
基于表达谱分析陆地棉DUF642基因家族抗逆功能
2019-11-28 00:51解美霞杨君王国宁李志坤张艳孟成生马峙英王省芬
棉花学报 2019年6期
解美霞,杨君,王国宁,李志坤,张艳,孟成生,马峙英,王省芬
(教育部华北作物种质资源研究与利用重点实验室/ 河北省棉花产业协同创新中心/河北省作物种质资源重点实验室/ 河北农业大学农学院,河北保定071001)
在蛋白家族数据库Pfam 中有3 700 多种未知功能域(Domains of unknown function,DUF)蛋白,占到所有已知结构域蛋白的25%左右[1]。基因组学与蛋白质组学的快速发展为系统研究DUF家族蛋白提供了重要的生物信息学数据,更为揭示这些未知结构域家族基因在调控植物生长发育以及响应逆境胁迫方面的研究奠定了基础[2]。
DUF642 家族蛋白包含1~2 个高度保守的DUF642 结构域,是种子植物所特有的高度保守的细胞壁相关蛋白[3]。在水稻(Oryza sativa)和拟南芥(Arabidopsis thaliana)中分别有 12 个和 10个 DUF642 家族成员[4-6]。DUF642 在植物抵抗病原菌侵染和非生物逆境胁迫响应过程中发挥重要作用。拟南芥DUF642 基因 At5g25460 和At3g08030 可受细菌侵染和害虫取食诱导表达。在接种青枯雷尔氏菌(Ralstonia solanacearum)后At5g25460 下调表达[7],而 At3G08030 在接种带化红球菌(Rhodococcus fascians)后上调表达[8]。拟南芥 DUF642 家族蛋白 BIIDXI(BDX)参与调节不同组织中细胞壁果胶甲基酯化[9]。Xie 等[10]从中国葡萄(Vitis quinquangularis)丹凤 2 号中克隆到VqDUF642,其过表达能够使植株果胶甲酯酶(Pectin methylesterases,PME)活性增强,从而提高对灰葡萄孢菌(Botrytis cinerea)的抗性,表明VqDUF642 与浆果发育和抗病性有关。籽粒苋(Amaranthus hypochondriacus)AhDGR2 是 1 个编码DUF642 蛋白的基因,在盐和干旱等胁迫诱导下发生显著表达变化[11]。玉米ZmDUF642 基因在干旱、盐和铝等胁迫诱导下也能发生显著表达变化[12]。
