
油用牡丹‘凤丹'种子内源抑制物研究
2019-10-23 07:51:52朱铭玮邹雨婷李永荣李淑娴
西南林业大学学报 2019年6期
朱铭玮 邹雨婷 李永荣 李淑娴
( 1. 南京林业大学南方现代林业协同创新中心,江苏 南京 210037;2. 南京绿宙薄壳山核桃科技有限公司,江苏 南京 210014)
牡丹(Paeonia suffruticosa)是毛茛科(Ranunculaceae)、芍药属(Paeonia)多年生落叶灌木,在我国分布范围较广。油用牡丹是牡丹组中结实力强,能够生产种子及加工食用牡丹籽油的一个特殊类型。2011年3月以牡丹籽仁为原料提炼出的牡丹籽油被卫生部批准为新资源食品[1]。对牡丹籽油的成分进行分析后,人们发现其主要以亚麻酸、油酸、亚油酸、棕榈酸和硬脂酸为主的37种成分组成,其中不饱和脂肪酸占总量83.42%[2],特别籽油中α-亚麻酸含量较高,是橄榄油的140倍[3],因此牡丹籽是一种适宜开发利用的良好油料资源,具有广阔的发展前景。
生产上油用牡丹多用种子进行繁殖,但是该种子存在休眠习性,需要经过6个月以上的层积才能正常萌发,这给播种繁殖带来了很多不便。以往的研究表明,造成凤丹(P. ostill‘Fengdan')种子休眠的原因是种胚形态发育不完全以及种子内存在抑制物引起的生理休眠,是综合休眠[4]。宋会兴等[5]认为四川牡丹种子(P. decomposita)的种皮、胚乳中含有抑制白菜种子萌发及幼苗生长的物质,并且随着该物质浓度的增加,抑制作用更为显著,但是对造成其休眠的抑制物的种类却未开展研究。本研究以油用牡丹的主栽品种群——凤丹种子为材料,从内源抑制物质角度讨论其休眠原因,通过系统溶剂分离法对凤丹种皮和种胚的浸提液进行分离,并对各提取相进行生物测定,结合气相色谱-质谱联用(GC-MS)技术分析造成凤丹种子休眠的内源抑制物的成分,探讨内源抑制物与种子休眠的关系,以期为种子生物学及休眠机理研究提供参考。
1 材料与方法
1.1 实验材料
油用牡丹‘凤丹'种子,从安徽亳州采后置于5 ℃冰箱储藏;市售白菜种子(矮脚黄),纯度≥95.0%,发芽率≥85%。
1.2 实验方法
1.2.1 凤丹种子内源抑制物的提取
称取饱满凤丹种子50 g,将其分为种皮、种胚2个部分,用粉碎机磨碎后,置于500 mL锥形瓶中,再加入400 mL的80%甲醇溶液,于4 ℃条件下密闭浸提,其间多次取出搅拌,使浸提充分。24 h后过滤浸提液,再向锥形瓶中加入400 mL的甲醇溶液,连续浸提3次,最后将3次浸提液合并。在60 ℃下减压蒸除甲醇和部分水,最后将浓缩液定容至200 mL。
1.2.2 凤丹种子内源抑制物的初步分离
采用系统溶剂分离法[6]对上述种子各部位的甲醇提取液进行初步分离,将得到的石油醚相、乙醚相、乙酸乙酯相、甲醇相和水相浸提液,分别置于旋转蒸发仪上进行浓缩蒸发,然后定容为200 mL,置于4 ℃冰箱中保存。
1.2.3 凤丹种子分离相的生物活性测定
以白菜种籽为材料,对上述各相提取的溶液进行生物测定,具体为:取各相浸提液5 mL,分别加入铺有一层滤纸的培养皿中,待有机溶剂挥发后,加5 mL蒸馏水,对照加入相同体积的蒸馏水。随机数取白菜籽50粒,3次重复,置于培养皿中,随后将培养皿放在25 ℃的恒温光照培养箱内进行发芽实验,当胚根伸长至1 cm时即认为该粒种子已经发芽,每8 h统计1次,连续3 d种子发芽粒数不超过1%时即认为发芽结束,最后计算各处理的发芽率。
