
延迟首次投喂对乌江中华倒刺鲃仔鱼补偿生长的影响
2019-10-16 10:02:30蒋天勇李忠利蔡红艳胡兰黄秀中吴江毅
水产养殖 2019年10期
蒋天勇,李忠利,蔡红艳,胡兰,黄秀中,吴江毅
(铜仁学院农林工程与规划学院,贵州 铜仁 554300)
鱼类的生长是对摄取的外界饵料进行同化合成和异化分解,通过进一步的利用从而形成对物质和能量的积累过程,且同化和异化是紧密联系的动态平衡关系。在自然或人工养殖条件下,早期鱼类经常遭遇温度、光照、盐度及营养等环境因素的胁迫而偏离正常生长,其中饵料缺乏、营养不足甚至饥饿是早期鱼类普遍遇到的营养胁迫因子[1-2]。在营养胁迫下,早期鱼类代谢水平降低,生长减缓甚至停滞,当饵料补充后鱼体又会表现出不同程度的补偿生长特征[3-4],主要分为超速补偿、完全补偿、部分补偿和不能补偿四种类型[5]。李芹等[6]通过饥饿和再投喂试验,发现饥饿15 d内的厚颌鲂(Megalobrama pellegrini)稚鱼在恢复投饵后表现出不同程度的补偿生长现象,且随着饥饿时间的延长,补偿生长能力越弱。Li等[7]研究发现16日龄的岩原鲤(Procypris rabaudi)仔鱼饥饿 4~5 d 后再投饵 10 d,体质量与正常组存在统计学意义差异,仅能完成部分补偿生长。
有关延迟首次投喂在鲤形目鱼类中(Cyprini-formes)的报道,王川等[8]研究发现饥饿4 d及以上再投喂的胭脂鱼(Myxocyprinus asiaticus)仔鱼显示出部分补偿生长特点。在印度2种鲤科鱼类(Cyprinidae)的研究中发现,越接近PNR点投喂的仔鱼,其体质量增长率越低,死亡率越高[9-10]。而有关中华倒刺鲃(Spinibarbus sinensis)仔鱼的延迟首次投喂研究,高元彧[11]侧重分析了消化酶活力方面的内容,其他有关该鱼的饥饿再投喂试验主要在幼鱼上[12]。该研究探讨了延迟首次投喂条件下,中华倒刺鲃仔鱼的全长、体质量的恢复生长特征,分析仔鱼对饵料缺乏的生态适应对策,为研究自然环境下仔鱼的胁迫适应和人工养殖过程中苗种培育的生产管理提供参考资料。
1 材料与方法
1.1 试验材料的获取
2018年6月从乌江彭水人工增殖放流站采集中华倒刺鲃初孵仔鱼,运回实验室后暂养于3个白色塑料水簇箱中(50 cm×35 cm×30 cm),至 8日龄肠道贯通(以肛门处胎粪为依据)开始试验。试验期间水温(26.5±1.2)℃,溶氧(5.4±0.3)mg/L,试验用水为曝气24 h的自来水,每天早上换水1次,换水量为总量的1/3。
1.2 试验分组与设计
选择体表正常的仔鱼移入1 L玻璃水簇箱中,每箱放鱼60尾。试验共设计0 d(即正常组H0)、1 d(饥饿1 d H1,下同)、3、5、7、9和11 d共 7个不同处理组(每组3个重复),在肠道贯通后饥饿不同天数下进行首次投喂,饵料为经24 h孵化的卤虫无节幼体(Artemia sinica,山东天成生物科技有限公司生产,粗蛋白约60%,粗脂肪约20%,粗灰分约10%),早晚各1次,以箱中无剩饵为限,记录实际投喂量。各饥饿组恢复投喂时间17 d。
1.3 指标的测定与计算
全长、体质量测定:各组每间隔4 d测量1次,每个水簇箱随机抽取10尾仔鱼,用游标卡尺测量全长(0.1 mm),电子天平称量体质量(0.1 mg,10尾1次,每箱取样3次),
初次摄食率:首次投喂饵料时摄食的个体尾数占测试尾数的百分比。
PNR点:参考殷名称[13]方法统计,即摄食尾数少于50%时的天数。
生长率:参考殷名称[13]方法,特定生长率gs=(ln M2-ln M1)/(t2-t1),其中M2和M1分别为时间t2、t1时的全长或体质量。
