
滇龙胆草7-脱氧番木鳖酸-7-羟化酶基因的克隆与表达分析
2019-11-13 03:39张晓东李彩霞王元忠
江苏农业科学 2019年17期
张晓东 李彩霞 王元忠

摘要:克隆滇龙胆7-脱氧番木鳖酸-7-羟化酶基因GrDL7H,并进行表达分析,为龙胆苦苷生物合成途径的解析奠定基础。根据滇龙胆转录组GrDL7H序列,使用RT-PCR(逆转录PCR)和3′RACE(cDNA末端快速扩增)技术从滇龙胆幼叶中克隆该基因及其启动子,并进行序列分析和组织特异性表达分析。结果表明,GrDL7H基因(登录号:KT306971)全长2 154 bp,包含5个外显子和4个内含子,其中GrDL7H基因(登录号:KP340980)开放阅读框长 1 542 bp,编码513个氨基酸;GrDL7H蛋白质相对分子质量为59.08 ku,等电点(pI值)为8.88,属于CYP450蛋白超家族成员,可能定位于叶绿体;GrDL7H蛋白质无信号肽,为亲水稳定蛋白质,主要由α-螺旋和环构成;GrDL7H蛋白质具有CYP450蛋白质保守结构域,功能为参与氧化还原反应;滇龙胆GrDL7H蛋白质与黄金鸡纳树CcDL7H蛋白质的亲缘关系最近;GrDL7H基因启动子主要包含6个光应答元件、1个赤霉素应答元件、6个参与茉莉酸甲酯应答的顺式调控元件和1个热胁迫应答元件等;组织特异性表达分析结果表明,GrDL7H基因主要在叶片中表达。
关键词:滇龙胆;7-脱氧番木鳖酸-7-羟化酶;基因克隆;表达分析
中图分类号: S188;Q78 文献标志码: A 文章编号:1002-1302(2019)17-0076-06
滇龙胆(Gentiana rigescens Franch. ex Hemsl.)为多年生草本植物,主要分布于云南、四川等地,其根部主要用来治疗肝炎和胆囊炎[1-2]。目前,由于滇龙胆的市场需求量逐年增加,野生滇龙胆遭到人为大肆破坏[3]。从合成生物学角度来看,滇龙胆的主要药效成分为龙胆苦苷,要大量合成龙胆苦苷,首先必须先弄清龙胆苦苷的生物合成途径及其调控机制。
萜类生物合成途径包括基本前体形成、碳骨架结构形成和萜类化合物的结构后修饰3个阶段[4]。7-脱氧番木鳖 酸-7-羟化酶(CrDL7H)是萜类龙胆苦苷生物合成途径第二阶段重要的酶,能够催化7-脱氧番木鳖酸生成番木鳖酸[5]。在长春花中,有研究者通过病毒诱导的基因沉默技术鉴定了 7- 脱氧番木鳖酸-7-羟化酶,CrDL7H基因沉默后,裂环番木鳖苷含量减少了70%,7-脱氧番木鳖酸鲜质量含量增加到4 mg/g,且在裂环番木鳖苷途径中羟基化先于羧基氧甲基化[5]。使用表达CrDL7H酶的酵母微体进行酶动力学分析,结果表明其米氏常数Km和最大速率Vmax分别为(111.07±14.80) μmol/min和(5.50±0.77) μmol/min[6]。