
圈养大熊猫初生幼仔声音通讯行为研究
2019-11-22 07:39:14刘璇薛飞郭俊良吴蔚侯蓉周江齐敦武张志和
四川动物 2019年6期
刘璇,薛飞,郭俊良,3,吴蔚,侯蓉,周江,齐敦武*,张志和
(1.贵州师范大学生命科学学院,贵阳550001;2.成都大熊猫繁育研究基地,四川省濒危野生动物保护生物学省部共建实验室,成都610081;3.西华师范大学生命科学学院,珍稀动植物研究所,四川南充637002)
大熊猫Ailuropodamelanoleuca是我国特有的珍稀濒危动物之一,母兽在繁殖期和育幼期的主要通讯方式是声音(潘载扬,2015;魏辅文,2018),而大熊猫幼仔在初生时的视觉、听觉和嗅觉尚未健全,且不具备运动能力(胡锦矗,2001;张志和,魏辅文,2006),因此声音信号是新生幼仔与母兽识别和联系的重要途径(Phillips &Stirling,2000;Collinsetal.,2006)。大熊猫幼仔刚出生时就可以发出非常洪亮的叫声(黄祥明等,2001;Baoticetal.,2014),且不同情境下的叫声可以反映其觉醒程度(Stoegeretal.,2012),从而传达自身某些特定状态的信号(Baoticetal.,2014)。本研究假设大熊猫幼仔在需求不同时发出的声音信号亦不相同,且当幼仔通过叫声表达不同需求时,母兽也会用不同行为进行回应。在分析大熊猫幼仔与母兽交流声音特征的基础上,探讨两者在声音信号发出后的行为互应关系,进一步探究圈养大熊猫的育幼行为,为人工育幼提供科学依据和技术支持。
1 实验地点和研究方法
1.1 实验地点和研究对象
研究对象来自成都大熊猫繁育研究基地的6只幼仔和4只母兽(表1)。于2017年7—9月和2018年5—8月对幼仔从出生之日起到日龄45 d这一阶段的声音和行为资料进行持续收集。实验期间,当幼仔饥饿时,饲养员将幼仔还给母兽,待幼仔吃奶完成后,饲养员将幼仔由母兽处取出,立即放于恒温的育幼箱,在育幼箱中观察对象可以自由活动。因本次实验目的在于探究幼仔和母兽的声音通讯,所以只采集将幼仔还给母兽这一时间段的声音数据。
1.2 数据采集
用Shure VP89M指向性麦克风(频率范围:20 Hz~20 kHz,+0.9/-4 dB)和Tascam DR-100MKII录音机(频率范围:10 Hz~40 kHz,+0.1/-0.5 dB)采集声音,并使用SONY FDR-AXP55摄像机同步摄像。声音录制时观察母兽行为,将幼仔发出叫声时的情境分为2种:一是当幼仔发出叫声时,母兽回应;二是当幼仔发出叫声时,母兽不回应。
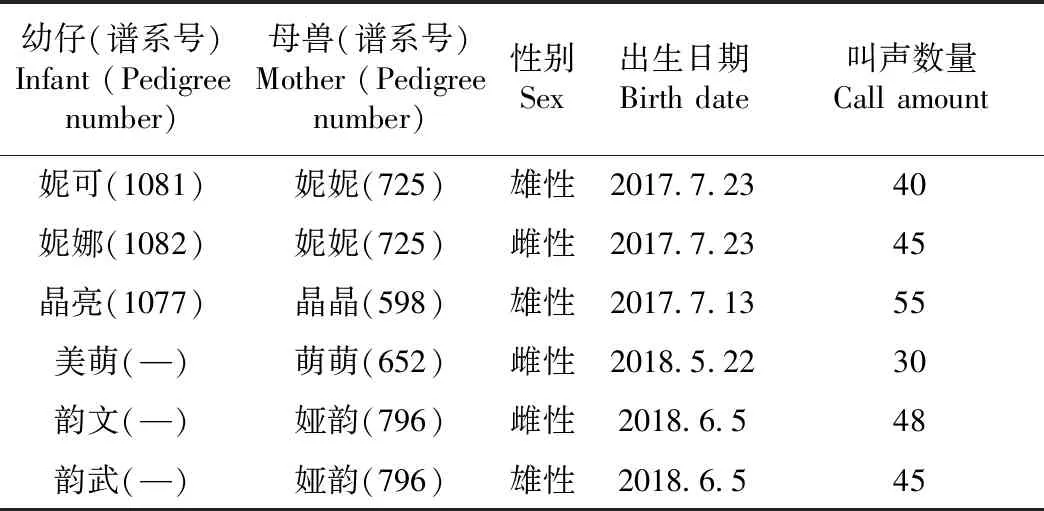
表1 母兽及幼仔的个体信息Table 1 The individual information of mother and infant giant pandas
注:— 2018年幼仔的谱系号尚未公布
Note:— pedigree numbers of the infant born in 2018 have not been published
根据听觉分辨、肉眼对声音窄带语图和结构的的观察等,对声音类型进行区分。使用Adobe audition 3.0对声音数据进行分析,声音参数包括基频平均数(meanf0)、主频平均数(dominant frequency)、时长(duration)和谐波数(number of harmonics)。
1.3 行为观测和行为谱
采用焦点动物取样法和连续记录法观察和记录母兽行为。
参照胡锦矗等(1985)、刘定震等(2002)、Snyder等(2003)、张志和和魏辅文(2006)对圈养大熊猫行为的定义,以及何永果等(2015)对大熊猫育幼期行为的研究结果对大熊猫母兽在育幼期的行为进行分类(表2)。
1.4 数据分析
对所有用于分析的声音,使用主成分分析法对提取出的参数进行概述,然后将获得的主成分得分值输入判别分析中进行评价,从而将叫声类型分成不同的组别。判别分析的结果使用声音分类正确的百分数表示。对分辨出的声音分类后,将对声音有回应的母兽行为进行分类,累计各个行为的频次并计算该种行为在所有行为频次中所占的百分比;然后通过卡方检验对不同叫声下的行为差异性进行比较。所有数据均使用SPSS 22分析,显著性水平设置为α=0.05。

