
高粱TCP基因家族全基因组鉴定及表达分析
2019-10-21 08:01白雪婷李会云
河南农业科学 2019年10期
郑 玲,白雪婷,李会云
(洛阳师范学院 生命科学学院,河南 洛阳 471934)
TCP基因是3个不同物种的4个基因的缩写,分别是玉米中的TB1(Teosinte branched 1)基因,金鱼草中的CYC(Cycloidea)基因和水稻中的PCF1(Proliferating cell factors 1)和PCF2基因[1-3]。TCP是植物特有的转录因子家族,大多数TCP蛋白在N端有1段约60个氨基酸残基组成的与DNA结合的非典型bHLH(Basic-helix-loop-helix)保守区,又称为TCP结构域。此外,TCP蛋白还存在1个保守的R结构域,R结构域并不是所有的TCP转录因子共有的,它富含精氨酸、赖氨酸和谷氨酸等极性氨基酸,可以形成1个亲水性的ɑ螺旋[2,4]。
根据TCP基因家族结构域同源性,可将TCP基因家族分为ClassⅠ和ClassⅡ 2个类群。ClassⅡ的TCP结构域比ClassⅠ多了4个氨基酸残基[1]。ClassⅠ中最先确定的功能基因是水稻的PCF1和PCF2,因此,TCP ClassⅠ也被称为PCF(或TCP-P)[5]。研究发现,拟南芥AtTCP14基因参与调控种子的胚胎发育[6];AtTCP15基因不仅参与节间长度调控,还与叶片发育调控有关[7];AtTCP20基因参与调控植物生长发育、茉莉酸(JA)生物合成及叶片衰老[8-9];AtTCP22基因在叶片衰老中起负调控作用,并与AtTCP15基因存在功能冗余现象[10];AtTCP16基因在发育的小孢子中表达,在转基因植株中下调该基因可导致1/2的花粉异常[11];AtTCP11基因与维管束后生木质部导管分子的分化和形成有关[12];AtTCP17基因参与光信号传导和生长素合成从而调节细胞伸长[13]。
ClassⅡ类(TCP-C)[7]成员又可分为CYC/TB1和CIN (Cincinnata)2个亚类,CYC/TB1亚类中包含1段非保守的谷氨酸-半胱氨酸-谷氨酸的延伸结构[14];CIN亚类中包含有microRNA miR-319的靶基因[15]。CIN亚类起源于金鱼草(Antirrhinum)中的CINCINNATA,包含涉及侧生器官发育的基因;而CYC/TB1亚类(或ECE亚类)包含腋分生组织发育成花或侧枝的基因[16]。研究发现,玉米TB1基因是玉米顶端优势的决定因素之一,此外,TB1基因还参与调控玉米叶分生组织的分化,防止在低节点生成芽以及在高节点上生成雌花序[16];水稻TB1基因也参与调控水稻侧枝发育[17];TB1与其同源基因(AtTCP12、AtTCP18、SlTCP7、SlTCP9)都在腋芽中表达,调控侧生器官发育[18]。