目前,CrDL7H酶基因已从长春花、小曼长春花、萝芙木、秀丽藤、金银花、金鸡纳树、水甘草等许多植物中分离[5],但是目前仅在长春花中进行过详细的研究。DL7H基因表达具有组织特异性。在川西獐牙菜中,SmDL7H基因在叶中的表达量最高,其次是莖和花,在根中的表达量最低;并且与G10H、IS、IO、SLS2等基因表达量相比,DL7H基因的表达量相对较低[7]。在长春花中,CrDL7H基因主要在叶中表达,在第1对叶中表达量最高,在第2、3、4、5对叶中的表达量逐渐降低,且对应的长春碱与裂环番木鳖苷的含量逐渐减少;其次是茎和花,表达量最低的为根[8]。
目前,由于龙胆科植物的基因组并未测序,导致龙胆苦苷生物合成途径并不完全清楚[9]。本研究根据笔者于2013年在滇龙胆转录组数据库中检索的GrDL7H基因序列,设计特异性引物,通过逆转录PCR(RT-PCR)技术成功从滇龙胆叶片中扩增到GrDL7H基因,然后根据所克隆基因序列设计引物,对该基因的启动子序列进行扩增和分析,最后对二年生滇龙胆根、茎、叶、花等不同组织中GrDL7H基因的表达情况进行分析,以期为滇龙胆龙胆苦苷生物合成途径的解析提供帮助。
1 材料与方法
1.1 试验材料
基因克隆和启动子扩增所用材料为滇龙胆无菌苗幼叶,采自玉溪师范学院分子生物学实验室,经云南省农业科学院药用植物研究所金航研究员鉴定为滇龙胆,采样日期为2014年5月17日。qPCR(实时定量基因扩增)分析使用的二年生滇龙胆植株采自临沧市云县临沧耀阳生物药业科技有限公司后箐基地,由该公司技术经理董顺福鉴定为滇龙胆,采样日期为2018年4月30日。
1.2 试验方法
1.2.1 GrDL7H基因的扩增 采用多糖植物组织提取试剂盒提取滇龙胆幼叶总RNA,采用逆转录试剂盒合成cDNA,采用植物基因组DNA提取试剂盒[宝生物工程(大连)有限公司]提取滇龙胆叶片DNA。根据前期测序的滇龙胆转录组GrDL7H基因转录本序列,设计1对特异引物GrDL7H EcoRⅠ-F、GrDL7H-R(表1)。以cDNA为模板进行PCR扩增,反应体系如下:PrimeSTAR Max Premix(2×,TaKaRa)25 μL,cDNA模板2 μL,正反向引物(10 μmol/L)各1 μL,加ddH2O补足50 μL。PCR反应条件如下:98 ℃ 10 s,55 ℃ 10 s,72 ℃ 10 s,30个循环;72 ℃ 5 min。PCR产物经1.0%琼脂糖凝胶电泳分离,然后割胶,使用胶回收试剂盒(德国QIAGEN)对目的片段进行回收,将其连接到pMD19-T载体上。转化大肠杆菌DH5α[宝生物工程(大连)有限公司]后进行蓝白斑筛选,挑取12个白斑摇菌;使用试剂盒(北京百泰克生物技术有限公司)提取质粒,经酶切检测正确后进行测序[生工生物工程(上海)股份有限公司],获得重组质粒pMD19-fGrDL7H。根据测序结果,设计基因特异性引物GrDL7H 3RACE-F,进行3′ cDNA末端快速扩增(RACE)。使用DNAMAN软件比对后,进行基因片段拼接,获得GrDL7H基因ORF(开放阅读框)全长。设计基因特异性引物,分别以cDNA和gDNA为模板进行PCR扩增,PCR产物进行TA克隆后,分别进行DNA测序。