目前,我国各大棉区都遭受不同程度的盐碱、干旱、冷害、病害、虫害的威胁[13-20]。不断发掘和筛选具有抗逆功能的基因成为棉花研究领域的重要科学问题之一。陆地棉(Gossypium hirsutum)基因组测序工作取得明显进展[21-22],使系统地鉴定和研究棉花基因家族成为可能。DUF642基因已经显示出参与植物抗逆反应的潜在重要性,但目前尚未见其在棉花中的研究报道。本研究基于陆地棉全基因组数据,对DUF642 基因家族进行了系统鉴定和生物信息学分析,特别是应用转录组数据和实时定量聚合酶链式反应(Real-time quantitative polymerase chain reaction,RT-qPCR)技术分析了DUF642 家族基因在不同组织和多种逆境胁迫下的表达规律,为后续解析其功能奠定基础。
1 材料与方法
1.1 DUF642基因鉴定与生物信息学分析
通过 TAIR(https://www.arabidopsis.org)网站下载拟南芥DUF642 家族基因序列,在陆地棉TM-1 基因组数据 (NAU version 1.1) 中进行BLAST 分析,并利用Pfam 数据库进一步鉴定。通过 ExPASy-ProtParam tool(http: //www.expasy.org/tools/protparam.html) 在线软件分析 DUF642蛋白的氨基酸残基数量、相对分子质量、理论等电点及不稳定指数等[23]。使用CELLO V.2.5(http://cello.life.nctu.edu.tw/) 在线工具进行蛋白亚细胞定位预测[24]。使用 SignalP 4.0(http://www.cbs.dtu.dk/services/SignalP/)预测信号肽[25]。
1.2 DUF642基因染色体定位和基因结构分析
从 Cotton Functional Genomics Database(CottonFGD)(https://cottonfgd.org/)[26]提取陆地棉DUF642 基因家族的基本信息,包括基因序列、编码序列(Coding sequence,CDS)、染色体位置等。利用TBtools 软件分析基因的染色体定位[27]。通过 GSDS2.0(http://gsds.cbi.pku.edu.cn/)在线软件分析基因结构[28]。
1.3 DUF642基因家族系统进化分析
利用软件MEGA7.0 对陆地棉与拟南芥DUF642 基因编码的蛋白进行多序列比对,使用相邻连接法(Neighbor-Joining,NJ)构建系统进化树[29]。
1.4 DUF642基因上游顺式作用元件分析
截取陆地棉DUF642 基因上游1 500 bp DNA 序列,利用 PlantCARE 数据库(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)预测可能存在的顺式作用元件。
1.5 转录组表达谱分析
