
剑麻感染烟草疫霉菌后几种重要防御酶活性的变化
2019-10-22 08:36郑金龙易克贤习金根黄兴吴伟怀高建明张世清陈河龙贺春萍粱艳琼陆英
中国麻业科学 2019年5期
郑金龙,易克贤*,习金根,黄兴,吴伟怀,高建明,张世清,陈河龙,贺春萍*,粱艳琼,陆英
(1.中国热带农业科学院环境与植物保护研究所/农业农村部热带作物有害生物综合治理重点实验室,海南海口571101;2.中国热带农业科学院热带生物技术研究所/农业农村部热带作物生物技术重点开放实验室,海南海口571101)
剑麻是我国热区乃至全球最重要的经济作物[1],其不仅是主要的热带纤维原料[2],同时也是一种重要的生物质能源植物[3]。我国剑麻产业面临良好发展机遇的同时,也面临严峻的挑战。40多年来,我国种植剑麻的主栽品种为从国外引进的杂交种H.11648,此品种虽然高产,但易感斑马纹病。斑马纹病是当前剑麻生产中最严重的毁灭性病害之一,其病原菌主要为烟草疫霉菌(Phytophthora nicotianaevan Breda de Haan)[4-6]。目前,防治该病的主要措施是使用杀菌剂和种植抗病品种。但由于该病潜伏期长、致病机理不明以及剑麻植株的蜡质层、鳞片极大地阻碍了药剂的顺利渗入等原因,导致杀菌剂的防治效果差;同时由于剑麻是多年生植物,生长周期长达十年以上,抗原十分有限,给抗病育种带来了很大困难。
植物受到病原物侵染而发生主动防卫反应时,体内一系列防御酶系会发生某些变化[7],它们相互协同发挥作用,不同植物甚至同种植物的不同品种、不同胁迫因子等条件下酶活性的变化与抗病性也不相同,甚至相反[8]。近年来,学者们对防御酶活性变化与植物抗病性的关系进行了广泛的研究。魏颖颖[9]对烟草接种CMV前后CAT活性与抗病性的关系进行了研究。陈利锋等[10]利用禾谷镰孢菌接种不同抗赤霉病的小麦品种,比较了接种后不同时间穗组织中超氧化物歧化酶(SOD)和过氧化氢酶(CAT)的活力变化。邵登魁等[11]研究了白菜型冬油菜在感白粉病品种全生育期和全病程中超氧化物歧化酶(SOD)、过氧化物酶(POD)、多酚氧化酶(PPO)的活性变化。而关于剑麻防御酶活性的研究,目前仅有少数几篇报道[12-15]。本文以剑麻易感病品种H.11648为材料,在接种烟草疫霉菌前后,分别测定剑麻叶片中苯丙氨酸解氨酶(PAL)、超氧化物歧化酶(SOD)、多酚氧化酶 (PPO)、过氧化氢酶 (CAT)、过氧化物酶 (POD)、几丁质酶(CHT)、&-1,3葡聚糖酶(GLU)等7种酶的活性,以了解剑麻感染烟草疫霉前后几种重要防御酶活性的变化,并分析病菌对剑麻生理反应的影响,旨在为进一步研究烟草疫霉菌与剑麻之间的互作机理奠定生理生化基础,这对高效选育抗病品种和持久利用剑麻的抗病性具有重要的理论和实践意义。
1 材料与方法
1.1 试验材料
剑麻(H.11648)盆栽苗,株高50 cm;烟草疫霉菌强致病力菌株CH0008由中国热带农业科学院环境与植物保护研究所分离、鉴定和保存。
1.2 试验方法
1.2.1 接种前酶活性测定
取14株生长基本一致的盆栽苗,其中7株作为对照,7株作为处理组,每个处理3次重复。从最下部开始,每株各割取一片完好的叶片,带回实验室,洗净、晾干后,及时测定其PAL、SOD、PPO、CAT、POD、CHT和GLU等7种酶活性,酶活性测定方法均按照酶活试剂盒(苏州科铭生物技术有限公司)步骤测定。
1.2.2 菌株的制备及接种
将保存的烟草疫霉菌用PDA培养基活化,置于28℃生化培养箱中活化培养7 d,以备接种,从麻苗最下部的叶片开始选择完好叶片全部接种,接种位置距叶片基部10 cm,接种时先用75%酒精消毒,刺伤表皮组织,将直径6mm的菌块带菌丝一面贴于伤口处(对照的3株,将同样大小的空白 PDA贴于伤口处),用棉花保湿,置于28℃环境条件下培养[14]。
1.2.3 接种后酶活性测定
分别于接种后24、48、72、96、120 h取剑麻幼叶提取粗酶液。检测不同时间剑麻体内相关防御酶的活性变化。叶片取样及酶活性测定方法同1.2.1。
1.2.4 统计学分析
试验所得数据均采用JMP10软件进行方差分析,并用杜凯氏差距(Tukey HSD)检验法进行多重比较。
2 结果与分析
2.1 对苯丙氨酸解氨酶(PAL)活性的影响
由图1可知,接种烟草疫霉菌后各个时间段的PAL活性均比对照(CK)高,说明烟草疫霉菌可以诱导剑麻叶片PAL活性增强。对照组剑麻叶片PAL活性在0~120 h之间没有明显变化。接种烟草疫霉菌的剑麻叶片,PAL活性在0~120 h之间随着时间的推移先呈逐步上升的趋势,96 h时达到峰值78.888 U/g,随后随着时间的推移呈逐步下降的趋势。

