
武夷山中亚热带常绿阔叶林凋落物量动态研究
2019-11-04 07:28:20林捷叶功富黄石德高伟谭芳林金昌善徐自坤游慧明欧阳亚群
防护林科技 2019年10期
林捷,叶功富,黄石德,高伟,谭芳林,金昌善,徐自坤,游慧明,欧阳亚群
(1.福建省林业科学研究院,福建 福州 350012;2.武夷山自然保护区管理局,福建 武夷山 353000)
森林凋落物量是指单位面积、单位时间内的森林地段上所有的森林凋落物总量。是森林生态系统物质和能量循环流动的主要途径,凋落物通过微生物的分解向土壤输入大量的养分和能量,对林地土壤肥力的维持极为重要,是森林自我培肥地力主要来源之一[1]。凋落物的数量及组成可以反映出林木在各个生长发育过程中新陈代谢的情况,其影响因素较多,主要有群落里树种组成结构及生境等气候条件。我国对凋落物量的研究从20世纪80年代开始有较大进展,至今已涉及不同气候带的不同森林群落,包括山地雨林、半落叶季雨林、鼎湖山南亚热带常绿阔叶林、广西亚热带常绿阔叶林、浙江天童常绿阔叶林等,但已有报道多为1年或几年的研究结果,而凋落物量的变化往往与当地气象要素有一定的相关关系,需要长期的定位监测才能得到更有意义的结论。
米槠(Castanopsiscarlesii)、甜槠(C.eyrei)为常绿乔木,具分布范围大,适应性强、稳定性高等特点,主要生长在海拔300~1 700 m丘陵、山地的疏林或密林中,在针叶阔叶混交林或常绿阔叶林中常为主要树种,有时会形成小片纯林,作为中亚热带顶级群落的米槠、甜槠天然林物种组成丰富,其森林服务功能价值较高,这对改善人工营造用材林的景观结构、提高人工林培育生境多样性和生态系统稳定性具有重要意义[2,13]。已有研究人员对不同地点的常绿阔叶林的群落结构和种群动态等方面进行了研究[3~11],但目前对中亚热带米槠、甜槠天然林群落凋落物动态变化的研究很少。
本文以福建武夷山森林生态系统国家定位观测研究站内米槠、甜槠天然林群落为研究对象,观察研究米槠、甜槠天然林群落凋落物量的动态变化特征,以便为该群落的长期监测提供基础资料,对深入了解中亚热带森林生态系统的结构和功能,以及持续利用中亚热带常绿阔叶林资源具有重要意义。
1 试验地概况
福建省武夷山自然保护区(117°27′—17°51′ E,27°35′—27°54′ N),位于福建南平的武夷山、建阳、光泽和邵武4县(市)交界处,为现今世界同纬度留存面积最大且保护最完整的中亚热带森林生态系统[9],多年平均气温为8.5~18.0 ℃,极端最低温为-15 ℃,年降水量为1 486~2 150 mm左右,年相对湿度为82%~85%,主要土壤类型为山地红壤、黄红壤,主要森林群落是以米槠、青冈(Cyclobalanopsisglauca)、甜槠、丝栗栲(Castanopsisfargesii)等群落为主的中亚热带常绿阔叶林。
2 研究方法
2.1 样地设置
样地位于武夷山森林生态系统国家定位观测研究站的米槠、甜槠天然林内,海拔分别为600 m和650 m,坡度分别为30°和40°,坡向分别为南坡和西南坡,选择立地条件相似、人为影响较小、生长发育良好、林分结构完整的24个10 m×10 m标准地作为国定样地。
米槠天然林群落中乔木层主要有米槠、甜槠、黄瑞木(Adinandramillettii)等,灌木主要有少叶黄杞(Engelhardtiafenzelii)、弯蒴杜鹃(Rhododendronhenryi)、密花树(Rapaneaneriifolia),藤本主要有尖叶菝葜(Smilaxarisanensis)、酸味子(Antidesmajaponicum)、马银花(Rhododendronovatum),草本层主要以芒萁(Dicranopterisdichotoma)为主。