1.2.2 GrDL7H基因启动子的克隆 根据“1.2.1”节中克隆到的滇龙胆GrDL7H基因序列,设计基因特异引物GrDL7H-sp2和GrDL7H-sp3(表1)。采用Universal Genome Walker 2.0试剂盒,分别使用DraⅠ、EcoR V、PuvⅡ和StuⅠ酶切滇龙胆基因组DNA,加接头后,构建文库DL1、DL2、DL3和DL4,然后进行2轮PCR扩增。第1轮PCR总体系为25 μL,具体成分如下:DL1 g DNA 1 μL,50×Advantage 2聚合酶混合物0.5 μL,10×Advantage 2 PCR Buffer 1 μL,AP1(10 μmol/L)1 μL,dNTP(10 mmol/L)1 μL,GrDL7H-sp2(10 μmol/L)1 μL,ddH2O 19.5 μL,反应条件为94 ℃ 25 s、72 ℃ 3 min,7个循环;94 ℃ 25 s、67 ℃ 3 min,32个循环;63 ℃ 7 min。第2轮PCR总体系为25 μL,具体成分如下:第1轮PCR产物 1 μL,50×Advantage 2聚合酶混合物0.5 μL,10×Advantage 2 PCR 1 μL,AP1(10 μmol/L) 1 μL缓冲液,dNTP(10 mmol/L)1 μL,GrDL7H-sp3(10 μmol/L)1 μL,ddH2O 19.5 μL,反应条件为94 ℃ 25 s、72 ℃ 3 min,5个循环;94 ℃ 25 s、67 ℃ 3 min,20个循环;63 ℃ 7 min。第2轮PCR产物经TA克隆,获得重组质粒pMD19-pGrDL7H,菌液送生工生物工程(上海)股份有限公司进行DNA测序。
1.2.3 GrDL7H基因及启动子的生物信息学分析 用美国国立生物技术信息中心(NCBI)网站的BLAST程序进行序列比对,用Genetyx 6.1.8软件进行翻译,用DNAMAN 7进行多序列比对;用Clustal X 2.1进行比对,然后使用MEGA 6.0软件内置的NJ(邻接)法构建系统进化树,设置Bootstrap(自抽样)=1 000;利用在线数据库(http://molbiol. edu.ru/eng/scripts/01_11.html)进行稀有密码子分析。使用ChloroP服务器v 1.1进行叶绿体转运肽预测;使用Interpro软件进行保守结构域预测;使用ProtScale软件进行疏水性分析,方法为Hphob./Kyte & Doolittle量表法;使用PredictProtein对二级结构进行预测;使用Phyre2(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index)对三级结构进行预测;利用Expasy中的TMHMM工具预测蛋白质的跨膜螺旋区;利用在线工具Wolf Psort预测蛋白質的亚细胞定位情况。