从 NCBI SRA(Sequence Read Archive)数据库下载陆地棉的7 种器官(根、茎、叶、雌蕊、雄蕊、花萼和花瓣)和4 种逆境胁迫(冷、热、干旱和盐)处理后的转录组数据(基因组测序计划编号(Genome sequencing project accession):PRJNA 248163)。棉花材料处理和数据获取过程详见Zhang 等[21]的研究报道。以 FPKM(Fragments per kilobase of transcript per million fragments mapped)>1 作为基因表达的筛选标准。通过对数据进行log2(1+FPKM)标准化。以表达量倍数变化(Fold change,FC)>1.5 或<0.8 作为基因具有显著表达差异标准。使用HemI(Heatmap Illustrator,version 1.0) 软件绘制基因表达热图[30]。
1.6 棉苗培养、接菌与取样
供试棉花品种为抗病陆地棉农大601(ND601),由河北农业大学棉花品种创新与产业化团队提供。棉种先用无菌水浸泡24 h,然后置于湿润毛巾上于室温催芽;大约1 d 后,挑选长势一致(芽长0.5 cm)的发芽种子播种到六棱钵中,于16 h 光照/8 h 黑暗和25 ℃条件下的生长室中进行培养。棉苗生长约7 d 后用于接种黄萎病菌。参考Yang 等[31]的方法进行黄萎病菌孢子悬浮液的制备和棉苗接种处理。每棵棉苗接种悬浮液30 mL (孢子含量 107mL-1)。在接菌 0 h、6 h、12 h、24 h、48 h 时,取棉花根部,用蒸馏水清洗干净,于液氮速冻后-80 ℃保存备用。每个时间点取3株棉苗,每次取3 个生物学重复。以蒸馏水代替菌液处理的棉苗作为对照(CK)。
1.7 RT-qPCR分析
按照EASYspin Plant RNA kit 试剂盒操作说明提取棉花根组织总RNA。经1.5%(质量分数)的琼脂糖凝胶电泳和Nano Drop 2000 分光光度计检测RNA 样品的质量和浓度后,使用Rever-Tra AceqPCR RT Master Mix with gDNA Remover(TOBOYO)反转录试剂盒合成 cDNA。采用 THUNDERBIRDSYBRqPCR Mix(TOBOYO) 定量试剂盒进行 RT-qPCR。反应于7500 Real Time PCR System(Applied Biosystems)中进行,程序设置:95 ℃预变性 3 min;95 ℃变性 10 s,58 ℃退火 20 s,72 ℃延伸 34 s,40 次循环。所用引物详见表1,其中GhHis3 作为内参[32]。每个样品进行 3 次重复检测,采用 2-ΔΔCT法计算基因的相对表达量[33]。使用GraphPad Prism7 软件对数据进行Tukey 多重比较测试和绘图。

表1 本研究所用引物Table 1 Primers used in this study
