尤其是中、高剂量组最为明显,差异具有统计学意义(P<0.01)(见图3)。此外,模型组的BAX、CYTC和C-CAS3蛋白表达量较空白组均有所增加,差异具有统计学意义(P<0.01);而九香虫干预组较模型组有一定程度的降低,其中BAX高剂量组以及CYTC和C-CAS3各剂量组的蛋白表达量均明显降低,差异具有统计学意义(P<0.01,见图3)。

与空白组相比## P<0.01;与模型组相比,* P<0.05,** P<0.01。图3 九香虫水提液对大鼠睾丸组织中BAX、BCL-2、CYTC和C-CAS3蛋白表达的影响
3 讨论
自然环境中锰不能被生物降解,只能在各种形态之间发生转化,其造成的环境污染很难被消除,因而其对人体产生的影响和危害已成为人们关注的热点问题[11]。过量锰在大鼠睾丸细胞内主要蓄积于可溶性组分和微粒体中,可导致细胞内微量元素分布改变及巯基含量减少,这在锰的雄性生殖毒性方面具有重要意义[12]。睾丸对锰的敏感性极高,氯化锰可通过血睾屏障、损伤生精功能及抑制精子发生[13]。张先平等[14]研究发现染锰睾丸组织中P53高表达、Bcl-2低表达以及抗氧化能力的下降可能是大鼠生精细胞凋亡增加的重要原因之一;而何志全等[15]认为九香虫对锰中毒雄性大鼠的生殖损伤具有修复作用,与提高睾丸组织和血清中SOD、T-AOC等活性有关,即抑制自由基引发的脂质过氧化作用;同时,付惠惠等[16]利用九香虫饲料对染锰大鼠睾丸损伤进行干预,发现其阻抑凋亡发生可能与调节相关基因Bcl-2和Bax的表达有关。本研究在前人研究基础上,对九香虫干预锰中毒雄性大鼠生殖损伤中的可能机制进行了深入研究,发现除了Bcl-2和Bax基因外,凋亡信号通路的其它关键基因CytC和Caspases3也发生相应改变,且这些改变对睾丸生精细胞具有明显的抗凋亡作用。因此,通过九香虫干预后可以在一定程度上改善锰中毒造成的雄性大鼠生殖损伤并具有抑制睾丸细胞凋亡的作用。
细胞凋亡存在于多细胞生物的整个生命过程当中,参与这些细胞凋亡过程的主要有4类蛋白分子,即凋亡蛋白酶(caspases)、衔接蛋白(adapter proteins)、B淋巴细胞瘤-2基因(Bcl-2)和凋亡抑制蛋白(IAPs)[17]。线粒体在细胞凋亡过程中起着重要作用,而CYTC从线粒体释放到胞浆是细胞凋亡程序关键的一步[18];同时,Bax和Bcl-2可形成异源二聚体,使凋亡受到抑制[19];半胱氨酸天冬氨酸蛋白水解酶(Caspase)家族是一组能特异性的切割天冬氨酸残基上的肽键,从而引起细胞凋亡的蛋白水解酶,是多细胞动物体内所有凋亡蛋白中最主要、最核心的死亡执行者,Caspase3是最经常被激活的,激活后经自行裂解或被动裂解为活动性小体,再重新结合成有凋亡活性的蛋白酶而发挥作用。在各种细胞的凋亡过程中,均发现有Caspase3被激活并发挥作用。Caspase3是主要的凋亡执行者,它比其它Caspase家族成员显示了更多更复杂的多样活性[20]。Cleaved-Caspase3(C-Cas3)是Caspase3活化过程中经过剪切产生的活性片段,其表达程度可反映Caspase3的活性和细胞凋亡情况[21]。本研究中各组大鼠睾丸中BCL-2、BAX 、CYTC和C-CAS3蛋白表达发生一系列改变,均提示九香虫对锰中毒引起的大鼠睾丸损伤的修复作用可能是通过调节凋亡相关基因的表达进而阻抑凋亡发生,以达到修复生殖损伤的作用。
综上所述,九香虫水提液修复锰致大鼠生殖损伤的机制可能与抑制Bax表达、提高Bcl-2表达,导致二者的异源二聚体增多有关;此外,上调Bcl-2表达可抑制CytC的表达增加,阻止线粒体凋亡信号的传递及其下游信号C-Cas3的低表达,进而抑制睾丸生精细胞凋亡。
猜你喜欢
现代医院(2022年6期)2022-08-26
中国临床医学影像杂志(2022年5期)2022-07-26
大理大学学报(2021年2期)2021-03-18
小学阅读指南·低年级版(2020年9期)2020-10-12
阅读与作文(小学低年级版)(2019年11期)2019-12-26
中国生殖健康(2019年7期)2019-01-06
大众健康(2016年1期)2016-05-26
飞碟探索(2016年5期)2016-05-10
医学研究杂志(2015年12期)2015-06-10
海外星云 (2000年18期)2000-06-12

- 遵义医科大学学报的其它文章
- 两种手术方式对单侧完全性唇腭裂患者上颌骨发育影响的对比研究
- Phylogenetic characteristics and population genetic diversity of 15 autosomal STR loci in Tujia population of Guizhou of China
- PAX1甲基化与蛋白检测在宫颈鳞癌及上皮内病变中的应用价值分析
- Effects of icariside II on hepatic cytochrome P450 expression in mice
- 过表达Nrf2对氧化应激状态下骨髓间充质干细胞凋亡的保护作用
- 鞘内注射ZD7288对糖尿病大鼠周围神经痛及血糖的作用