
雷州半岛不同林龄尾巨桉人工林根系特征及乔木层生物量分配
2019-10-31 11:06:16张学德伍勇黄宇超黄胜张明思苏寒颜炳辉王志超
桉树科技 2019年3期
张学德,伍勇,黄宇超,黄胜,张明思,苏寒,颜炳辉,王志超
雷州半岛不同林龄尾巨桉人工林根系特征及乔木层生物量分配
张学德1,伍勇1,黄宇超1,黄胜1,张明思1,苏寒1,颜炳辉1,王志超2,3*
(1.广西壮族自治区国有七坡林场,广西 南宁 530225; 2. 国家林业和草原局桉树研究开发中心,广东 湛江 524022; 3. 广东湛江桉树林生态系统国家定位观测研究站,广东 湛江 524022)
针对桉树人工林生物量估算难度大、精度低的问题,本文通过研究不同林龄尾巨桉人工林的根系生物量特征、地上部分各器官以及整株生物量分配,建立以胸径为变量的各林龄尾巨桉人工林不同器官的数学模型,为桉树人工林乔木层碳储量的准确测算提供科学依据和数据支持。结果表明:各林龄尾巨桉的根系,随径级的增加,细根和中根差异不大,而大根显著增大,且随着林龄的增大,大根比例迅速增加,细根和中根比例逐渐降低。各林龄尾巨桉不同器官的生物量分配均是树干>树根>树枝>树皮>叶片,2 a生、6 a生及10 a生尾巨桉林分总生物量分别为47.18 t·hm-2、85.09 t·hm-2和254.36 t·hm-2,不同林龄各器官的生物量与胸径的拟合方程2在0.450 ~ 0.989之间,且均达到极显著水平,说明各林龄不同器官的模型方程均适用于尾巨桉乔木层生物量的估测。
尾巨桉;人工林;根系特征;生物量;回归模型
森林作为全球第二大碳库[1],其在维持全球碳平衡、减缓全球气候变化方面影响重大[2]。生物量作为森林生态系统的数量特征,是评估森林固碳能力和固碳效益的重要内容,也是衡量森林生态系统生产力的主要指标[3-5]。人工林作为森林生态系统重要的组成部分,在全球碳循环中发挥着越来越重要的作用。准确获取林分生物量并分析其分配特征,进而提高森林生物量估算精度,是研究森林碳循环中的关键。然而由于地下根系的难操作性,以往很多有关林木生物量的研究往往只关注地上部分,而忽视了根系。随着人们对森林生物量认识的不断加深,根系在全球碳循环等方面的地位和作用也越来越被重视。研究表明,树木根系生物量是重要碳库,平均约占森林生物量的30%[6-7],根系的生物量分配对树木的固着、水分和矿物质的吸收以及改良土壤具有重要作用[8],因此,研究林木根系生物量分配对于认识森林的碳汇功能以及生物化学循环等具有重要的意义。
桉树()作为世界三大速生树种之一,至2015年我国目前种植面积约450万公顷,虽仅占我国人工林面积的6.5%,但年产木材超3 000万立方米,占全国木材产量的26.9%[9]。桉树是我国南方重要的战略树种,其超高的固碳速率使其碳汇功能越来越受到业内人士的重视,但由于缺乏正确的经营,使得桉树人工林目前出现地力衰退、生产力下降的现象,维持桉树人工林长期稳定生产力已成为国内外研究者亟待解决的重点问题。桉树林生物量作为分析桉树人工林生态系统碳循环和碳动态的基础,目前国内已有部分研究,但大多数没有将根系的特征进行细化,只是笼统的一个总生物量,这不利于以较高精度测算桉树人工林的生物量,准确评估其生态系统服务功能价值。因此本研究以2 a生、6 a生、10 a生尾巨桉(×)人工林尾研究对象,调查包括根系在内的整株生物量,分析不同林龄尾巨桉的根系特征及其生物量分配,为桉树人工林生物量的测算、碳储量及碳汇功能的评价提供更高精度的数据支持和理论支撑。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于广东雷州半岛地区,试验样地设置在广东湛江桉树森林生态系统国家定位观测站内(21°30′N,111°38′E),海拔90 m,地势平坦,属海洋性季风气候,年均降雨量约1 500 mm,多集中在5-10月份,为雨季,占全年降雨量的77% ~ 85%;年均温度23℃,年总辐射量4 240 MJ·m-2,年均相对湿度80.4%。试验区土壤类型主要是玄武岩砖红壤,土层厚度2 m以上,0 ~ 80 cm土层内平均有机质含量1.6%以上,pH 4.5 ~ 5.3,土壤肥力属中等水平。乔木层主要有尾叶桉();林下植被丰富,主要灌木有五色梅()、野牡丹()、马樱丹()等;主要草本植物有飞扬草()、飞机草()、蟛蜞菊()以及白花鬼针草()等。