高粱(SorghumbicolorL.)是禾本科高粱属植物,叶和玉米叶很像,具有圆锥形花序和红褐色籽粒。高粱是我国的重要经济作物之一,其籽粒是酿造茅台、竹叶青等诸多名酒的主要原材料,在未来的生物能源中也将发挥重要作用[19]。近年来,由于人们发现TCP基因在植物生长发育过程中起着重要的调控作用,其家族成员的全基因组鉴定和分析成为研究热点,目前已在多个物种中进行了研究,如大豆[20]、辣椒[21]、毛竹[22]、拟南芥[23]、番茄[24]、玉米[25]等,还未见关于高粱TCP 基因家族全基因组鉴定和分析的报道。为此,对高粱TCP家族基因进行鉴定和分析,并推测高粱TCP家族部分基因的功能,分析高粱TCP基因在不同器官和非生物胁迫下的表达情况,旨在为进一步研究TCP基因在高粱生长发育及逆境胁迫中的作用提供参考。
1 材料和方法
1.1 试验材料
高粱的TCP序列来自PlantTFDB(http://planttfdb.cbi.pku.edu.cn/)、Phytozome(http://phytozome.net/)数据库,拟南芥(Arabidopsisthaliana)TCP基因序列来自PlantTFDB网站; TCP基因在高粱各组织中的表达量数据来自qTeller(http://qteller.com/)网站。
1.2 试验方法
1.2.1 高粱TCP基因家族成员的鉴定 从Pfam 网站(http://pfam. sanger.ac.uk/)下载TCP基因家族种子序列(PF03634),并以PF03634为搜索序列在Phytozome数据库(http://phytozome.net/)中下载全部高粱TCP基因序列(E值小于1×10-10)。将下载的结果与PlantTFDB数据库(http://planttfdb.cbi.pku.edu.cn/)中下载的序列进行比对,去除冗余,得到候选基因。利用InterPro(http://www.ebi.ac.uk/interpro/)在线验证候选基因,鉴定其编码蛋白质是否含有TCP结构域。使用ExPASy(http//www.expasy.org/)对最终所筛选出的氨基酸序列进行在线分析,以确定蛋白质的氨基酸长度、分子质量、等电点、疏水性平均值。
1.2.2 高粱TCP基因的染色体定位 利用Mapinspect工具对高粱TCP基因进行染色体定位。