使用GT-AG原则,将ORF序列与gDNA序列进行比对,进行外显子与内含子的分析。使用启动子在线分析网站PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对克隆到的GrDL7H基因启动子序列进行分析。
1.2.4 GrDL7H基因的荧光实时定量分析 分别取二年生滇龙胆的根、茎、叶、花,使用MiniBEST Plant RNA Extraction Kit[宝生物工程(大连)有限公司]提取总RNA,使用PrimeScript RT reagent Kit with gDNA Eraser[宝生物工程(大连)有限公司]处理除去基因组DNA,然后反转录合成第1链cDNA。以转录组中GrACTIN基因(GenBank登录号为KM061807)作为内参和GrDL7H基因ORF序列设计特异性引物(表1)。PCR体系如下:TB GreenPremix Ex TaqⅡ(Tli RNaseH Plus)10 μL,ddH2O 7 μL,cDNA 1 μL,正反向引物各1 μL。qPCR条件:95 ℃ 5 min;95 ℃ 15 s,60 ℃ 15 s,72 ℃ 20 s,重复4次。在LightCycler 480 Ⅱ荧光定量PCR仪(Roche,瑞士)上进行扩增反应,扩增曲线、溶解曲线、标准曲线由软件自动生成。使用内参基因GrACTIN表达校准后,计算根、茎、叶中GrDL7H基因的相对表达量。采用比较CT值的2-ΔΔCT的方法进行定量数据的分析处理。
2 结果与分析
2.1 滇龙胆GrDL7H基因的克隆
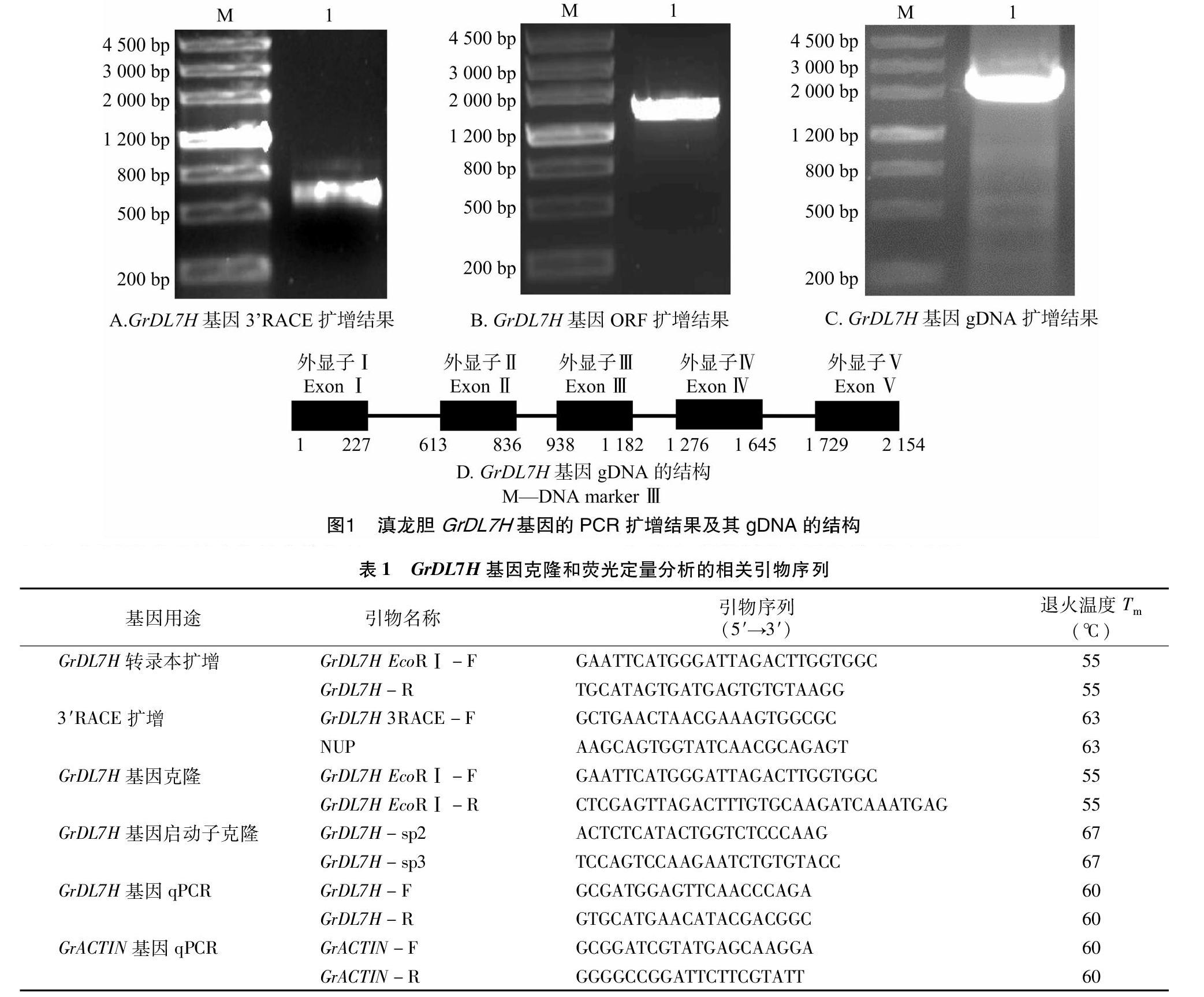
以滇龙胆幼叶cDNA为模板,使用表1中的基因特异性引物GrDL7H EcoRⅠ-F和GrDL7H-R扩增出约1 500 bp的片段。通过TA克隆获得重组质粒pMD19-fGrDL7H,然后进行DNA测序。结果表明,fGrDL7H长1 502 bp,不含3′末端。使用SMART RACE试剂盒进行扩增,获得600 bp DNA片段(图1-A)。使用DNAMAN软件对GrDL7H基因片段进行组装,结果发现GrDL7H基因ORF全长1 542 bp(图1-B,登录号:KP340980)。使用GrDL7H EcoR Ⅰ-F和GrDL7H EcoR Ⅰ-R对GrDL7H基因的基因组DNA(gDNA)进行扩增,结果显示GrDL7H基因长2 154 bp(图1-C,登录号:KT306971),包含5个外显子和4个内含子(图1-D)。
2.2 GrDL7H基因的生物信息学分析