1.2 研究方法
1.2.1 样地的选择
在2 a生、6 a生及10 a生尾巨桉人工林随机分别选取3个20 m × 20 m样地,共计9个样地,并对样地进行每木检尺,各样地的基本信息如表1。
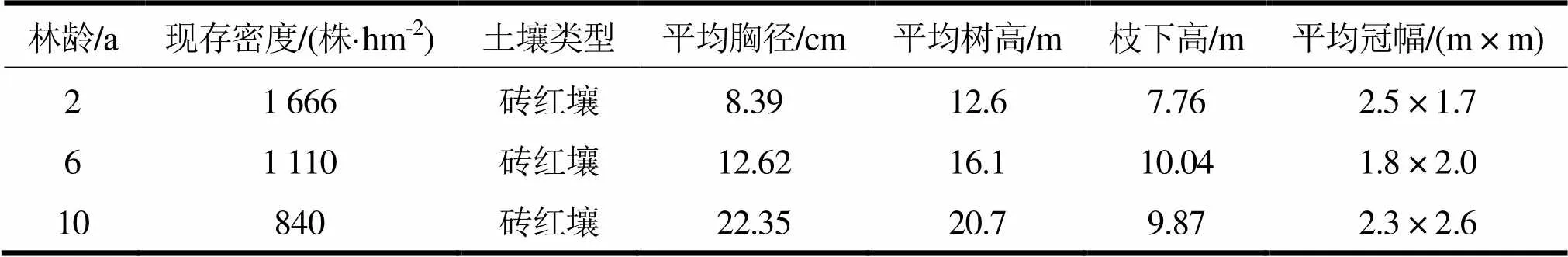
表1 不同林龄尾巨桉人工林样地基本特征
1.2.2 乔木层地上生物量的获取
根据各样地每木检尺的结果,在每块样地中分别按3 cm一个径阶选择3株样木,每个林龄共计9株样树(表2),共计27株样木进行树干解析。分别称取各样木叶片、树枝、树干、树皮的总鲜质量,然后对各部分器官分别取样,并带回实验室,将样品在105℃烘箱内杀青2 h,调至75℃下烘干至恒质量,求出样品各器官干鲜质量比,进而利用各器官质量比换算出样木整株的各器官的生物量及地上总生物量。

表2 各林龄所选样木的基本特征
1.2.3 样木根系特征调查及生物量获取
对选取的样木根系进行全部挖取,并清理干净,利用游标卡尺按照小根(< 2 mm)、中根(2 ~ 5 mm)、大根(>5mm) 3组进行分别称取鲜质量,并分别取样100 g左右,带回实验室进行生物量的测定(方法同1.2.2),最终获得各样木不同根系的分配特征及其样木根系生物量。
1.2.4 各器官生物量与胸径的方程拟合
利用样木各器官(树干、树枝、叶片、树根、树皮)的干质量和总干质量建立各器官及总生物量与样木胸径()的幂回归方程(=aD)。根据样地每木调查的结果及相应的回归方程计算乔木层各个体的生物量,并由此获得乔木层的总生物量。
1.3 数据处理
采用Excel 2016制表并作图;利用SPSS 19.0软件对数据进行多重比较分析、回归方程模型的建立。
2 结果与分析
2.1 不同林龄尾巨桉根系特征
由表3可知,2 a生尾巨桉随着径级的增加,细根和中根均无差异,而大根随着径级的增加而显著增加,其中第Ⅲ径级的大根生物量显著大于第Ⅰ径级(<0.05)。6 a生尾巨桉随着径级的增加,细根无显著性差异,中根第Ⅲ径级显著大于第Ⅰ和Ⅱ径级,而大根随着径级的增加,根生物量均显著增加(<0.05)。10 a生尾巨桉随着径级的增加,细根和中根均无显著差异,而大根第Ⅲ和Ⅱ径级均显著大于第Ⅰ径级,第Ⅲ径级与第Ⅱ径级间无显著差异。随着林龄的增加,根系生物量显著增大,但细根和中根的比例逐渐减低,大根的比例逐渐增大,其中10 a生大根的比例达94.8%。

表3 各林龄不同径级尾巨桉根系特征
注:同列数据后不同小写字母表示<0.05
2.2 不同林龄尾巨桉乔木层生物量估算模型的建立
有关森林生物量的研究证明[10-12],乔木层各器官的生物量与胸径及树高间存在一定的关系,因此可以通过数学模型将乔木生物量与胸径及树高的关系表达出来,进而提高森林生物量的估算精度。经研究发现:各器官及整株乔木生物量与胸径和树高间存在=ax的模型[13],其中为各器官生物量或总生物量,为自变量,可以选用胸径或者胸径平方乘以树高(2),和均是估算参数。由于树高的测定精确度较小,因此本位采用胸径作为自变量,对各林龄尾巨桉各器官及单株生物量进行非线性拟合(=aD),经回归拟合得出各林龄尾巨桉不通器官及整株生物量的拟合方程(表4)经统计学检验,各林龄尾巨桉的树干、树枝、叶片、树根和总生物量数学模型的相关系数在0.450 ~ 0.989 之间,检验均达到极显著水平(<0.01),说明拟合的方程可用于尾巨桉乔木层生物量的估测。