1.2.3 高粱TCP蛋白保守结构域、保守基序(Motif)分析及系统进化树构建 将TCP基因家族成员保守结构域序列在ClustalX软件上进行比对,在GeneDoc软件中进行灰度标注。利用在线网站MEME(http://meme-suite.org/tools/meme)对高粱TCP基因家族成员进行Motif分析,参数中预测数目设置为10,其余均为默认设置。将所有的高粱TCP蛋白和PlantTFDB网站下载的拟南芥TCP蛋白在MEGA 6.0软件中通过最大简约法构建进化树。
1.2.4 高粱TCP基因表达模式分析 通过qTeller数据库(http://qteller.com/)得到高粱TCP基因在不同组织中的表达量,整合数据后通过MEV 4软件制图。
2 结果与分析
2.1 高粱TCP基因家族成员的鉴定及染色体分布
经筛选鉴定,共获得27个高粱TCP基因(表1)。高粱TCP基因编码的氨基酸数目差异较大,最多可以编码627个氨基酸(SbTCP10),最少编码80个氨基酸(SbTCP5),平均编码325个氨基酸。高粱27个TCP蛋白的分子质量在8 194.28~65 896.42 ku,等电点平均值为7.9,其中有17个大于7.0,呈碱性,有10个小于7.0,呈酸性。
27个高粱TCP基因在染色体上的位置见图1。
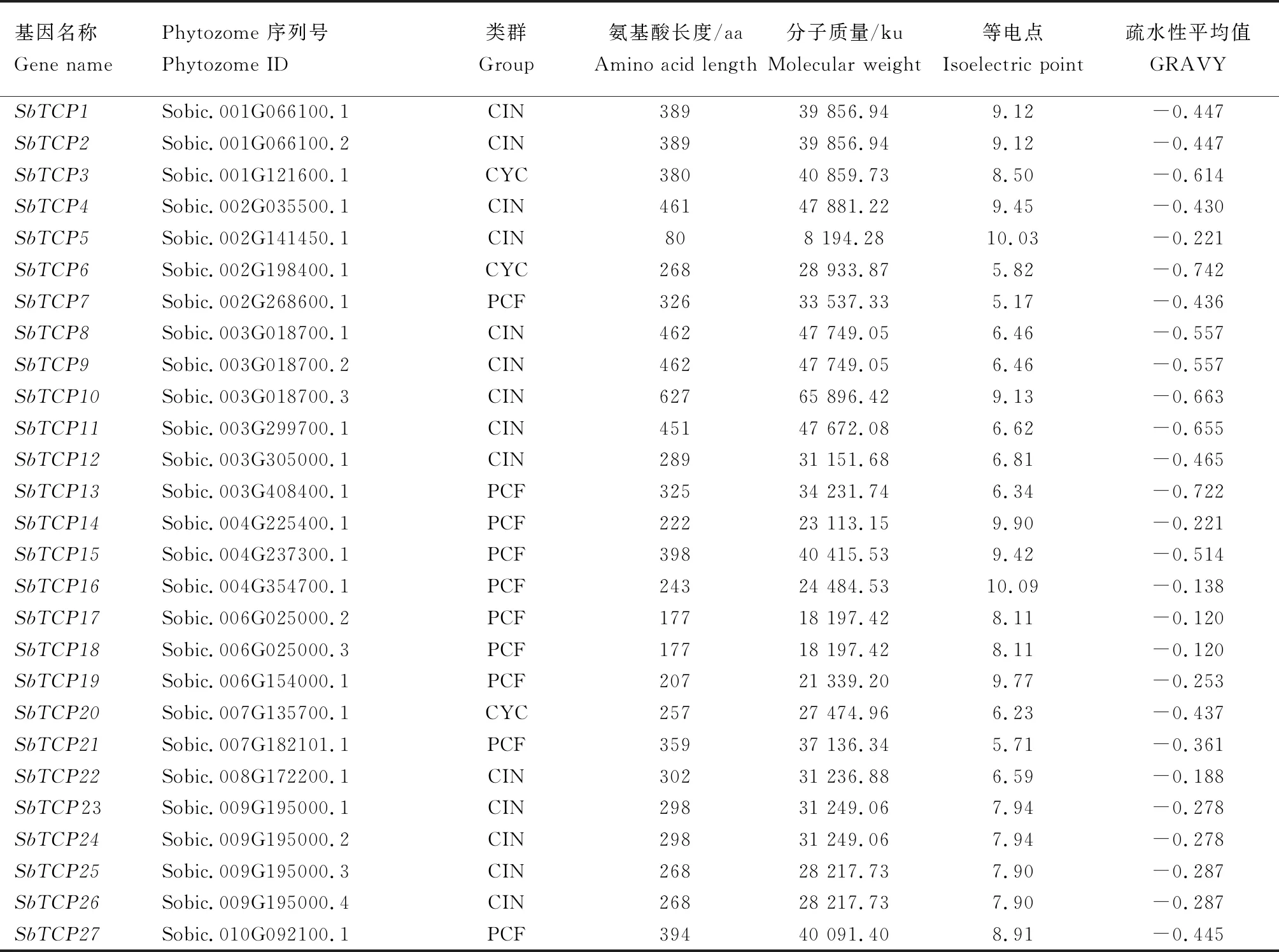
表1 高粱TCP基因家族成员基本信息
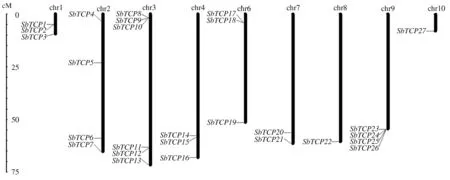
图1 高粱TCP基因染色体定位Fig.1 Chromosome localization analysis of TCP genes in sorghum
27个高粱基因在10条染色体上不均匀分布,5号染色体上没有分布。其中,1、4、6号染色体上各有3个基因,2、9号染色体上各有4个基因,3号染色体上有6个基因,7号染色体上有2个基因,8、10号染色体上各有2个基因。在1号染色体上SbTCP1和SbTCP2聚为一簇,3号染色体上的SbTCP8、SbTCP9和SbTCP10聚为一簇, 6号染色体上的SbTCP17和SbTCP18聚为一簇,9号染色体上的SbTCP23—SbTCP26基因聚为一簇。
2.2 高粱TCP蛋白的分类及系统进化分析