2.2.1 GrDL7H基因的ORF分析和多序列比对分析 利用Genetyx 6软件对GrDL7H基因的ORF序列进行分析,该基因(GenBank登录号为KP722033.1)长1 542 bp,编码513个氨基酸。
利用GenBank数据库的BLASTP程序对GrDL7H蛋白质进行比对分析,结果表明,滇龙胆GrDL7H与萝芙木RsDL7HT(78.17%)、黄金鸡纳树CcDL7H(77.78%)、长春花CrDL7H(77.73%)的序列相似性均较高,与拟南芥AtDL7H(51.47%)的蛋白质相似性稍低。利用DNAMAN 7将GrDL7H蛋白质序列与NCBI中相似性较高的部分序列进行多序列比对分析,结果表明,GrDL7H蛋白质与已知蛋白质序列的保守性较高(图2)。利用MEGA 7.0软件将GrDL7H氨基酸序列与NCBI中相似性较高的部分序列进行系统发育分析,结果显示,滇龙胆GrDL7H蛋白质与黄金鸡纳树CcDL7H的亲缘关系较近(图3)。
2.2.2 GrDL7H蛋白质的理化特性分析 使用ExPASy ProtParam tool对GrDL7H蛋白质进行分析,结果表明,GrDL7H蛋白质单体的相对分子质量为59 077 u,理论等电点(pI值)为8.88,这与长春花CrDL7H蛋白质类似[8];带正电氨基酸残基(Arg+Lys)为70个,带负电氨基酸残基(Asp+Glu)为63个,化学方程式为C2 686H4 240N708O743S23;不稳定指数为31.37,属稳定蛋白质;脂肪指数为94.81;总平均疏水性(GRAVY)为-0.173,为亲水蛋白质(图4)。GrDL7H蛋白质含有20种基本氨基酸,其中亮氨酸含量最高,为11.9%;其次是赖氨酸、异亮氨酸、苏氨酸,含量分别为7.60%、6.40%、6.40%;半胱氨酸含量最低,为1.00%。
2.2.3 GrDL7H蛋白质的二级结构、三级结构分析 利用PredictProtein软件对GrDL7H蛋白质进行二级结构分析,结果表明:该蛋白质二级结构中α-螺旋(H)占54.58%,环(L)占37.43%,延伸带(E)占7.99%。利用Phyre 2软件预测GrDL7H蛋白质的三级结构,结果见图5,该模型以细胞色素p4504b1酶[c5t6qA]为模板,在第15~513位氨基酸处建模,序列相似度为24%。
2.2.4 GrDL7H蛋白质序列的分析归类 使用InterPro 6.0在线工具对GrDL7H蛋白质进行分析和归类,结果表明,GrDL7H蛋白质属于细胞色素P450超家族E类、类群Ⅰ(IPR002401)的成员,包含细胞色素P450保守结构域(IPR036396)(图6)。GO预测结果表明,在生物过程方面GrDL7H蛋白质参与氧化还原过程,在分子功能方面,该蛋白质参与铁离子结合、氧化还原酶活性、血红素结合,在细胞成分方面未预测到结果。