表2 大熊猫母兽育幼期的行为类型和描述Table 2 Type and description of mother giant panda behaviors during the breeding stage
2 结果
2.1 声音分类结果
幼仔声音被分成3种类型,Squawk:基频值较高(820.53 Hz±160.45 Hz)、主频值较高(931.98 Hz±435.12 Hz)、时长最短(290.48 ms±148.70 ms)、谐波数最多(11.33±2.61),当幼仔发出Squawk叫声时,嘴巴是张开的;Squall:基频值最高(1 160.96 Hz±224.99 Hz)、主频值最高(2 239.94 Hz±742.73 Hz)、时长较长(294.31 ms±99.67 ms)、谐波数较多(10.65±2.27),当幼仔发出Squall叫声时,嘴巴是张开的;Croak:基频值最低(728.74 Hz±120.34 Hz)、主频值最低(735.92 Hz±138.48 Hz),但时长最长(447.74 ms±167.49 ms)、谐波数最少(1.82±0.74),当幼仔发出Croak叫声时,嘴巴是闭合或轻微张开的(图1)。

图1 大熊猫初生幼仔3种叫声的时域和频域特征Fig.1 Time-domain and frequency-domain of the 3 call types of infant giant pandasA.Squawk,B.Squall,C.Croak
主成分分析法将基频、主频、时长和谐波数提取主成分因子(PC1:53.5%,PC2:26.5%,PC3:12.8%,PC4:7.2%),其中,PC1和PC2能够代表声音中80%的变量(表3),因此,选择PC1和PC2对声音进行分类分析。其中,PC1与基频、主频和时长参数显著相关(χ2=228.43,df=31,P<0.01),PC2与谐波数显著相关(χ2=203.83,df=17,P<0.01);利用主成分分析法获得叫声散点图(图2)。声音类型判别函数揭示出声音类型之间的差异有统计学意义(n=263,Wilks’λ=0.144,χ2=503.663,df=4,P<0.01),263个声音数据中有235个被正确分类,正确率达到89.4%。

表3 大熊猫初生幼仔各声音变量的主成分分析Table 3 Principal component analysis of the voice variables of infant giant pandas

图2 利用主成分分析法获得的叫声散点图Fig.2 Scatterplot of the different call types by principal component analysis
2.2 母兽对幼仔叫声的行为回应
263个声音数据中,Squawk叫声共90个,母兽回应的有52个(57.8%);Squall叫声共95个,母兽回应的有64个(67.4%);Croak叫声共78个,母兽回应的有46个(59.0%),表明母兽对幼仔不同的叫声的回应比例有差异,但差异无统计学意义。
对于幼仔发出的3种叫声,母兽回应的行为类型以及所占的比例不同:在幼仔发出Squawk叫声时,母兽回应的行为包括舔仔(37.0%)、叼仔(44.2%)、哺乳(5.7%)、调姿(9.6%)、抱仔(3.8%)5种行为;在Squall叫声的刺激下,母兽回应的行为有舔仔(37.5%)、叼仔(36.0%)、哺乳(15.6%)、调姿(4.6%)、调整位置(6.2%)5种行为;在听到幼仔的Croak叫声后,母兽则是用舔仔(67.3%)、叼仔(6.5%)、抱仔(6.5%)、调姿(15.0%)、调整位置(4.3%)等5种行为进行回应。同一种回应行为在不同叫声的刺激下的比例不同:当幼仔发出Squawk和Squall叫声时,母兽舔仔行为比例之间的差异无统计学意义(χ2=0.11,df=1,P>0.05),而在Croak叫声的刺激下,母兽舔仔行为的比例显著高于Squawk(χ2=9.29,df=1,P<0.05)和Squall(χ2=9.57,df=1,P<0.05);在听到Croak叫声时,母兽的叼仔行为显著低于Squawk(χ2=17.81,df=1,P<0.05)和Squall(χ2=12.83,df=1,P<0.05),而Squawk和Squall之间的差异无统计学意义(χ2=0.825,df=1,P>0.05);哺乳、调整位置、抱仔和调姿等行为在不同类型叫声刺激下的差异均无统计学意义(图3)。
3 讨论
本研究将大熊猫幼仔的叫声分为3个类型,与Baotic等(2014)划分类型一致,但本文中Croak叫声的基频参数(728.74 Hz±120.34 Hz)与之(36.89 Hz±15.06 Hz)差异较大。Riede和Brown(2013)明确指出动物叫声的基频与身体大小成反比,大熊猫初生幼仔个体仅有近百克,该体质量对应的叫声基频为1 000 Hz,因此,本研究的Croak叫声的基频参数应当是可信的,上述差异推测与2个研究中的录音设备以及环境不同有关。