甜槠天然林群落中乔木层主要有甜槠、少叶黄杞、米槠等,灌木主要有鹿角杜鹃(Rhododendronlatouchae)、绿樟(Meliosmasquamulata)、赤楠(Syzygiumbuxifolium)等,藤本主要有流苏藤(Thysanospermumdiffusum)、网脉叶酸藤果(Embeliarudis)、念珠藤(Alyxiafascicularis)等,草本层主要以里白(Diplopterygiumglaucum)为主。
两个样地的表层土壤理化性质见表1。

表1 样地表层土壤的理化特征(2003年)
2.2 凋落物收集与测定
在样地内设置10个1 m×1 m的凋落物收集框,按照局部控制加随机处理的原则分布(还要兼顾坡向、坡位和密度),用尼龙网为收集网,其孔径为0.5 mm,收集框距地约25 cm。试验从2003年1月开始,每月的上旬和下旬各收集一次,到2013年12月为止,收集现场称鲜质量,并将凋落物按叶、枝、花果、树皮和碎屑分为五个组成部分,分别记录数据,将其带回定位站的实验室内,先用80 ℃杀青,再用65 ℃烘干至恒质量,称出干质量,算出含水率,最后就可以换算出单位面积内的凋落物干质量。
3 结果与分析
3.1 凋落物总量动态
米槠、甜槠天然林凋落物11年来的总干质量变化如图所示。
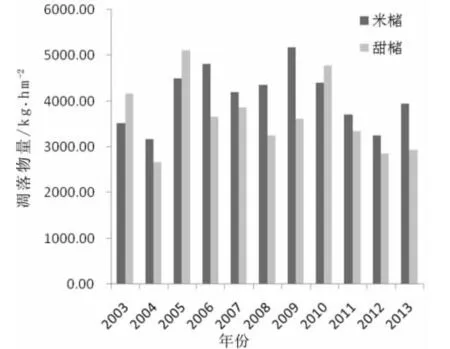
图1凋落物量年动态变化(2003—2013)
从图1中可知,米槠林群落凋落物总量最高值为2009年的5 177.95 kghm-2,最低值是2004年的3 174.74 kghm-2;甜槠林群落总凋落量最高值为2005年的5 110.71 kghm-2,最低值也是2004年的2 670.40 kghm-2。从2003年到2013年11年监测期内米槠林群落凋落物量仅2003、2005、2010这3个年份小于甜槠林群落,米槠林群落其余8个年份都大于甜槠林群落,这表明米槠林群落年凋落物量是大于甜槠林的,它们的年平均凋落物量分别是4 096.32 kghm-2、3 662.57 kghm-2,定位站长期观测研究的中亚热带常绿阔叶林的年凋落物量总平均是3 879.45 kghm-2。从两个群落的年凋落物量动态变化图来看,凋落物量升高、下降趋势不明显,说明这两个群落都还是处在生长活跃期内,所以年季凋落物量变化显著。
3.2 各组分凋落物量

表2 凋落物各组分年动态变化及其占总量百分比
中亚热带常绿阔叶林的凋落物可分为枝、叶、花果、树皮、碎屑五个组成部分。米槠林群落和甜槠林群落的各组分年动态变化及其占总量的百分比见表2。从表2可以看出米槠林枝占总量的百分比分布在7.60%~19.01%之间,对应的时间分别是2006年和2010年,平均占总量的百分比是14.41%,其标准差是3.87,年际变异系数是26.87;叶占总量的百分比分布在44.45%~79.87%之间,对应的时间分别是2004年和2007年,平均占总量的百分比是70.32%,其标准差是9.94,年际变异系数是14.14;花果占总量的百分比分布在2.02%~27.10%之间,对应的时间分别是2003年和2004年,平均占总量的百分比是8.26%,其标准差是8.59,年际变异系数是103.90,主要是2004年其花果的凋落量比其他年份大了许多,为正常年份的2~3倍,另外2009年的花果的凋落量也是如此,所以显得变异系数比较大;树皮占总量的百分比分布在0.20%~3.10%之间,对应的时间分别是2004年和2007年,平均占总量的百分比是1.18%,其标准差是0.87,年际变异系数是73.91;碎屑占总量的百分比分布在3.78%~9.00%之间,对应的时间分别是2003年和2004年,平均占总量的百分比是5.83%,其标准差是1.35,年际变异系数是23.15。