将27个高粱TCP蛋白与24个拟南芥TCP蛋白构建进化树(图2)。结果表明,27个高粱TCP蛋白可分为2类:ClassⅠ和ClassⅡ。ClassⅠ(PCF)包含10个高粱TCP蛋白、13个拟南芥TCP蛋白;ClassⅡ中的CYC/TB1包含3个高粱TCP蛋白、3个拟南芥TCP蛋白, CIN包含14个高粱TCP蛋白、8个拟南芥TCP蛋白。

图2 高粱和拟南芥TCP蛋白的系统进化树Fig.2 Phylogenetic tree of TCP proteins in sorghum and Arabidopsis
2.3 高粱TCP蛋白保守结构域分析
对高粱TCP蛋白的保守结构域进行比对(图3)发现,27个高粱TCP蛋白均包含有TCP结构域,即bHLH结构域,且在Basic区保守程度较高。ClassⅠ类成员在Basic区缺少第12、13、14、16位的4个氨基酸,ClassⅡ类的CIN亚组的SbTCP5蛋白缺少双螺旋区的氨基酸。在这27个TCP蛋白中发现2个存在R结构域,分别为SbTCP3、SbTCP4(图4)。
2.4 高粱TCP蛋白的Motif分析
通过MEME在线分析高粱TCP蛋白的Motif,将鉴定的10个Motif命名为Motif1—Motif10(图5)。结合高粱系统进化树进行分析发现,ClassⅠ类成员都只含有Motif1和Motif9(除SbTCP13还含有1个Motif10外);ClassⅡ类中除SbTCP5蛋白只含有1个Motif5外,其他成员都有1个Motif1和1个Motif5,且ClassⅡ类的CYC/TB1亚类中SbTCP3、SbTCP6、SbTCP20蛋白都仅含1个Motif1和1个Motif5,而ClassⅡ类的CIN亚类中SbTCP22蛋白也仅有1个Motif1和1个Motif5,其余TCP蛋白还含有其他Motif。
2.5 高粱TCP基因的表达模式和功能分析
由图6可知,高粱TCP基因在高粱各组织中的表达量不同。大部分高粱TCP基因在根、嫩枝、胚乳、胚、种子、叶片、花序原基、早期花序、雌蕊、花药、花粉中均有所表达。但ClassⅡ中的CYC/TB1亚类基因(SbTCP3/6/20)在这些组织中都不表达或表达量很低。SbTCP25/26/24/23在胚、早期花序、叶片中表达量较高;SbTCP1/2在胚、早期花序、雌蕊、种子、花序原基中表达量较高;SbTCP8/9/10在胚、雌蕊、早期花序、种子中表达量较高;SbTCP17/18在胚、嫩枝、早期花序、雌蕊、种子中表达量较高。其他TCP基因在高粱各组织中的表达量各不相同,其中在雌蕊、胚、早期花序中表达量较高,在胚乳、花粉和花药中表达量很低。总体来看,ClassⅠ大部分成员在种子、胚、雌蕊、早期花序中表达量较高,ClassⅡ中的CIN亚类在种子、叶片、胚、雌蕊、早期花序中的表达量也较高。