1.2.4 种皮、种胚分离相的GC-MS鉴定
分别取凤丹种子的种皮、种胚各有机相浓缩液100 mL,在旋转蒸发仪上减压浓缩蒸干,得到浓缩的干物质,然后再用相应的有机溶剂溶解定容至3 mL,即得到各有机相提取物的浓缩样品。上述浓缩样品在清华大学分析中心质谱室进行鉴定,鉴定条件如下:气质仪器:GC-MS QP2010 ultra(Shimadzu,Kyoto,Japan)。气质分析方法:柱子 Rxi-5Sil MS(Restek,USA,30 m×0.25 mm×0.25 μm),进样口300 ℃,分流进样分流比20∶1,载气为高纯氦气,流速为1 mL/min。升温程序:50 ℃保持 5 min,以 5 ℃/min涨至 300 ℃保持30 min。质谱条件:离子源:EI;离子源温度:200 ℃;接口温度:250 ℃,溶剂切除时间:3 min;扫描45~500 m/z。将GC-MS鉴定出的各个成分的质谱图与标准库谱图进行核对,确定各个物质的名称、分子式、分子量和相对含量等。
2 结果与分析
2.1 种皮和种胚各分离相的生物测定
凤丹种子的种皮、种胚各相提取液对白菜籽发芽的影响见表1。从表1可知:种子各部位、各相提取液对白菜籽发芽均有一定的抑制作用,且除石油醚相及种胚乙酸乙酯相外,其他各相均使白菜籽发芽显著下降,但种子部位不同、提取相不同,其对白菜籽发芽率的抑制程度有所不同。种皮提取液中,乙醚相的抑制作用最强,处理后白菜籽几乎不能发芽,发芽率由对照的86.7%下降到0.7%;其次是乙酸乙酯相,发芽率下降到6.7%,降低了80.0%;甲醇相、水相处理后,发芽率也有较大程度的下降,分别较对照下降了52.0%、37.4%。种胚部位,仍是乙醚相的抑制作用最强,使白菜籽发芽率显著下降到3.3%,甲醇相、水相处理后,白菜籽发芽率分别显著下降到44.7%、56.0%,乙酸乙酯相处理后白菜籽的发芽率为83.4%,较对照差异不显著。同一提取相中,不同部位对白菜籽发芽的影响程度也不尽相同,种皮乙酸乙酯相和甲醇相的抑制作用显著高于种胚,特别是乙酸乙酯相,种皮提取液处理后仅有6.7%的白菜籽能够发芽,但种胚的提取液却使白菜籽发芽率下降不显著;乙醚相、水相中,种皮、种胚部位的抑制作用差异不显著;石油醚相中则是种胚的抑制作用略强于种皮,但二者均较对照差异不显著。总的来说,同一提取相,种皮部位的抑制作用略强于种胚。种子不同部位各相生物测定的发芽率结果,均是乙醚相的抑制作用最强,因此凤丹种子的内源抑制物可能主要存在于乙醚相中。
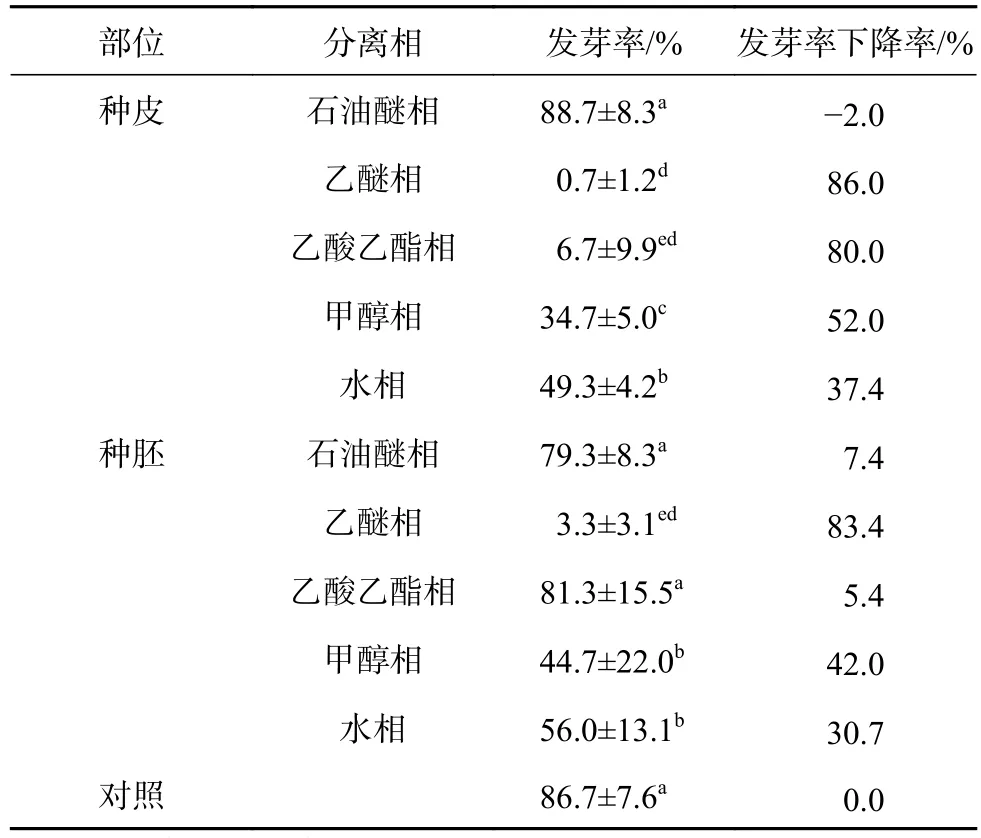
表1 凤丹种子不同部位各相提取物对白菜籽发芽的影响Table 1 Effects of extracts of P . ostii ‘Fengdan' seeds to the germination of cabbage seeds
2.2 凤丹种皮各分离相GC-MS鉴定结果