图3 大熊猫幼仔3种叫声类型下母兽回应行为占比Fig.3 Proportions of the mother behavior responding to the 3 call types of infant giant pandas* P<0.05
声音通讯是动物交流信息的重要手段(孙儒泳,2001),不同参数特征的声音信号能够传达发声个体当前的状态和需求等信息(Charltonetal.,2009,2010;Cuietal.,2010)。相比于其他通讯方式,声音通讯具有传播距离远、不容易受到遮蔽障碍物屏蔽以及发声个体不需要进行非常复杂的行为等特征(孙儒泳,2001;潘载扬,2015),这种通讯方式非常适合初生时发育程度十分低下、缺乏视力和行动能力的大熊猫幼仔(胡锦矗,2001)。
育幼期是动物生长发育和种群延续的关键时期,而育幼行为则是能够保障下一代健康成长的关键。藏酋猴Macacathibetana幼仔体温下降或感到不适时会发出哭叫声,进而引起母兽的注意和帮助(Blumberg &Sokoloff,2001);黑猩猩Pantroglodytes幼仔遇到危险时,会发出尖叫声向母兽寻求帮助(Levréro &Mathevon,2013)。根据声音信号参数、结构的不同,大熊猫幼仔叫声可以划分为Squawk、Squall、Croak 3种类型(Baoticetal.,2014)。对任意一种叫声,母兽的动作回应都有舔仔、叼仔,而其他育幼行为则只出现在1种或2种叫声中。这表明舔仔、叼仔这2种行为可能是母兽用于解决幼仔需求的最优先行为。当幼仔发出Squawk和Squall叫声后,母兽主要的回应方式是通过叼仔、抱仔和调姿等行为让幼仔能够更舒适地待在自己身上;同时当幼仔发出Squawk、Squall叫声,母兽做出的同一种行为反应在比例上并不具有显著的差异性,因此我们认为Squawk和Squall在本质上是具有相同功能的叫声类型,通过高频叫声来给予母兽强烈的刺激,从而使母兽能够在短时间内满足自身的迫切需求。这种声音能够向母兽传达的信息可能会相对单一,但能够使母兽尽快进行行为回应。而当幼仔发出Croak叫声时,母兽则通过舔仔和调姿进行回应。同时相对于其他2种叫声,Croak叫声得到的行为回应有显著的差异,这表明Croak叫声是独立的一种叫声类型,幼仔可能更多使用这种叫声来表达出一些并不迫切的需求,而母兽会通过舔舐幼仔安抚并进一步确认幼仔的状态,以便做出下一步的行为回应。因此,我们认为大熊猫幼仔不同的叫声类型是具有明确的功能意义差别,能够引导母兽对其作出正确的行为回应从而更好地抚育幼仔。
虽然声音通讯在大熊猫育幼过程中起到了重要作用,但它并不是大熊猫个体之间的主要交流方式(Nieetal.,2012;魏辅文,2018)。大熊猫幼仔眼睛在出生40 d左右会睁开(黄祥明等,2001),视力逐渐清晰,对外界的感知能力会增强,运动能力也会随着成长不断增强。在黑猩猩中,随着幼仔成长,母幼之间的交流方式逐渐从声音通讯向其他通讯方式过渡(Levréro &Mathevon,2013)。因此,推测大熊猫母幼之间的通讯方式也会随着幼仔的发育产生变化,从而更好地适应外界环境以提高生存几率。虽然随着幼仔的成长,声音通讯的作用会逐渐减弱,但在个体发育前期,通过分析幼仔发出叫声的类型,能够快速对幼仔的状态和需求进行初步判断,这在一定程度上能够减少人工育幼过程中对幼仔不同状况的反应时间,从而提高个体的生存几率和生存质量。
猜你喜欢
中国人民公安大学学报(自然科学版)(2022年1期)2022-07-20 02:51:14
四川畜牧兽医(2022年5期)2022-06-30 10:48:16
野生动物学报(2022年2期)2022-05-16 10:26:30
新疆大学学报(自然科学版)(中英文)(2022年2期)2022-03-27 02:08:08
四川动物(2022年1期)2022-01-26 07:43:56
小天使·五年级语数英综合(2021年5期)2021-07-11 11:10:31
野生动物学报(2020年4期)2020-10-27 02:41:58
山东交通科技(2020年2期)2020-08-13 09:24:06
电子制作(2017年20期)2017-04-26 06:57:35
中国畜牧兽医文摘(2015年9期)2015-12-29 03:38:11