图3 高粱TCP蛋白保守结构域序列比对分析Fig.3 Alignment of conserved sequence of TCP proteins in sorghum

图4 高粱TCP蛋白R结构域序列分析Fig.4 Alignment of the R-domain sequence of TCP proteins in sorghum
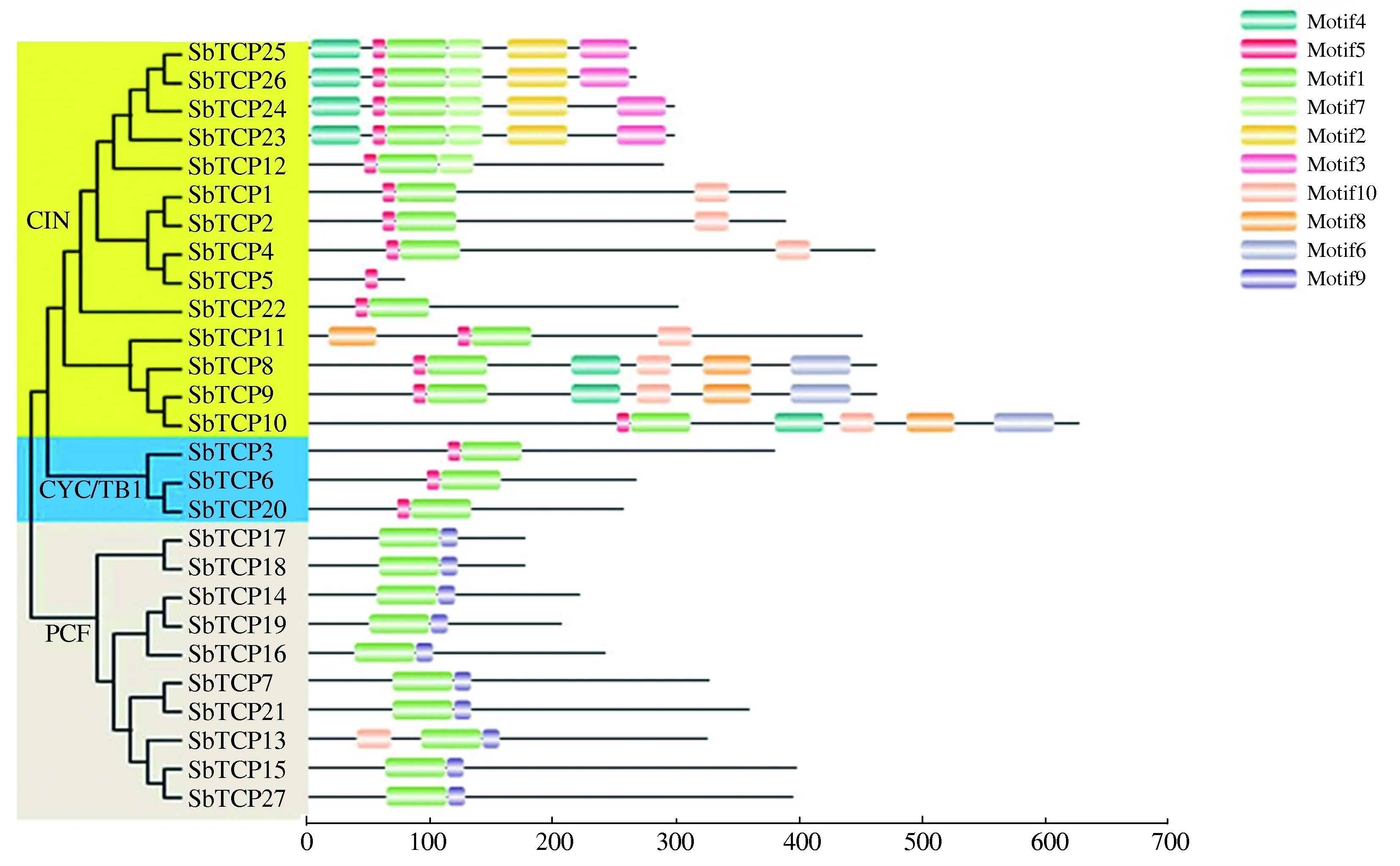
图5 高粱TCP蛋白家族成员Motif分析Fig.5 Conserved motifs analysis of TCP proteins in sorghum

图6 高粱TCP基因表达模式分析Fig.6 Expression pattern analysis of TCP genes in sorghum
从qTeller网站下载高粱根和嫩枝TCP基因在ABA(脱落酸)、NaOH、PEG(聚乙二醇)和H2O处理下的表达量,其中ABA处理与NaOH处理进行对比分析,PEG处理与水处理进行对比分析。由图6可知,与 NaOH处理相比,ABA处理后根中大部分TCP基因表达量降低,如SbTCP25/26/24/23/1/2/4/8/9/10/17/18/14/19/16/15/27等;在嫩枝中有5个TCP基因表达量明显降低,包括SbTCP25/26/24/23/12,2个TCP基因(SbTCP19/15)表达量明显升高。与水处理相比,PEG处理根中大部分TCP基因表达量明显升高,如SbTCP25/26/24/23/12/11/14/15/27等;在高粱嫩枝中,大部分TCP基因表达量下降,如SbTCP25/26/24/23/12/10等,但有个别基因表达量升高,如SbTCP4/15等。
3 结论与讨论
TCP是植物特异转录因子家族,不同物种中TCP家族基因数量不同,如大豆中有54个TCP基因[20],辣椒中有30个[21],毛竹中有19个[22],拟南芥中有24个[23]。高粱TCP成员与这些植物的TCP家族成员有着共同的特征,都含有保守的TCP结构域,且只有2个成员含有R结构域,而番茄中有6个成员具有R结构域[24],拟南芥中有5个成员具有R结构域[23]。