2 结果与分析
2.1 陆地棉DUF642基因家族鉴定
基于陆地棉基因组数据,通过BLAST 共鉴定到23 个DUF642 基因。根据它们在染色体上的位置依次命名为GhDUF642-01~GhDUF642-23(表2)。其中GhDUF642-18 基因序列不完整,故根据华中农业大学公布的陆地棉基因组信息(HAU version 1.1) 对该基因序列进行了修正[22]。陆地棉DUF642 家族基因开放读码框(Open reading frame,ORF)长度为 612~1 245 bp,编码的蛋白含有203~414 个氨基酸残基,相对分子质量介于 22.10 ~45.15 kDa,理论等电点在4.37~9.41,不稳定指数为 27.85~41.13,结构相对稳定。除GhDUF642-11 外,家族其他成员都含有信号肽。除 GhDUF642-23 含有 1 个 DUF642结构域外,其他家族成员都含有2 个结构域。蛋白亚细胞定位预测显示,GhDUF642 中有8 个定位在细胞膜,6 个定位在胞外,5 个定位在叶绿体,2 个定位在细胞质,2 个定位在线粒体。

表2 陆地棉DUF642 基因信息Table 2 The information of Gossypium hirsutum DUF642
23 个GhDUF642 基因分布在陆地棉的14条 染 色 体 (A02、A05、A06、A07、A08、A09、A10、D03、D05、D06、D07、D08、D09、D10)和 1 条 scaffold (scaffold3920_D03) 上。GhDUF642-06 与GhDUF642-07、GhDUF642-11 与GhDUF642-12、GhDUF642-22 与GhDUF642-23 分别串联分布在A07、A10 和 D10 染色体(图1)。
2.2 GhDUF642基因结构分析
根据基因全长和CDS 序列对GhDUF642 进行基因结构分析。如图2 所示,GhDUF642 大多含有2~4 个外显子。根据内含子和外显子的数量与位置,该家族基因可分为3 组,即Ⅰ、Ⅱ和Ⅲ,分别包括10 个、6 个和7 个基因。同组内基因显示出相似的结构。
2.3 陆地棉与拟南芥DUF642家族的系统进化分析
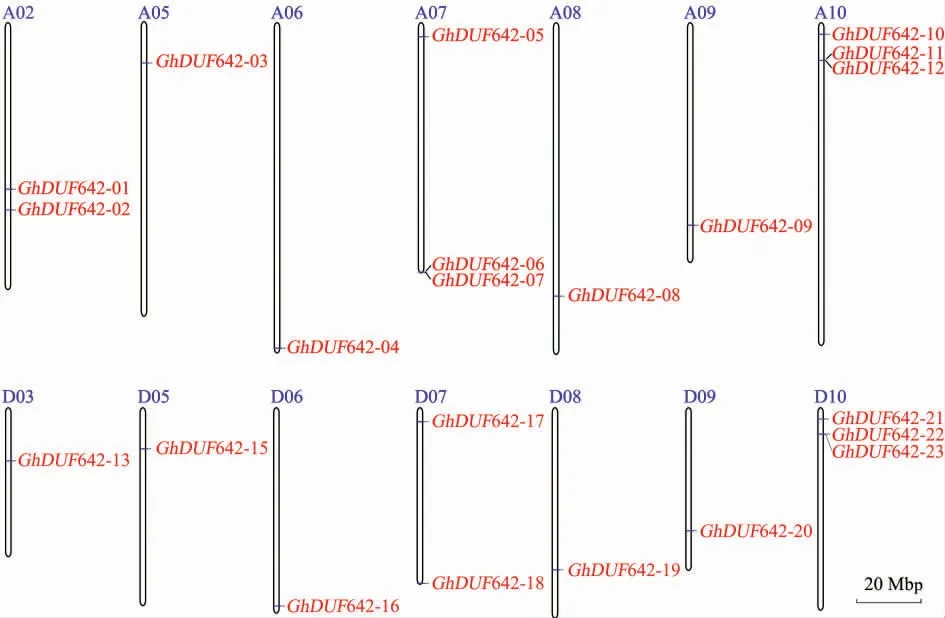
图1 陆地棉DUF642 基因的染色体定位Fig.1 Chromosome location of Gossypium hirsutum DUF642 genes

图2 GhDUF642 基因结构Fig.2 Gene structure of GhDUF642