2.2.5 GrDL7H蛋白质信号肽分析 利用ExPASy SignalP 41服务器分析GrDL7H蛋白质,未发现信号肽,表明该蛋白质为非分泌型蛋白质。利用TMHMM 2.0工具预测GrDL7H蛋白质的跨膜螺旋区,结果表明:GrDL7H蛋白质在5~27位氨基酸含有跨膜区域,其余的1~4位氨基酸在细胞内,28~513位氨基酸在细胞外(图7)。
2.2.6 GrDL7H蛋白质亚细胞定位分析 使用WoLF PSORT软件进行GrDL7H蛋白的亚细胞定位分析,结果显示,GrDL7H蛋白质在叶绿体、细胞核、核骨架、细胞膜、细胞质、液泡的定位系数分别为 6.0、3.5、2.5、2和1.0。
2.2.7 GrDL7H基因稀有密码子分析 使用在线软件对GrDL7H基因进行稀有密码子分析,结果表明,在大肠杆菌和酿酒酵母中,GrDL7H基因中稀有密码子分别占1.36%、0.19%,且均无二联或三联稀有密码子连续出现的情况,因此可选用大肠杆菌表达菌BL21、Rosetta(DE3)或酿酒酵母进行蛋白质表达。
2.3 GrDL7H基因启动子的克隆
以滇龙胆叶片DNA为模板,使用基因特异性引物分别进行2轮PCR扩增, 第2轮PCR扩增产物通过TA克隆获得重组质粒pMD19-pGrDL7H,测序后,上传至NCBI数据库,获得登录号(GenBank登录号:MG832585)。对获得的GrDL7H启动子进行生物信息学分析,结果表明,该启动子不但在-30区域附近含有转录起始的核心启动子元件TATA盒,而且含有启动子和增强子区域通用顺式作用元件CAAT盒;此外,还具有6个光应答元件、 1个赤霉素应答元件、6个参与茉莉酸甲酯应答的顺式调控元件、1个热胁迫应答元件和1个参与生物钟控制的顺式调控元件(表2)。
2.4 GrDL7H基因的组织表达分析
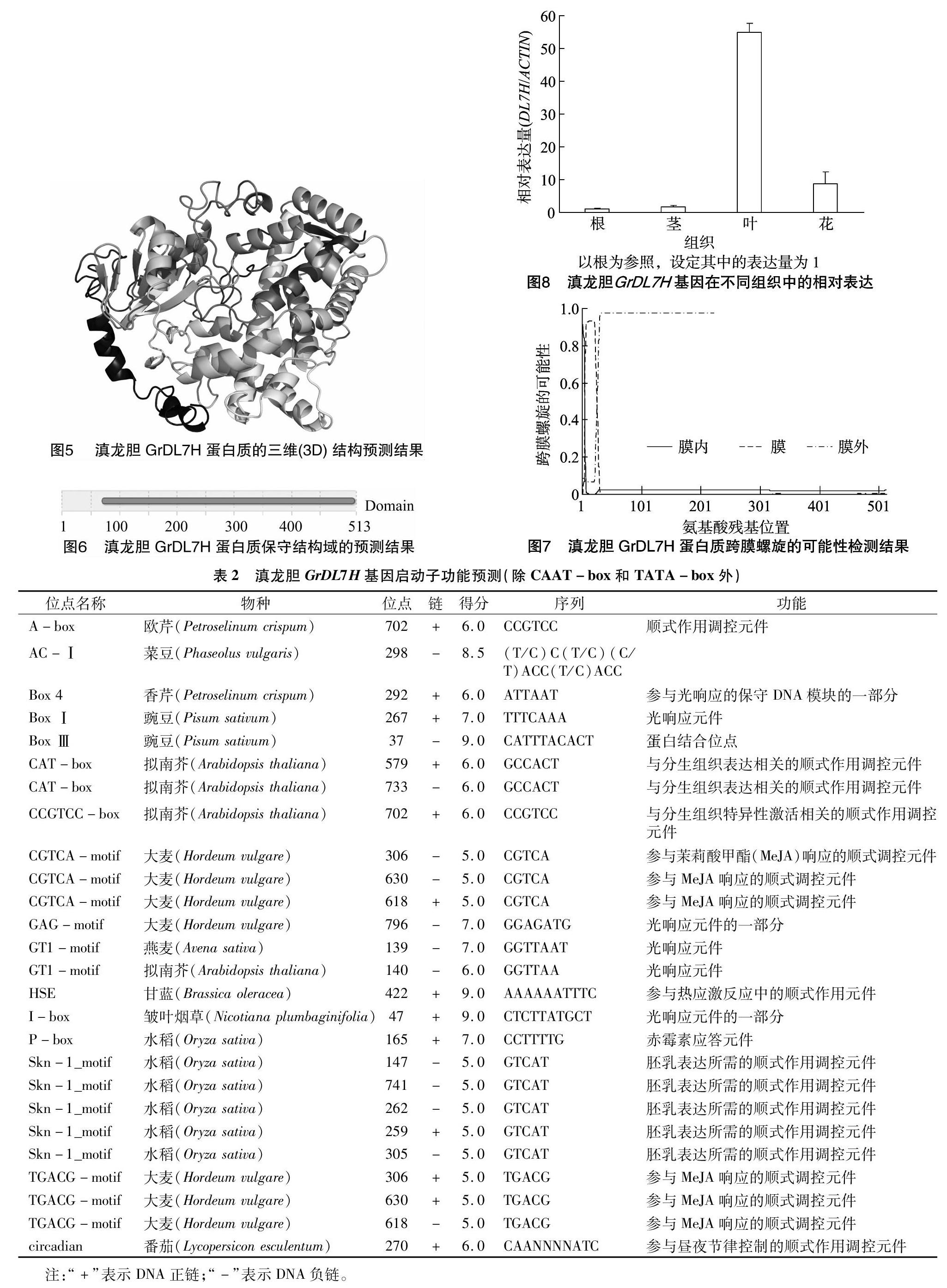
取二年生滇龙胆的根、茎、花和叶,通过荧光定量PCR(qRT-PCR)检测GrDL7H基因在根茎花叶等不同组织中的表达情况。结果显示,GrDL7H基因表达量最高的组织是叶,其表达量大约分别是根、茎、花中的50、32、6倍;其次是花,表达量最低的是根(图8)。这与川西獐牙菜SmDL7H基因在根茎叶花中的表达模式类似[7]。在长春花中,CrDL7H基因主要在叶中表达,随着叶龄的增加,其表达量逐渐降低,相应的长春碱与裂环番木鳖苷的含量逐渐减少;其次是茎和花,表达量最低的是根[8]。这些结果表明,叶是滇龙胆龙胆苦苷生物合成的主要场所。
3 讨论