甜槠林枝占总量的百分比分布在5.68%~20.99%之间,对应的时间分别是2007年和2005年,平均占总量的百分比是12.52%,其标准差是4.02,年际变异系数是32.11;叶占总量的百分比分布在59.48%~82.11%之间,对应的时间分别是2004年和2007年,平均占总量的百分比是73.99%,其标准差是6.76,年际变异系数是9.13;花果占总量的百分比分布在2.22%~10.48%之间,对应的时间分别是2011年和2004年,平均占总量的百分比是5.06%,其标准差是2.66,年际变异系数是52.68;树皮占总量的百分比分布在0.43%~5.92%之间,对应的时间分别是2008年和2004年,平均占总量的百分比是2.08%,其标准差是1.50,年际变异系数是72.24;碎屑占总量的百分比分布在3.51%~8.62%之间,对应的时间分别是2003年和2006年,平均占总量的百分比是6.35%,其标准差是1.22,年际变异系数是19.15。
3.3 凋落节律
米槠、甜槠天然林的凋落物总量的月变化节律如图2。从图中可知,米槠、甜槠天然林的凋落物总量最低值分别为162.01、139.23 kghm-2,都在1月;最高值都是在4月,分别为770.81、561.42 kghm-2。两个群落年凋落物曲线具有明显的变化规律,其动态曲线为双峰形,主峰都在4月,次峰也都是当年的11月,其季节变化为夏季>冬季>秋季>春季。

图2年凋落总量凋落节律
3.4 凋落物各组分的凋落节律
图3是米槠、甜槠群落的各组分凋落物曲线图,从凋落物枝的月变化可知,米槠林群落的凋落曲线是单峰型的在3月凋落量达到最大,凋落量为72.12 kghm-2,而甜槠林群落的凋落曲线是双峰形,峰顶分别是3月和9月,凋落量分别为53.08、63.57 kghm-2,米槠林群落凋落枝最少值是6月为33.89 kghm-2,甜槠林群落凋落枝最少值是6月为22.96 kghm-2。
从凋落物叶的月变化可知,米槠林群落的凋落曲线是双峰形的在4月和10月凋落量达到最大,凋落量分别为624.53、298.02 kghm-2,而甜槠林群落的凋落曲线也是双峰形,并与年凋落节律一致,峰顶分布是4月和11月,凋落量分别为456.81、289.01 kghm-2,两个群落凋落叶最少值都是1月,分别为86.36、87.38 kghm-2。

图32个群落各组分月凋落节律
从花果凋落物的月变化来看,米槠林群落的凋落曲线是双峰形的在4月和11月凋落量达到最大与年凋落节律一致,凋落量分别为41.78、112.42 kghm-2,而甜槠林群落的凋落曲线是多峰型,峰顶分布是4月、6月和11月,凋落量分别为22.39、22.69和27.06 kghm-2,两个群落花果凋落最少值都是2月,分别为2.62、3.42kghm-2。
从树皮凋落物的月变化来看,米槠林群落的凋落曲线是多峰型的,在2月、6月和12月凋落量达到峰值,凋落量分别为4.34、5.41和7.62 kghm-2,而甜槠林群落的凋落曲线是双峰形,峰值分布在9月和12月,凋落量分别为11.09、10.92 kghm-2,米槠林群落树皮凋落最少值在3月,为2.32 kghm-2,甜槠林群落凋落枝最少值在1月,为3.22 kghm-2。
从凋落物碎屑的月变化来看,米槠林群落的凋落曲线是多峰型的,在4月、7月和10月凋落量达到峰值,凋落量分别为39.10、24.63和25.61 kghm-2,而甜槠林群落的凋落曲线是双峰形,峰值分布在4月和7月,凋落量分别为31.34、25.24 kghm-2,两个群落凋落碎屑最少值都在2月,分别为10.33、10.95 kghm-2。
4 结论与讨论