利用陆地棉与拟南芥DUF642 家族基因编码的氨基酸序列构建系统进化树,结果(图3)显示,所有的DUF642 蛋白可分为4 个亚组。At2g41810与 GhDUF642-01、GhDUF642-09、Gh DUF642-13和 GhDUF642-20 这 4 个蛋白位于同一分支。At3g08030 与 GhDUF642-08 和 GhDUF642-19 位于同一分支。At5g11420、At5g25460 和At4g32460与 GhDUF642-02、GhDUF642-05、GhDUF642-14和GhDUF642-17 这4 个蛋白位于同一分支。每个亚组并没有因为物种差异而单独分成2 类,表明陆地棉与拟南芥DUF642 家族成员有较近的亲缘关系,可能具有相似的生物学功能。
2.4 GhDUF642基因启动子中顺式作用元件分析
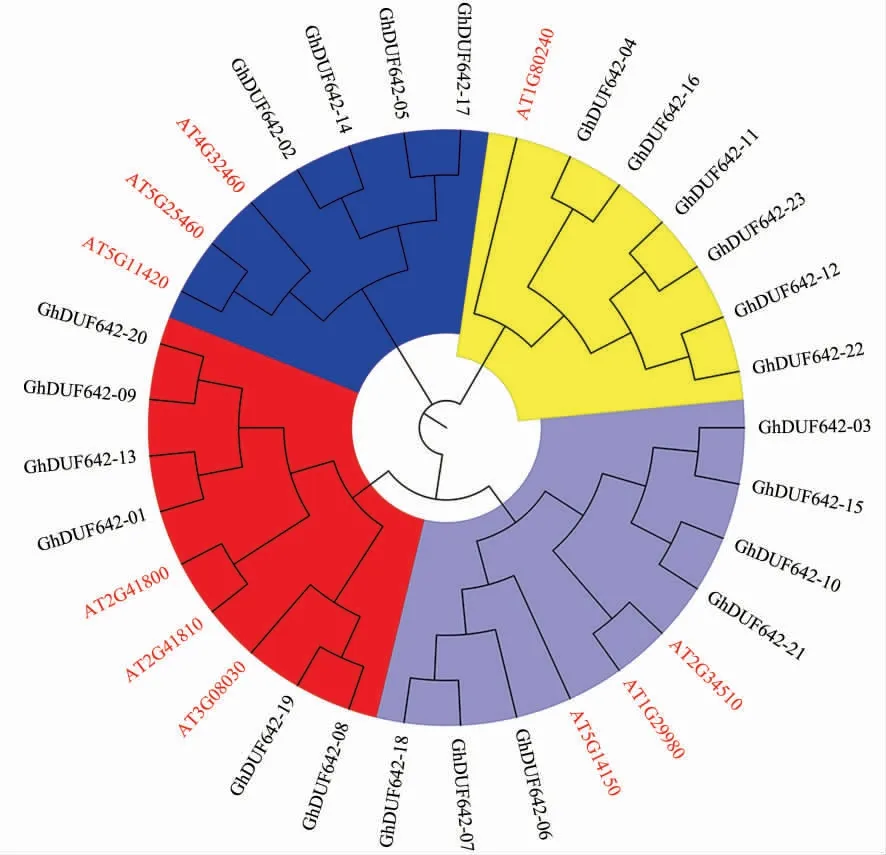
图3 陆地棉与拟南芥DUF642 家族的系统进化分析Fig.3 Phylogenetic analysis of DUF642 from Gossypium hirsutum and Arabidopsis thaliana
如图4 所示,陆地棉DUF642 家族基因上游1 500 bp 存在数量不等的响应植物激素和环境胁迫相关顺式作用元件。这些激素包括生长素(Indole-3-acetic acid,IAA)、脱落酸(Abscisic acid,ABA)、乙烯(Ethylene,ET)、水杨酸(Salicylic acid,SA)、赤霉素(Gibberellic acid,GA)和茉莉酸甲酯(Methyl jasmonate,MeJA)。环境胁迫相关的顺式作用元件有3 种,包括干旱响应元件(MBS)、防御和胁迫响应元件(Defense and stress)和低温响应元件(LTR)。其中GhDUF642-08 和GhDUF642-21 各含有9 个顺式作用元件,是具有调控元件最多的基因;GhDUF642 含有响应ABA 的顺式作用元件数量最多,如GhDUF642-01、Gh-DUF642-04 等 8 个基因各含有 3 个; 除了Gh-DUF642-01、GhDUF642-11 和GhDUF642-13 外,其余20 个DUF642 基因中都具有1 个响应ET的顺式作用元件。由此可见,每个GhDUF642 启动子含有不同数量和种类的顺式作用元件,表明它们可能通过不同的信号通路参与多种环境胁迫反应。
2.5 GhDUF642基因的组织表达特异性分析
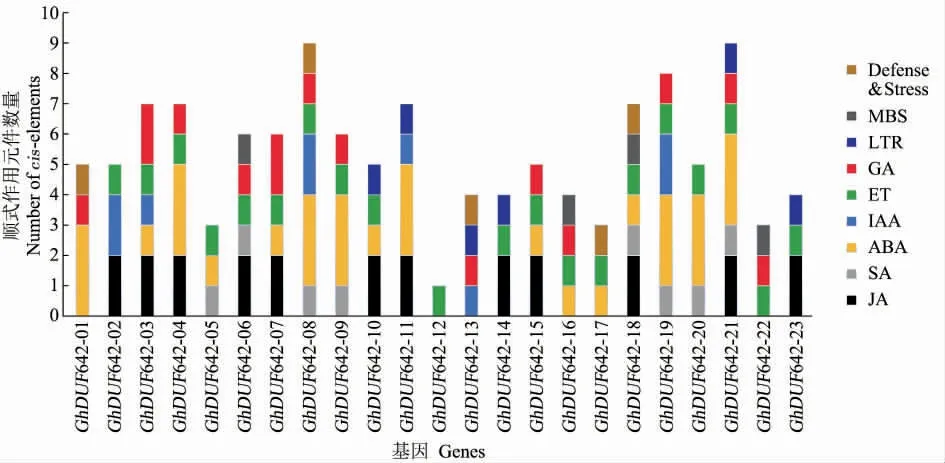
图4 GhDUF642 顺式作用元件分析Fig.4 cis-acting elements analysis of GhDUF642 genes