以抑制作用最强的乙醚相为例,经GC-MS分析得到的总离子流色谱图见图1,通过质谱系统检索并与标准谱图进行核对,筛选总离子流色谱图中峰面积较大、相似度大于80%的有机化合物,将质谱图结果与计算机检索谱库图比对后,得到了吻合性较好的14种物质,见表2。由表2可知,种皮乙醚相中14种有机物主要为酚类物质3种(双酚F、苯酚、2,6-二叔丁基对甲酚),相对含量为12.3%;醇类物质2种(beta-谷甾醇、羽扇豆醇),相对含量为10.1%;酸类物质2种,棕榈酸、反油酸,相对含量为6.4%;酰胺类物质1种,芥酸酰胺,含量为1.4%,其他物质2种,未知物质4种。
图1 凤丹种皮中乙醚相提取物的离子流程Fig. 1 The ion flow of extract with aether solution in seed coat of P. ostii ‘Fengdan'
按照相同的方法,筛选出种皮其他相中的有机物,见表3。由表3可知,种皮乙酸乙酯相中共鉴定出17种有机化合物,主要为酚类物质,其总相对含量为20.3%,主要包括双酚F、苯酚、邻苯二酚、间苯三酚,另外还有有机酸类、酰胺类物质,如棕榈酸、苯甲酸、反油酸、芥酸酰胺、油酸酰胺等,还含有少量的6,7-二羟基香豆素[7](1.1%)。种皮甲醇相也鉴定出17种有机物,主要为酚类和有机酸物质,其相对含量较高的为苯甲酸(13.2%)、邻苯二酚(10.5%)、苯酚(9.4%)、芥酸酰胺(2.6%)等。种皮石油醚相共鉴定出24种物质,主要是烷类物质其总相对含量为20.4%,还含有棕榈酸(3.7%)、芥酸酰胺(3.3%)等物质。
2.3 种胚分离相的GC-MS鉴定结果
本次筛选出种胚各有机相中的化合物见表4。种胚乙醚相、甲醇相、乙酸乙酯相和石油醚相分别鉴定出了34、26、24、26种有机化合物,各相鉴定出的物质种类数比种皮部位稍多些。种胚乙醚相中鉴定出的有机化合物相对含量较高的主要是有机酸类(19.9%)及其酯类化合物(18.5%),有机酸类主要为苯甲酸、油酸、棕榈酸、硬脂酸等,酯类物质可能是由于提取及测试过程中某些酸类物质发生了酯化作用造成的,因此有机酸的实际相对含量可能还要高些;另外,该有机相内虽然含有2,6-二叔丁基对甲酚、芥酸酰胺、苯酚等物质,但是相对含量较低。甲醇相主要为酚类和有机酸类物质,酚类物质主要为苯酚,其相对含量为8.3%,苯甲酸及其酯化物的总含量为8.8%。乙酸乙酯相中鉴定出的主要为酯类物质,存在于该相的酚类物质为苯酚、2,6-二叔丁基对甲酚,有机酸类为反油酸、硬脂酸、苯甲酸和棕榈酸,酰胺类为芥酸酰胺和油酸酰胺,但酚类、有机酸类和酰胺类物质含量都只在1%左右。石油醚相中鉴定出的有机化合物主要为酯类(20.1%)、烷类(19.1%)和醇类(13.8%)物质,该有机相中也存在甲基苯、酰胺类以及有机酸类物质,但是其含量很低。

表2 凤丹种子种皮乙醚相中的化合物及相对含量Table 2 Compounds and their relative contents in aether extract of P . ostii ‘Fengdan' seed coat

表3 凤丹种皮各分离相的化合物及相对含量Table 3 Compounds and their relative contents in seed coat extracts of P . ostii ‘Fengdan'

表4 凤丹种胚各分离相的化合物分类及相对含量Table 4 Compounds and their relative contents in decoated seeds extracts of P . ostii ‘Fengdan'