中亚热带常绿阔叶林内以米槠、甜槠为主的两个群落,总凋落量平均为4.10、3.66 thm-2,稍低于福建武夷山天然杉木混交林5.034 thm-2凋落量、但远低于广西亚热带常绿阔叶林7.99 thm-2凋落量、广东鼎湖山南亚热带常绿阔叶林8.45 thm-2凋落量、浙江天童常绿阔叶林8.896 thm-2凋落量,只与程伯容等研究的长白山阔叶红松林3.84 thm-2相近。可见森林群落树种组成的差异,所处纬度、温度、降水等生境条件的不同,凋落物的年凋落量是大不相同的。是否也与这些群落里主要树种所处的生长季不同有关,比如鼎湖山南亚热带常绿阔叶林的年凋落量呈下降趋势,而武夷山中亚热带常绿阔叶林的年季凋落量是变异较大,但无明显下降的痕迹。是否还与收集的年限有关等等,因为上述的许多研究的凋落物收集年限都只有一年或2、3年,这有待于定位站今后的持续观察。
凋落物的组成有一定的比例规律,落叶在森林凋落物中一般情况下都占据着绝对的优势,落叶占凋落物总量的比例常常为60%~80%,小枝可以占到10%~15%,其他成分仅占10%左右。米槠林和甜槠林群落的年平均落叶占凋落物总量的比例分别是70.32%、73.99%,小枝占年平均凋落物总量的比例分别是14.41%、12.52%,这个结果与大多数研究人员得出的结论相一致。
中亚热带常绿阔叶林的凋落物在全年的凋落过程是连续的,这可以从试验安排定位站工作人员每半个月收集凋落物一次的收集频率可知,植物由于受到生长特性差异以及光照、气温、降水、风力等因素的影响,森林凋落物的量具有明显的季节变化规律,其凋落物各个组成的数量在各个月的分布是不相同的。许多研究都表明凋落物季节变化的曲线,可以是单峰的,也可以是双峰的或不规则形的,差异较大。某一曲线的出现主要是与组成群落的目的树种有关,本研究的米槠林、甜槠林群落其年凋落物动态曲线为双峰形,主峰都在4月,次峰也都在当年的11月。
图2、3中可以看出米槠林和甜槠天然林的凋落物总量及其组成的季节变化较明显。凋落物量及其组成的最小值都出现在春季,即1—3月,而最大值大都是出现在夏季,也就是4—6月,并以4月为主,冬季的11月也都是米槠林和甜槠林分凋落物及各组分产量的次高峰期,仅次于夏季的4—6月。根据图中的凋落物及各组成的月变化曲线,可以将凋落物及各组成量的年变化按季节划分为4各阶段。第一阶段为春季,即1—3月,凋落物产量最小,仅占全年总凋落量的10%左右,可能因为该阶段是新叶和幼枝才刚开始萌动并生长发育的时期,因而凋落物的数量总体较小。第二阶段为夏季,即4—6月,凋落物量远高于第一阶段,占据全年凋落物量的40%。该阶段林木生产旺盛,是米槠林、甜槠林群落一年中的主要生长期;第三阶段为7—9月,即秋季,凋落物量占据全年的20%左右,这一阶段福建虽然多台风,但在该阶段米槠、甜槠群落生长还未停止。第四阶段为冬季,即10—12月,应该是随着冬季的来临,生境温度在不断降低,林木生长逐渐停止,老的枝叶大量脱落,凋落量也就大量增加。该阶段凋落物量占全年的30%左右,因而米槠、甜槠天然林群落各组分凋落物产量的季节表现为夏季>冬季>秋季>春季。
猜你喜欢
长江技术经济(2024年5期)2024-01-01 00:00:00
地下水(2023年5期)2023-10-13 06:03:28
绿色中国(2019年18期)2020-01-04 01:57:08
亚热带农业研究(2019年2期)2019-09-05 09:39:06
农家科技下旬刊(2017年8期)2017-11-13 18:09:22
植物研究(2017年5期)2017-11-10 01:47:20
星星·诗歌原创(2017年5期)2017-08-13 10:09:55
五邑大学学报(社会科学版)(2016年2期)2016-03-13 09:24:47
中国果菜(2015年2期)2015-03-11 20:01:08
亚热带资源与环境学报(2015年4期)2015-01-22 07:06:16