滇龙胆的主要药效成分为龙胆苦苷,属于环烯醚萜类物质。该类物质的合成主要包括3个阶段,第1阶段是通过甲羟戊酸(MVA)和甲基赤藓糖-4-磷酸(MEP)途径合成IPP(异戊烯焦磷酸)和DMAPP(二甲基丙烯基二磷酸-三铵盐)的基本前体,第2阶段是IPP和DMAPP在异戊烯基转移酶和萜类合酶作用下形成萜类化合物和中间产物,第3阶段是萜類化合物的碳骨架结构在后修饰酶的催化下,分别进行羟基化、甲基化、糖基化等,形成结构稳定、具有活性的萜类化合物[4]。为阐明龙胆苦苷生物合成途径及其调控机制,本研究克隆了1个萜类合成第2阶段的GrDL7H基因,其表达情况可能影响龙胆苦苷的生物合成。由于许多萜类合成途径中的底物不能通过商业化途径获得,导致学界对DL7H基因的研究较少,目前研究比较清楚的仅有长春花中的CrDL7H基因。本研究从滇龙胆幼叶中克隆获得GrDL7H基因,通过多序列比对分析,表明滇龙胆GrDL7H与其他植物DL7H蛋白质具有较高的相似性,特别是与长春花CrDL7H蛋白质的序列相似性高达77.73%,表明二者具有相似的结构与功能。在系统发育分析中,滇龙胆GrDL7H与黄金鸡纳树CcDL7H的亲缘关系较近,且GrDL7H蛋白质具有典型的CYP450蛋白质结构域,属于E类家族成员,这些结果表明,所克隆的基因为DL7H基因。
在长春花中,CrDL7H基因在根、茎、叶、花芽和花中均有表达,但在第1对幼叶中的表达量远远高于其他组织[8]。组织表达特异性研究结果表明,滇龙胆GrDL7H基因在叶中的表达量最高,远远高于根、茎和花,因此推测滇龙胆叶片为龙胆苦苷生物合成的主要器官,这与朱宏涛等的研究结果[10]一致。目前研究表明,滇龙胆全株可以入药[11-12],但其根中龙胆苦苷含量最高[13]。因此,根中的龙胆苦苷主要是由叶片和茎中合成的龙胆苦苷通过运输到达根中进行积累而成的。
在真核生物基因中,启动子由核心启动子和上游启动子2个部分组成,是位于转录起始位点及其5′端上游近端大约100~200 bp的1组具有独立功能的DNA序列,每个元件长度约为7~20 bp,是决定RNA聚合酶转录起始和转录频率的关键元件[14]。本研究扩增到的滇龙胆GrDL7H基因启动子片段大小为818 bp,不但具有核心启动子TATA盒上游启动子CAAT盒,而且具有6个光应答的元件、1个参与赤霉素应答元件、6个参与茉莉酸甲酯应答的顺式调控元件、1个热胁迫应答元件、1个参与生物钟控制的顺式调控元件,表明GrDL7H基因的表达受光、赤霉素、茉莉酸甲酯、热胁迫、生物钟等多种因素的调控,这同时也表明,可通过调控基因表达来提高龙胆苦苷的含量。
本研究為滇龙胆GrDL7H基因功能的解析奠定了基础。今后将对GrDL7H基因进行蛋白质表达、纯化和酶活分析、基因调控机制分析来研究其功能,从而为龙胆苦苷生物合成途径及其调控机制的阐明奠定基础。
参考文献:
[1]Suyama Y,Tanaka N,Kawazoe K A,et al. Rigenolides B and C,conjugates of norsecoiridoid and secoiridoid glucoside from Gentiana rigescens Franch[J]. Tetrahedron Letters,2017,58(15):1459-1461.