1.4 数据的处理与分析
用Microsoft Excel 2010软件将数据初步整理后用Spss Satistics 17.0进行统计检验并作图,全长、体质量的比较用单因素方差分析(One-Way ANOVA)-DUNCAN法进行分析,试验结果以平均值±标准差(Mean±SD)表示。
2 结果
2.1 初次摄食率、PNR点和病死率
8日龄仔鱼肠道贯通,此时肛门处出现一团胎粪,腹部还残留极少量卵黄。初次摄食率及即时病死率见表1。H0、H1和H3初次摄食率为100%,从H5开始,初次摄食率明显下降,至H11降至33.3%,此时即为PNR点。初次摄食率随饥饿时间的延长呈减小趋势。正常组无死亡,H1、H3、H5和H7有少量仔鱼死亡,H9和H11出现部分死亡现象,即时病死率随饥饿时间的延长呈增加趋势。从H5开始仔鱼鱼体明显消瘦、胆囊膨大,H9和H11胸角明显。

表1 各组饥饿后的初次摄食率和病死率
2.2 全长的补偿生长
各组全长值的比较见表2。各饥饿组饥饿后经17 d投喂,组间终末全长除了H1、H3和H5,H7和H9之间差异不具统计学意义外(P>0.05),其余各组都存在统计学意义差异(P<0.05)。各饥饿组间全长增长值比较,除 H1、H3 和 H5,H7 和 H9,H9 和 H11之间差异不具统计学意义外(P>0.05),其余各组间存在统计学意义差异(P<0.05);和对应时间段的H0相比较,除H1、H3和H5差异不具统计学意义外(P>0.05),其余各组全长增长值都存在统计学意义差异(P<0.05)。
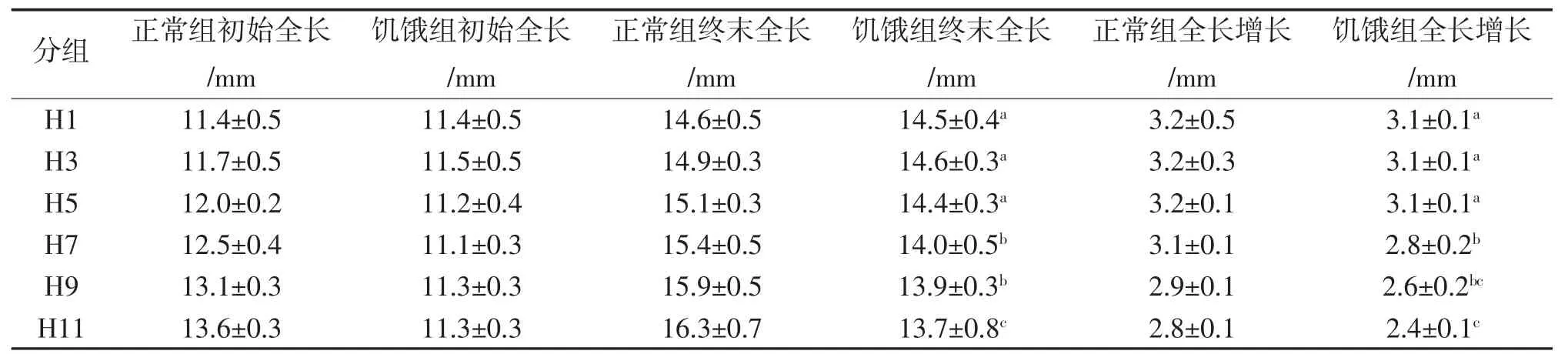
表2 各组的全长值的比较
2.3 体质量的补偿生长
各组体质量值的比较见表3。各饥饿组饥饿后经17 d投喂,组间终末体质量除了H1、H3和H5,H7和H9之间差异不具统计学意义外(P>0.05),其余组间终末体质量存在统计学意义差异(P<0.05);各组间体质量增长值比较,除H1、H3和H5,H7和H9之间差异不具统计学意义外(P>0.05),其余各组间存在统计学意义差异(P<0.05);和对应时间段的H0相比较,除H9和H11差异具统计学意义外(P<0.05),其余各饥饿组体质量增长值差异都不具统计学意义(P>0.05)。

表3 各组的体质量值的比较
2.4 特定生长率的变化
全长特定生长率的变化见表4,除H9和H11在第5天趋近于0且差异具统计学意义外(P<0.05),其余各组间特定生长率的差异都不具统计学意义(P>0.05)。而与正常组(测定值范围0.007-0.027)相比较,除H9和H11存在统计学意义性差异外(P<0.05),其余各组在第 5、9、13 和 17天与对应H0之间差异都不具统计学意义(P>0.05)。各组内全长的特定生长率都无明显的递增或递减变化趋势。

表4 不同天数下各组全长的特定生长率
不同天数下各组体质量的特定生长率见表5。各组间特定生长率在第5、9、13和17天都存在不同程度的统计学意义性差异(P<0.05)。而与H0(测定值范围 0.022~0.128)相比较,各组在第 5、9、13 和17天与对应H0之间都存在统计学意义性差异(P<0.05)。各组内体质量的特定生长率都无明显的递增或递减变化趋势。
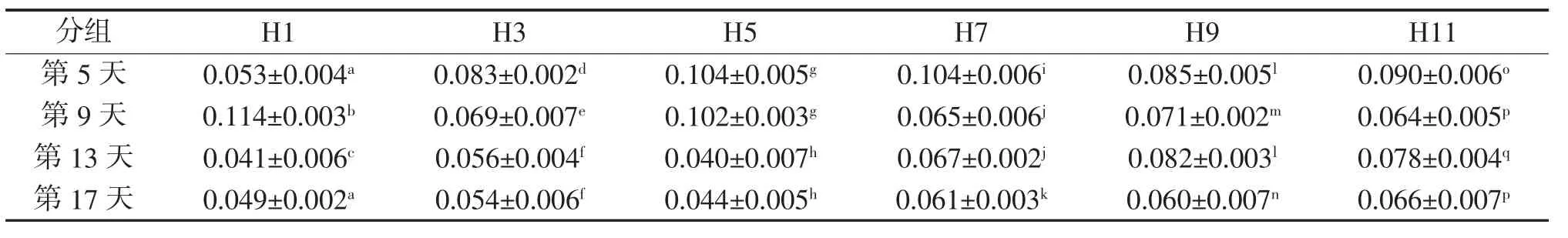
表5 不同天数下各组体质量的特定生长率
3 讨论
3.1 延迟首次投喂对仔鱼存活的影响