3 结论与讨论
凤丹种子萌发需要较长时间,这与其种子具有深休眠特性有关[8]。本研究从抑制物角度,探讨引起凤丹种子生理休眠的原因。以往的研究表明,内源抑制物的存在是导致许多种子休眠的主要原因之一,如黄精(Polygonatum sibiricum)[9]、乌桕(Sapium sebiferum)[10]、南方红豆杉(Taxus chinensisvar.mairei)[11]等种子中都含有内源抑制物。对凤丹种子各有机相的生物测定结果同样表明,种子中内源抑制物质的存在可能是导致其休眠的原因之一。
要确定内源抑制物的存在,通常采用系统溶剂法对种子的甲醇相提取液进行分离,并结合各有机相对白菜籽等非休眠植物种子的发芽实验进行生物测定。尚旭岚等[12]采用系统溶剂法对青钱柳(Cyclocarya paliurus)种子的浸提液进行分离,通过生物测定发现抑制物质主要存在于乙醚相中;史锋厚等[13]对南京椴(Tiliam iqueliana)种子的研究发现,果皮中的抑制物质主要存在于乙醚相和甲醇相中,而种子中的抑制物质主要在甲醇分离相中;张艳杰[14]对南方红豆杉种子的研究中发现,抑制物质主要存在于种子的乙醚相和甲醇相中。本研究中,对各有机相生物测定结果比较后发现,种皮和种胚的乙醚相的抑制作用最强,这和上述研究结果相似,因此该种子乙醚相鉴定出的物质可能与其生理休眠有密切关系。
以往的研究表明,酚类、有机酸类是植物种子中常见的抑制物质,它们通过阻碍种子吸水进程、抑制呼吸作用、抑制生物酶的活性以及阻碍胚根生长等方式,对种子萌发产生抑制作用[15-17]。近年来,发芽抑制物质的分离与鉴定已成为种子休眠机理研究的重要内容。
Eiuhellin[18]认为,酚类物质能够抑制种子萌发所需的关键酶的代谢,最终对种子的萌发有影响。酚类化合物对植物生长也具有较强的抑制作用,其对水葫芦的抑制作用主要表现在使水葫芦叶片失水干枯,根毛变黑,易脱落[19]。钱存梦[20]对乌桕种子休眠的研究中证实了2,6-二叔丁基对甲酚是重要的内源抑制物;韩东等[21]发现邻苯二酚对西洋参(Panax quinquefolius)种子萌发具有重要的抑制作用,刘序等[22]在香榧种子抑制其发芽和生长的内源物筛选研究中,发现邻苯二酚对白菜种子的发芽率和幼苗生长的抑制作用最强。本次GC-MS检测结果表明,对白菜籽发芽有显著影响的种皮乙醚相、甲醇相以及种胚的甲醇相中,酚类物质的相对含量较高,分别为12.3%、22.6%、9.4%,并且这几个相中同样含有苯酚、2,6-二叔丁基对甲酚和邻苯二酚等酚类内源抑制物质,因此酚类物质可能与该种子休眠有关。另外,这几个相中双酚F的相对含量也比较高,该物质在以往的研究中被证明对生物会有毒害作用,如潘芳[23]、任文娟等[24]发现双酚F对斑马鱼幼苗具有一定的毒害作用,但其对种子休眠是否有影响还需要进一步研究。
有机酸类物质也是已知的内源抑制物质,王艳华[25]认为苯甲酸、棕榈酸、硬脂酸等是大山樱(Prunus sargentii)种子休眠的重要内源抑制物质。李龙秀等[26]研究发现苯甲酸对萝卜种子萌发及幼苗生长有明显的抑制作用,表现为高浓度时抑制,低浓度时抑制作用减弱的特点。本次种皮的乙醚相、甲醇相以及种胚的乙醚相、甲醇相中,有机酸相对含量也比较高,分别为6.4%、16.9%、19.9%、4.1%,而且有机酸在实验过程中容易酯化,因此其实际相对含量可能还要高些。本次抑制作用较强的种胚乙醚相中,苯甲酸的相对含量高达17.4%,同时还含有一定量的棕榈酸和硬脂酸,这些酸类物质可能是该相对白菜籽发芽抑制较强的重要原因。