图1 接种烟草疫霉菌前后剑麻叶片PAL活性变化Fig.1 Changes of PAL activity in sisal leaves before and after inoculation with Phytophthora nicotianae
2.2 对超氧化物歧化酶(SOD)活性的影响
如图2所示,剑麻叶片受烟草疫霉菌侵染后,SOD活性在0~120 h之间随着时间的推移先呈下降的趋势,48 h时降到最低值50.959 U/g,且低于CK,说明剑麻叶片受烟草疫霉菌侵染前期SOD活性受到抑制;随后随着时间的推移而逐步上升,120 h时达到最高值88.013 U/g。对照组的剑麻叶片,SOD活性在0~120 h之间没有显著性变化。
2.3 对多酚氧化酶(PPO)活性的影响
如图3所示,剑麻叶片受烟草疫霉菌侵染后,PPO活性在0~120 h之间随着时间的推移呈现先上升后下降的趋势,96 h时达到最高值156.801 U/g,随后随着时间的推移而逐步下降,且48~120 h之间,处理组PPO活性显著高于CK,说明烟草疫霉菌可以诱导剑麻叶片PPO活性增强。对照组的PPO活性随着时间的推移没有显著性变化。
2.4 对过氧化氢酶(CAT)活性的影响
如图4所示,剑麻叶片受烟草疫霉菌侵染后,CAT活性在0~120 h之间随着时间的推移呈现先上升后下降的趋势,48 h时达到最高值1267.863 U/g,随后随着时间的推移而逐步下降,72 h时达到最低值0.259 U/g,随后没有显著性变化,说明烟草疫霉菌在侵染前期先诱导剑麻叶片CAT活性增强,随后又抑制了其酶活性。对照组的CAT活性随着时间的推移基本保持不变。

图2 接种烟草疫霉菌前后剑麻叶片SOD活性变化Fig.2 Changes of SOD activity in sisal leaves before and after inoculation with Phytophthora nicotianae

图3 接种烟草疫霉菌前后剑麻叶片PPO活性变化Fig.3 Changes of PPO activity in sisal leaves before and after inoculation with Phytophthora nicotianae

图4 接种烟草疫霉菌前后剑麻叶片CAT活性变化Fig.4 Changes of CAT activity in sisal leaves before and after inoculation with Phytophthora nicotianae
2.5 对过氧化物酶(POD)活性的影响
如图5所示,剑麻叶片受烟草疫霉菌侵染后,在0~120 h之间,处理组各个时间段的POD活性均比CK高,说明烟草疫霉菌可以诱导剑麻叶片POD活性增强。POD活性随着时间的推移呈现先上升后下降的趋势,96 h时达到峰值1854.583 U/g,随后随着时间的推移而逐步下降。对照组的POD活性随着时间的推移没有显著性变化。

图5 接种烟草疫霉菌前后剑麻叶片POD活性变化Fig.5 Changes of POD activity in sisal leaves before and after inoculation with Phytophthora nicotianae
2.6 对几丁质酶(CHT)活性的影响
如图6所示,剑麻叶片受烟草疫霉菌侵染后,CHT活性在0~120 h之间随着时间的推移呈现先下降后上升的趋势,而CK的CHT活性随着时间的推移没有显著性变化。其中,24~72 h期间,处理组CHT活性均低于CK,96 h以后处理组CHT活性高于CK,且120 h时达到显著差异。