对GhDUF642 基因在棉花根、茎、叶、雌蕊、雄蕊、花萼和花瓣7 个组织中的表达模式分析结果显示,23 个GhDUF642 基因中有14 个具有组织表达特异性(图5)。可分为5 种组织表达模式:模式Ⅰ只包含1 个基因GhDUF642-02,其主要在副萼和根中表达;模式Ⅱ包括GhDUF642-03、Gh-DUF642-09 、GhDUF642-15 、GhDUF642-20 和GhDUF642-22,它们在7 种组织中都具有较低表达量;模式Ⅲ包括GhDUF642-08、GhDUF642-10、GhDUF642-19 和GhDUF642-21,它们在根、茎、叶和雌蕊中具有较高表达量; 模式Ⅳ包括Gh-DUF642-04、GhDUF642-14 和GhDUF642-16,它们除在茎和叶中低表达,在其余5 种组织中都高表达;模式Ⅴ只有GhDUF642-17,其主要在副萼、花瓣和根中表达。
2.6 GhDUF642基因响应逆境胁迫表达分析

图5 GhDUF642 基因的组织表达特异性Fig.5 Tissue specificity expression of GhDUF642 genes
对陆地棉DUF642 家族基因在冷、热、干旱和盐4 种逆境胁迫处理后的表达分析结果显示:冷胁迫处理后,14 个GhDUF642 基因的表达发生了显著变化,其中GhDUF642-09 在处理1 h 时显著上调表达,随后转为显著下调表达; 而Gh-DUF642-18 在处理1 h 后显著下调表达,但在处理12 h 时变为显著上调表达;GhDUF642-20 在处理6 h 时显著上调表达;其余11 个GhDUF642在冷胁迫处理后均表现为显著下调表达 (图6A)。热胁迫处理后,13 个GhDUF642 基因表达量发生显著变化,其中GhDUF642-21 在处理1 h时显著上调表达;GhDUF642-09 和GhDUF642-20在处理6 h 时显著上调表达,随后转为显著下调表达;GhDUF642-16 和GhDUF642-22 在处理 6 h时显著下调表达;GhDUF642-02 和Gh-DUF642-14 在处理 12 h 时显著下调表达 (图6B)。通过聚乙二醇(Polyethylene glycol,PEG)模拟干旱胁迫效应,导致13 个GhDUF642 基因的表达发生显著变化,其中GhDUF642-09 和Gh-DUF642-16 在处理1 h 时显著上调表达,但Gh-DUF642-16 在处理12 h 时又显著下调表达;Gh-DUF642-22 在处理1 h 后先显著下调表达,而在处理12 h 时变为显著上调表达;GhDUF642-10和GhDUF642-20 在处理 3 h 和6 h 时分别发生显著下调表达;GhDUF642-02、GhDUF642-14 和GhDUF642-17 在处理12 h 时显著下调表达 (图6C)。