仔鱼开口上浮后需要即时进行摄食,以保证自身的营养需求。但在野生和人工养殖过程中,仔鱼经常遭遇饵料缺乏甚至饥饿。随着饥饿时间的延长,当饥饿的累积效应变得不可逆转时,饥饿仔鱼将达到PNR点,此时即使有食物供应,多数仔鱼也会因无法正常游泳和摄食而死亡[14-15]。Penaz[15]在对两种鲤科鱼类(Chondrostoma nasus和Barbusbarbus)的研究中认为这种死亡是仔鱼生活的外在生态条件和内在生理因素共同作用的结果。Malhotra和Munshi[9]在对异鲴(Aspidoparia morar)仔鱼的研究中认为,随着首次投喂时间的推迟,仔鱼病死率逐渐上升,该研究也观察到相似结果,且仔鱼的初次摄食率也会随首次投喂时间的延迟而逐渐降低,同时饵料日转化率的结果显示,饥饿11 d内的仔鱼在恢复投喂后出现一定的上升,但投饵17 d内都无法达到正常投喂水平。
3.2 延迟首次投喂下仔鱼的补偿生长
谢小军等[5]根据饥饿再投饵条件下鱼体体质量与正常投饵体质量的关系,将补偿生长分为四种类型。不同的鲤形目鱼类在延迟首次投喂下通常表现出不同的结果,在岩原鲤和胭脂鱼仔鱼的研究中发现,饥饿3 d及以内的仔鱼在恢复投喂后,其体质量均能显示出完全补偿生长,在之后的时间(岩原鲤2 d、胭脂鱼8 d)表现为部分补偿生长[7-8]。Mookerji和Rao[10]在南亚野鲮(Labeo rohita)仔鱼的研究中也发现,延迟首次投喂5 d后仔鱼体质量的增加量和存活率才出现明显下降。而该研究结果显示中华倒刺鲃仔鱼的显著变化时间稍晚,在延迟首次投喂7 d后的仔鱼在全长和体质量的增长值上才开始表现出统计学意义性差异,即在饥饿5 d内恢复投饵能表现出完全补偿生长,7~11 d仅能完成部分补偿生长。而厚颌鲂稚鱼的补偿生长效果更明显,李芹等[6]以30 d为试验天数,发现饥饿3 d内恢复投喂能表现出超补偿生长,这可能与不同阶段鱼体的发育状况、食性及饵料质量有关[16]。
胭脂鱼仔鱼全长和体质量的特定生长率随饥饿天数的延长而减小[8],而该研究中中华倒刺鲃仔鱼无明显变化趋势,这可能与仔鱼种类有关。
3.3 仔鱼的开口投喂时间
掌握最佳的首次投喂时间能保证仔鱼获得较高的成活率,并满足正常的生长发育以及后期的繁衍。卵黄囊期结束时,仔鱼可以短期忍受饥饿,但必须即时获取足够的饵料来满足生存和生长,延迟首次投喂试验为仔鱼的早期培育提供了理论参考,这在淡水和海水鱼类中都有报道[17-19]。在已报道的鲤形目鱼类中,以常规的全长、体质量、生长率、摄食率等生物学指标作为衡量标准,短期饥饿下仔鱼能完成完全补偿生长,随着饥饿时间的延长其补偿能力逐渐减弱。Malhotra和Munshi[9]指出不管是天然饵料还是人工饲料,异鲴仔鱼首次外源投喂的最佳时间都应在肠道发育完全到蛋黄吸收完全之间的时间。该研究中,尽管全长和体质量在饥饿5 d内投饵能表现出完全补偿生长,但结合初次摄食率和病死率来看,建议首次投喂的最佳时间应在肠道贯通时。
猜你喜欢
渔业研究(2022年5期)2022-10-28 01:56:14
当代水产(2021年1期)2021-03-19 05:17:14
防护林科技(2020年9期)2020-11-09 01:45:24
当代水产(2019年9期)2019-10-08 08:02:44
甘肃畜牧兽医(2019年2期)2019-03-20 01:49:42
中南林业调查规划(2018年3期)2018-11-24 03:18:48
广东海洋大学学报(2015年4期)2016-01-13 08:39:40
中南林业调查规划(2015年3期)2015-12-20 05:22:31
集美大学学报(自然科学版)(2015年4期)2015-02-28 01:13:34
应用海洋学学报(2014年1期)2014-11-22 07:17:50