图6 接种烟草疫霉菌前后剑麻叶片CHT活性变化Fig.6 Changes of CHT activity in sisal leaves before and after inoculation with Phytophthora nicotianae
2.7 对&-1,3葡聚糖酶(GLU)活性的影响
如图7所示,剑麻叶片受烟草疫霉菌侵染后,GLU活性在0~120 h之间随着时间的推移呈现先下降后上升的趋势,而CK的GLU活性随着时间的推移没有显著性变化。24~48 h期间,处理组GLU活性低于CK,其中24 h时呈显著性差异;72~120 h期间,处理组GLU活性显著高于CK。
3 讨论
剑麻叶片在用针刺伤后,喷无菌水,不接种烟草疫霉菌,其PAL、SOD、PPO、CAT、POD、CHT和GLU等7种酶活性在0~120 h期间没有显著性变化,说明针刺对剑麻表皮造成的微小伤口不足以对剑麻内部的生理生化等新陈代谢产生影响,因此,对叶片中酶活性的影响也较小。这与赵艳龙等[14]的研究结果相一致。
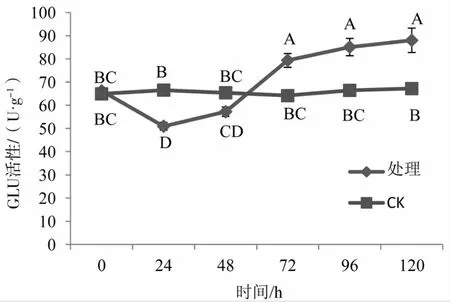
图7 接种烟草疫霉菌前后剑麻叶片GLU活性变化Fig.7 Changes of GLU activity in sisal leaves before and after inoculation with Phytophthora nicotianae
在本研究中,剑麻受烟草疫霉菌侵染后,PAL、PPO、CAT和POD的活性在0~120 h期间均高于对照,且随着时间的推移均呈现先上升后下降的趋势,这与其他文献报道的防御酶活性提高可增强植物抵抗包括病害在内逆境的能力这一论点相一致[16-18]。说明在烟草疫霉菌侵染的剑麻叶片中PAL、PPO、CAT和POD等酶活性的增加与寄主抵抗病原入侵有一定的相关性,可以将其活性作为植物抗病的一个生理指标。但与赵艳龙等[14]的研究结果不尽相同,这可能与取样的时间、剑麻植株种植区域、病原菌的来源及致病力大小等有关,需要后续实验进一步验证。
剑麻叶片受烟草疫霉菌侵染后,SOD活性在0~120 h之间随着时间的推移呈现先下降后上升的趋势,这与朱友林等[19]的研究结果相一致,说明病原菌侵染初期,SOD活性下降与抗病系细胞质膜透性迅速增加、细胞膜结构较早变化、细胞过敏性坏死以及DIMBOA迅速大量释放等早期抗病性反应有密切关系,而病原菌侵染中、后期SOD活性较大幅度上升,与抗病系在病程中、后期限制病斑扩展的能力有关。
GLU和CHT作为在植物保卫反应中起主要作用的细胞壁水解酶,受到许多学者的关注[20]。左豫虎等[21]的研究表明,大豆疫霉菌能诱导大豆GLU和CHT的活性增强,但2种酶在积累速度和幅度上,抗病品种和感病品种有显著的差异。史娟等[22]的研究表明,葡萄霜霉病菌能诱导葡萄叶片GLU和CHT活性增高,且两种酶活性增高的速度和幅度与品种的抗病性呈正相关。在本研究中,接种烟草疫霉菌的剑麻叶片,GLU和CHT酶活性在24~120 h之间随着时间的推移呈逐步上升的趋势,这与前人的研究结果相一致[21-22],说明剑麻烟草疫霉菌能诱导剑麻叶片组织GLU和CHT的活性增强,同时也进一步证实了GLU和CHT在植物抗病中的重要作用。
本文以高感品种H.11648为研究材料,对接种病原菌前后几种防御酶的活性变化作了初步探讨。接种病原菌后,剑麻叶片中PAL、SOD、PPO、CAT、POD、CHT和GLU等7种酶活性均发生了变化,进一步验证了植物和病原菌相互作用中,植物体内防御酶活性发生变化是寄主的一种普遍性反应,和植物抗病性有一定的关系这一论点[23],这为研究酶活性与剑麻抗病性的关系奠定了一定基础,同时也为从分子水平上认识寄主植物与病原菌相互作用机制提供了理论依据。而有关病原菌侵染后各防御酶表达的分子机制以及是否可以将该类酶作为筛选抗性材料的生化指标,还需要进一步试验和研究。
猜你喜欢
热带作物学报(2022年7期)2022-08-06
当代水产(2022年1期)2022-04-26
作物学报(2022年4期)2022-02-10
中国农业科学(2021年23期)2022-01-14
纺织科技进展(2021年5期)2021-07-22
昆虫学报(2021年3期)2021-05-11
中国瓜菜(2020年8期)2020-09-26
热带农业工程(2019年6期)2019-12-22
世界热带农业信息(2018年6期)2018-03-05
星星·散文诗(2017年22期)2017-12-30