盐胁迫处理后,12 个GhDUF642 基因的表达量发生了显著变化,其中GhDUF642-09 和Gh-DUF642-16 在处理1 h 时显著上调表达,随后转为显著下调表达; 其余10 个基因在盐胁迫处理后部分时间点表现为显著下调表达(图6D)。这些GhDUF642 对于不同胁迫处理表现出不同的表达变化,表明它们可能在棉花应对不同的逆境胁迫时发挥不同的重要作用。
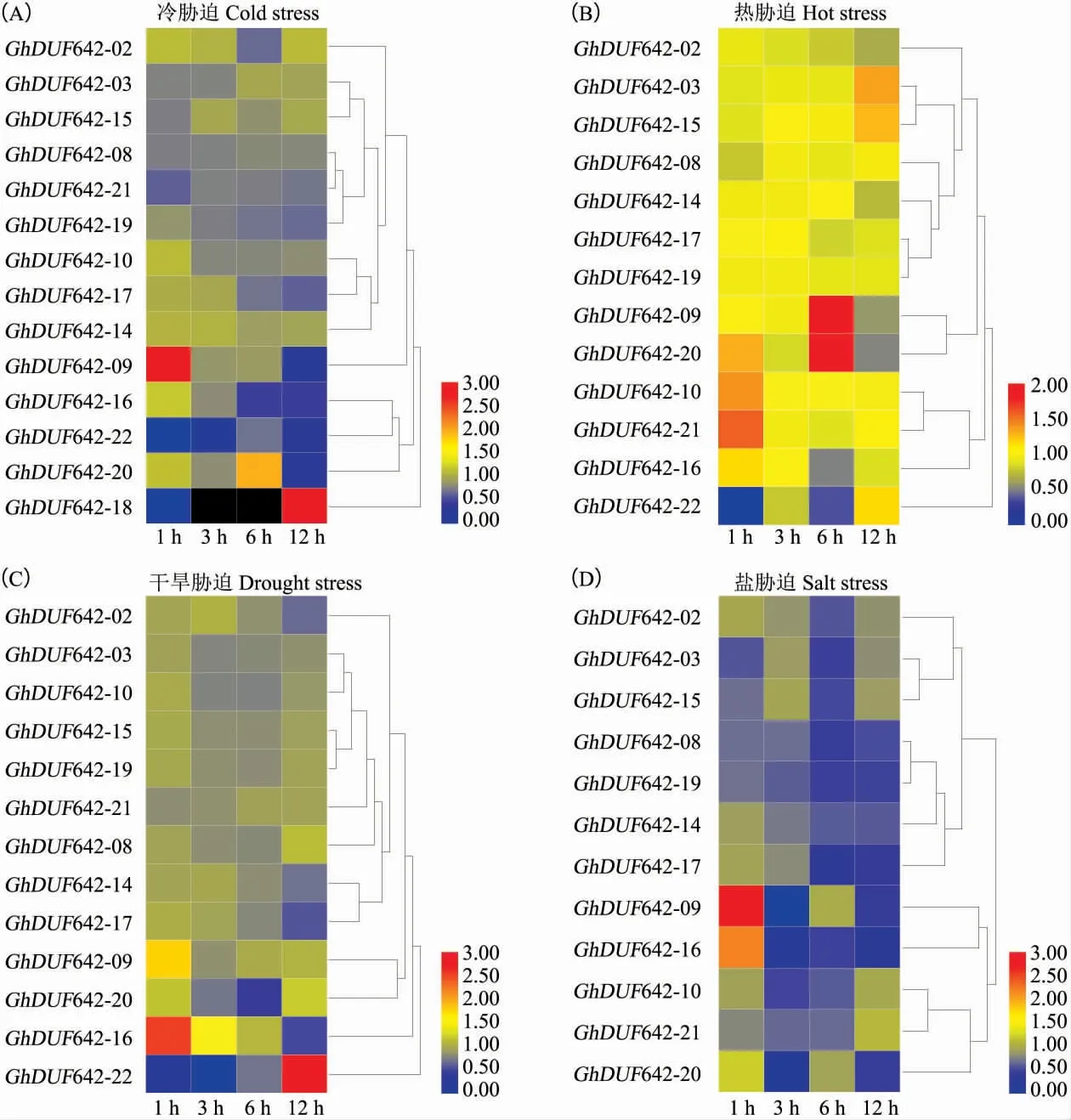
图6 陆地棉 DUF642 基因在冷(A)、热(B)、干旱(C)和盐(D)等 4 种逆境胁迫下的表达Fig.6 Expression profiling of DUF642 genes from Gossypium hirsutum under four stresses, including cold (A), hot(B), drought (C) and salt (D)
2.7 黄萎病菌胁迫处理后GhDUF642基因的表达分析
根据本研究团队已获得的大丽轮枝菌胁迫处理后的棉花根组织转录组数据(尚未发表),选择其中6 个具有潜在表达差异的GhDUF642 基因进行RT-qPCR 分析。结果显示,在一段时期内,GhDUF642-04、GhDUF642-08、GhDUF642-16、GhDUF642-19 和GhDUF642-21 这 5 个基因受黄萎病菌诱导后显著下调表达,而Gh-DUF642-10 显著上调表达(图7),表明这些基因参与棉花响应黄萎病菌侵染过程。
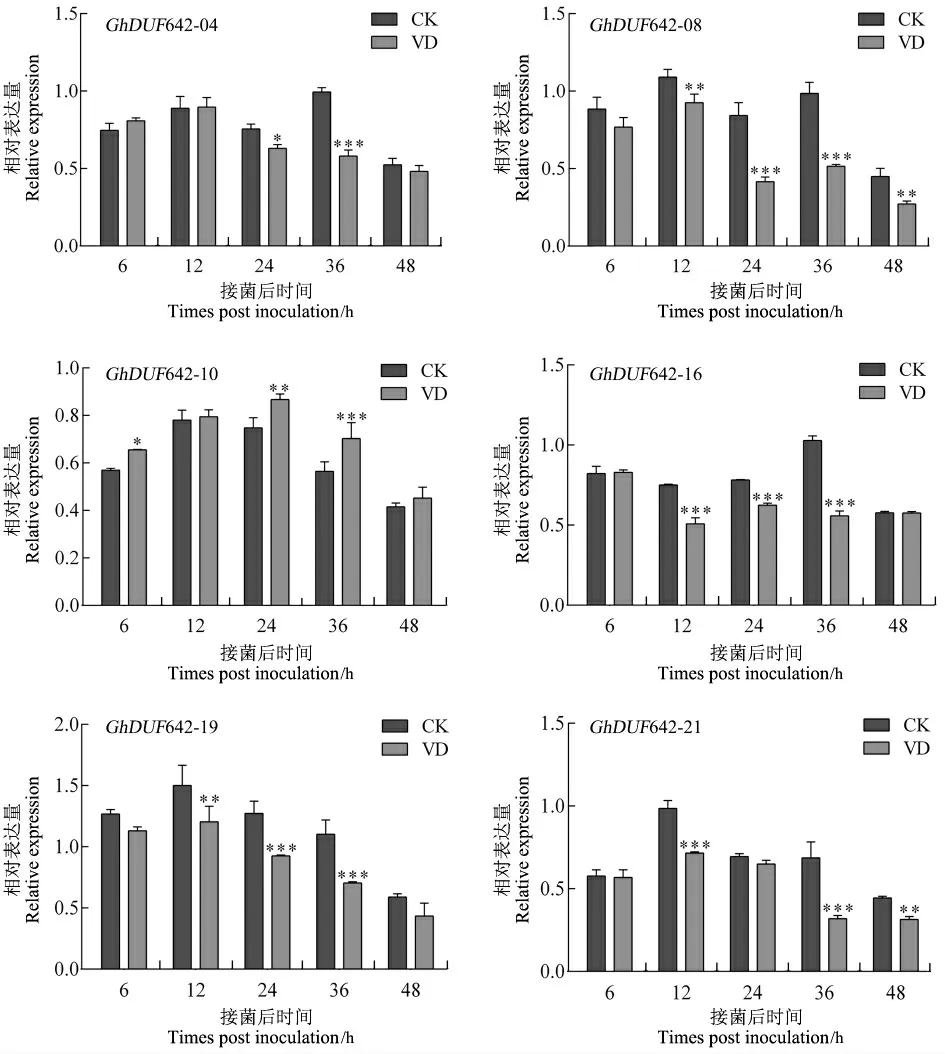
图7 黄萎病菌胁迫下陆地棉DUF642 基因表达分析Fig.7 The expression analysis of GhDUF642 genes from upland cotton induced with Verticillium dahliae
3 讨论与结论
近年来,棉花基因组测序工作的不断完成与更新,为从全基因组水平研究基因功能奠定了基础。目前,关于棉花DUF642 基因家族的研究还未见报道。本研究首次在陆地棉基因组中鉴定到23 个GhDUF642 基因(表 1),多于拟南芥(10个)和水稻(12 个)中已鉴定的DUF642 基因数量[4-6],这可能是由于棉花基因组加倍的结果。Gh-DUF642-06 与GhDUF642-07、GhDUF642-11 与GhDUF642-12 以及GhDUF642-22 与GhDUF642-23 分别串联于 A07、A10 和 D10 染色体 (图1),可能是由于基因串联复制导致该基因家族的扩张。