已有研究证实,大部分TCP转录因子在植物生长发育的过程中起重要作用,例如配子体发育、激素信号转导[17]、线粒体生物合成、植物生物节律调节、侧枝发育[13]、花发育[9]、种子萌发[8]以及叶片发育[9]等。高粱的大部分TCP基因在种子、胚、雌蕊、早期花序中表达量较高,在胚乳、花粉和花药中表达量较低,仅ClassⅠ(PCF)类成员的SbTCP21在花药中表达量较高, ClassⅠ大部分成员在种子、胚、雌蕊、早期花序中表达量较高。研究表明,拟南芥的ClassⅠ类基因AtTCP11/14/16在种子萌发和配子体发育过程中起重要的调控作用[26],暗示高粱ClassⅠ成员可能也有类似作用。而ClassⅡ中的CYC/TB1亚类基因(SbTCP3/6/20)在各组织中不表达或表达量很低,CIN亚类的SbTCP5基因也不表达或表达量很低。CIN亚类大部分成员在叶片、雌蕊、胚、种子、早期花序中表达量较高,拟南芥CIN亚类的AtTCP2/3/4/5/10/13/17/24基因参与拟南芥叶片、花发育过程[8],暗示高粱CIN成员可能也有类似作用。
TCP家族是一类与系统发育相关的植物特异转录因子,它可以在激素信号的上下游发挥作用或与激素信号互作来促进/抑制激素合成,调控相关基因表达,最终对植物的生长发育起调控作用[26]。在大豆中研究发现,GmCYL(CYC/TB1-like)基因属于TCP转录因子家族ClassⅡ类的CYC/TB1亚类成员,在叶和根中,分别不同程度地受ABA、油菜素内酯(BR)、氨基-环丙烷羧酸(ACC)、水杨酸(SA)和茉莉酸甲酯(MeJA)等诱导表达[27]。拟南芥AtTCP18基因受外源生长素的调控,从而参与植物分枝的形成过程[23]。ABA是一种重要的外界胁迫信号激素,参与维持和促进种子和芽的休眠[26]。在小麦的所有TCP基因中TaTCP5基因的表达量在热和干旱胁迫下明显上调[28]。本研究发现,ABA诱导下,根和嫩枝中的大部分基因的表达量下降;PEG处理下,根中大部分基因的表达量升高,嫩枝中大部分基因的表达量下降。在ABA和PEG胁迫下,上调表达的TCP基因可能在植物干旱胁迫响应中起重要作用。但是这些基因具体的功能和作用机制还要通过试验进一步验证。
猜你喜欢
古今农业(2022年2期)2022-08-15
植物研究(2022年4期)2022-08-04
青年文学家(2022年1期)2022-03-11
今日农业(2021年20期)2021-11-26
园艺与种苗(2020年12期)2021-01-08
果农之友(2018年5期)2018-10-19
科学种养(2017年6期)2017-06-13
科学种养(2017年5期)2017-05-17
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