2.3 不同林龄尾巨桉乔木层生物量分配特征
尾巨桉随着林龄的增加,固碳量显著增加,由表5可知:2 a生、6 a生及10 a生尾巨桉林分总生物量分别为47.18、85.09和254.36t·hm-2,差异均达到了显著水平(<0.05),其中 10 a生极显著大于2 a生和6 a生。树干和树根的变化规律与总生物量相同,但叶片、树枝和树皮的生物量,2 a生与6 a生差异不显著,但均极显著小于10 a生。不同林龄尾巨桉各器官的生物量所占比例均是:树干>树根>树枝>树皮>叶片,且随着林龄的增加,树干生物量所占比例逐渐增大、叶片和树皮逐渐减小、树枝先减小后增大,而树根所占比例先增大后减小。

表4 尾巨桉各器官单株生物量估算模型
注:**表示<0.01

表5 不同林龄尾巨桉人工林乔木层各器官生物量分配
注:1.同列不同大写字母表示差异极显著(<0.01),不同小写字母表示差异显著(<0.05);2.括号内数据为各器官占总生物量的比例
3 结论与讨论
林木根系具有固定和吸收土壤水分和养分的作用,是提供维持林木生命活动的物质和能量的重要器官[14]。不同大小根系其分工不同,其中小根与中根主要起吸收水分和养分的作用,而大根主要起固定作用。本文通过研究发现:尾巨桉各林龄不同径级个体间细根及中根生物量差异不大,而大根随着径级的增加显著增大,随着林龄的增加,细根、中根和大根均有不同程度的增加,其中大根的增加量显著大于细根和中根,大根生物量随着林龄的增大,占整株根系生物量的比例迅速增加,10 a生大根比例达94.8%,可见2 a生到10 a生的尾巨桉处于快速生长期,更多的大根分配,有助于固定林木,以支撑地上部分高生长的需要。
对不同林龄尾巨桉人工林的生物量分配研究结果表明:随着林龄的增加,林分生物量显著增加,其中2 ~ 10 a生尾巨桉人工林的生物量为47.18 ~ 254.36 t·hm-2,但不同器官的生物量对林龄的响应并不一致,其中2 ~ 6 a生的树枝和叶片生物量间差异不显著,这可能是由于2 ~ 6 a生时,大量的自然整枝现象导致的。不同器官生物量占比例变化也不一致。不分林龄的将各器官生物量与胸径进行模型拟合[15-16],这样会明显降低人工林生物量的估算精度。本文对不同林龄的各个器官及整株生物量分别于胸径做了模型拟合,为提高桉树人工林的乔木层生物量估算精度提供理论支持。
[1] 汪珍川,杜虎,宋同清,等.广西主要树种(组)异速生长模型及森林生物量特征[J].生态学报,2015,35(13):4462-4472.
[2] WOODWELL G M, WHITTAKER R H, REINERS W A, et al. The biota and the world carbon budget[J].Science, 1978, 199(4325):141-146.
[3] 李根,周光益,吴仲民,等.南岭小坑木荷群落地上生物量[J].林业科学, 2012, 48(3):143-147.
[4] 韩新生,邓莉兰,王彦辉,等.六盘山叠叠沟华北落叶松人工林地上生物量的坡面变化[J].林业科学,2015,51(3):132-139.
[5] GENET A,WEMSDORFER H,JONARD M,et al. Ontogeny partly explains the apparent heterogeneity of published biomass equations for Fagus sylvatica in central Europe[J].Forest Ecology and Management,2011, 261(7):1188-1202.