基因结构分析显示GhDUF642 家族基因可分为3 组,各组基因具有相似的基因结构,且内含子长度、外显子数目几乎一致,但各组之间内含子序列长度不同(图2)。另外,通过氨基酸序列进行的聚类分析显示,拟南芥与棉花的DUF642基因发生了聚集效应(图3)。GhDUF642-01、GhDUF642-09、GhDUF642-13、GhDUF642-20 与At2g41810 位于同一进化分支,而At2g41810 与拟南芥耐盐相关[34],因此推测这4 个GhDUF642 基因可能参与棉花盐胁迫响应。转录水平上的变化进一步表明了GhDUF642-09 参与棉花抗盐胁迫(图6D)。GhDUF642-08、GhDUF642-19 与At3g08030 位于同一进化分支。目前,At3g08030被证明参与种子发育过程[35],尚未见其参与抗逆功能的报道。本研究发现GhDUF642-08 和Gh-DUF642-19 序列上游都有响应 SA、ABA、IAA、ET 和GA 的调控元件(图4),并且它们在根中高表达(图5),受黄萎病菌诱导后显著下调表达(图7),表明这2 个基因可能参与棉花抗黄萎病菌侵染过程,但具体的抗病信号通路还有待进一步确认。At4g32460 和 At5g11420 编码的 DUF642 蛋白在拟南芥发育过程中可能是PME 活性的阳性调控因子。在种子萌发过程中过表达这2 个基因,转基因株系的PME 活性显著增强,种子萌发性能改善[36]。At5g25460 对根与叶的伸展和生长具有重要的作用[4]。GhDUF642-02、GhDUF642-05、GhDUF642-14 、GhDUF642-17 与 At4g32460 、At5g11420 和At5g25460 位于同一进化分支 (图3),推测这4 个GhDUF642 基因可能参与调节棉花生长发育过程。目前,关于植物DUF642 基因家族功能的研究还很有限。本研究通过生物信息学鉴定了陆地棉DUF642 基因家族,在转录水平上初步探讨了它们可能具有的生长发育和抗逆功能,为进一步研究该家族基因奠定了重要的基础。
猜你喜欢
作物学报(2022年9期)2022-07-18
中国特种设备安全(2022年1期)2022-04-26
农业科技与信息(2021年4期)2021-12-05
新农村(浙江)(2021年2期)2021-11-30
中国棉花(2021年11期)2021-03-05
园艺与种苗(2020年12期)2021-01-08
中学生数理化·中考版(2016年7期)2016-12-07
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
汽车维修与保养(2015年8期)2015-04-17