[2]Suyama Y,Kurimoto S I,Kawazoe K,et al. Rigenolide A,a new secoiridoid glucoside with a cyclobutane skeleton,and three new acylated secoiridoid glucosides from Gentiana rigescens Franch[J]. Fitoterapia,2013(91):166-172.
[3]金 航,张 霁,张金渝,等. 滇龙胆[M]. 昆明:云南科技出版社,2013.
[4]黄璐琦,刘昌孝. 分子生药学[M]. 3版.北京:科学出版社,2015.
[5]Salim V,Yu F,Altarejos J,et al. Virus-induced gene silencing identifies Catharanthus roseus 7-deoxyloganic acid-7-hydroxylase,a step in iridoid and monoterpene indole alkaloid biosynthesis[J]. Plant Journal,2013,76(5):754-765.
[6]王彩云,张晓东,沈 涛,等. 龙胆苦苷生物合成途径研究进展[J]. 江苏农业科学,2014,42(3):4-10.
[7]Liu Y E,Wang Y,Guo F X,et al. Deep sequencing and transcriptome analyses to identify genes involved in secoiridoid biosynthesis in the Tibetan medicinal plant Swertia mussotii[J]. Scientific Reports,2017(7):43108.
[8]Salim V,Yu F,Altarejos J,et al. Virus-induced gene silencing identifies Catharanthus roseus 7-deoxyloganic acid-7-hydroxylase,a step in iridoid and monoterpene indole alkaloid biosynthesis[J]. Plant Journal,2013,76(5):754-765.
[9]Zhang X,Allan A C,Li C,et al. De Novo assembly and characterization of the transcriptome of the Chinese medicinal herb,Gentiana rigescens[J]. International Journal of Molecular Sciences,2015,16(5):11550-11573.
[10]朱宏涛,郑传伟,赵 平,等. 野生坚龙胆及其组培苗中龙胆苦苷的含量分析[J]. 天然产物研究与开发,2011,23(3):482-485.
[11]赵志莲,张 琳,刘卫红,等. 滇龙胆裂环烯醚萜类活性成分积累规律的研究[J]. 时珍国医国药,2016,27(9):2073-2075.
[12]游维丽. 龙胆地上、地下部分药理作用的比较[J]. 内蒙古中医药,2017,36(8):135-136.
[13]杨美权,张金渝,沈 涛,等. 不同栽培模式对滇龙胆中龙胆苦苷含量的影响[J]. 江苏农业科学,2011(1):287-289.
[14]朱玉贤,李 毅,郑晓峰,等. 现代分子生物学[M]. 4版.北京:高等教育出版社,2013.
猜你喜欢
基层中医药(2022年7期)2022-11-17
天天爱科学(2021年2期)2021-09-10
孩子·小学版(2020年7期)2020-02-24
中成药(2017年12期)2018-01-19
种子(2016年9期)2016-12-04
中成药(2016年4期)2016-05-17
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
现代农业科技(2009年19期)2009-03-20
中学生数理化·八年级数学华师大版(2008年3期)2008-08-26
中国新闻周刊(2004年11期)2004-04-07