[6] BRUNNER I ,GODBOLD D L. Tree roots in a changing world[J]. Journal of Forest Research,2007,12(2):78-82.
[7] 贾呈鑫卓,李帅锋,苏建荣,等.思茅松人工林根系特征与生物量分配[J].生态学杂志,2017,36(1):21-28.
[8] RONALD L, HENDRICK P .The dynamics of fine root length, biomass, and nitrogen content in two northern hardwood ecosystems[J].Canadian Journal of Forest Research,1993,23(12):2507-2520.
[9] 谢耀坚.真实的桉树[M].北京:中国林业出版社,2015.
[10] 何有才,唐建维,段文贵,等.西双版纳热带季节雨林的生物量及其分配特征[J].植物生态学报,2007,31(1):11-22
[11] 王维枫,雷渊才,王雪峰,等.森林生物量模型综述[J].西北林学院学报,2008,23(2):58-63.
[12] 任海,彭少麟.鹤山马占相思人工林的生物量和净初级生产力[J].植物生态学报,2000,24(1):18-21.
[13] 明安刚,贾宏炎,陶怡,等.桂西南28 a生米老排人工林生物量及其分配特征[J].生态学杂志,2012,31(5):1050-1056.
[14] 燕辉,刘广全,李红生.青杨人工林根系生物量、表面积和根长密度变化[J].应用生态学报,2010,21(11):2763-2768.
[15] 张利丽,王志超,陈少雄,等.不同林龄尾巨桉人工林的生物量分配格局[J].西北农林科技大学学报(自然科学版),2017,45,(6):61-68.
[16] 付威波,彭晚霞,宋同清,等.不同林龄尾巨桉人工林的生物量及其分配特征[J].生态学报,2014,34(18):5234-5241.
Root Characteristics and Biomass Allocation ofPlantations of Different Ages in Leizhou Peninsula
ZHANG Xuede1, WU Yong1, HUANG Yuchao1, HUANG Sheng1, ZHANG Mingsi1, SU Han1, YAN Binghui1, WANG Zhichao2,3
(1.,,,; 2.,;3.)
In view of the difficulties of estimating biomass ofplantations and the low precision of estimates obtained, a study was carried out to examine root biomass, above-ground biomass and the biomass distribution in plantations of different ages, and then model biomass measures to the variable of diameter at breast height (DBH). Development of such a mathematical model of biomass components ofplantations provides support for accurate calculation of arbor layer carbon storage in eucalypt plantations.The result shows thatwith increasing DBH, the biomass of fine roots and mid-sized roots at different plantation ages shows little difference, while the large roots increase significantly. With increasing plantation age, the proportion of large roots increases rapidly while the proportion of fine roots and middle roots gradually decreases.The amount of biomass allocated to different tree component organs inplantations followed the order: stem > tree root > branch > bark > leaf. The total biomass of 2-year-old, 6-year-old and 10-year-oldplantations were 47.18 t·hm-2, 85.09 t·hm-2and 254.36 t·hm-2respectively.The correlation (2) between biomass components and DBH across ages ranged between 0.45 and 0.99, depending on the component organ; all correlations were highly significant indicating that the regression model equations were accurate for estimation of biomass offrom tree DBH.
; plantations; root characteristics; biomass; regression model
S718.5
A
10.13987/j.cnki.askj.2019.03.004
广西科技重大专项(桂科AA17204087-9);国家重点研发计划课题(2016YFD0600505, 2016YFD0600504); 广东湛江桉树林生态系统国家定位观测研究站运行项目(2019132141)
张学德(1979—),男,本科,工程师,主要从事森林培育研究,E-mail:zxd197979@sina.com
王志超(1988― ),男,硕士,助理研究员,主要从事生态水文研究,E-mail:wzc2254@163.com
猜你喜欢
广西林业科学(2022年4期)2022-11-07 01:50:34
林业调查规划(2022年3期)2022-06-13 09:31:04
广西林业科学(2021年6期)2022-01-21 07:39:10
防护林科技(2020年10期)2020-12-22 11:46:42
儿童故事画报·发现号趣味百科(2019年9期)2019-02-02 04:12:19
环球时报(2019-01-03)2019-01-03 09:06:44
防护林科技(2016年9期)2016-09-27 01:34:21
防护林科技(2016年5期)2016-09-05 01:19:43
广西林业科学(2016年2期)2016-03-20 05:53:22
广西林业科学(2016年4期)2016-03-16 05:44:51