徐敬东[27]在对紫藤(Wisteria sinensis)的研究中发现,提取物芥酸酰胺对真菌活性具有一定的抑制作用,蔡梦可[28]对花梨木(Peterocarpesspp.)类木材的耐腐性机理的研究中也发现了芥酸酰胺,认为其对真菌的活性有一定抑制作用。酰胺类物质中的油酸酰胺也已被证实能够诱导动物产生生理性睡眠[2]。本次各有机相中,除了酚类和有机酸类物质,还含有一定量的酰胺类物质,因此酰胺类物质对牡丹种子萌发的影响需要深入研究。
GC-MS结果分析时还发现,凤丹种皮乙酸乙酯相中相对含量最多的为酚类,占20.7%,而种胚则以酯类物质为多,酚类物质的相对含量只有5.7%;而且种皮中有机酸类和酰胺类物质的相对含量也高于种胚。表1中种皮乙酸乙酯相对白菜籽发芽的影响显著强于种胚(二者的发芽率分别是6.7%,81.3%),造成这一显著差异的原因可能也是由于种皮中拥有较多的酚类、有机酸类、酰胺类物质。
本实验在种皮、种胚石油醚相中鉴定出的有机化合物主要为烷类、酯类、醇类等物质,结合表1的生物测定结果,这些物质可能不是造成凤丹种子休眠的重要内源抑制物质。此外,种皮石油醚相中有机酸的相对含量达7.0%,但其生物测定的抑制性不高,该相相对含量较高的是棕榈酸,因此酚类、有机酸类和酰胺类物质中,哪些物质与凤丹种子休眠的有关还需进一步研究。表1生物活性测定结果中,水相对白菜籽发芽也具有一定的抑制作用,但由于仪器的限制,水相中具体的抑制物质并未能确定出来。
综上所述,内源抑制物的存在是凤丹种子生理休眠的原因之一,初步分析发现,其内源抑制物的主要类型可能是酚类、有机酸类及酰胺类物质。本研究中乙醚相的抑制作用最强,其可能含有影响种子休眠的主要抑制物质。种皮抑制物的主要成分是酚类物质,可能是双酚F、苯酚、邻苯二酚、2,6-二叔丁基对甲酚等;种胚的主要抑制物质是有机酸类物质,如苯甲酸、油酸、棕榈酸、硬脂酸等;一些酰胺类物质可能也具有抑制作用。本实验被鉴定出的物质中,有些物质的抑制作用已被证实,但其他物质是否也能抑制种子萌发以及和凤丹种子休眠的关系有待进一步证实。其中酚类、有机酸类和酰胺类物质,哪一类物质对凤丹种子生理休眠起主要作用,以及相互之间对种子生理休眠的作用关系也有待进一步研究。
猜你喜欢
特产研究(2022年6期)2023-01-17 05:06:02
粮油与饲料科技(2022年2期)2022-11-24 13:12:07
CHINA TODAY(2022年4期)2022-11-22 12:43:48
花卉(2021年17期)2021-09-15 08:51:16
植物资源与环境学报(2020年4期)2020-08-10 05:23:14
天然产物研究与开发(2018年3期)2018-05-07 06:38:52
天然产物研究与开发(2018年3期)2018-05-07 06:38:43
福建中医药(2017年6期)2018-01-09 06:49:54
北方牧业(2016年6期)2016-12-17 14:04:49
西南农业学报(2016年4期)2016-05-17 05:42:14
