
棉铃虫中肠特异性结合蛋白ABCC1对Cry1Ac毒力的影响
2019-10-12 01:57:30陈琳魏纪珍刘臣牛琳琳张彩虹梁革梅
中国农业科学 2019年19期
陈琳,魏纪珍,刘臣,牛琳琳,张彩虹,梁革梅
棉铃虫中肠特异性结合蛋白ABCC1对Cry1Ac毒力的影响
陈琳1,魏纪珍2,刘臣1,牛琳琳1,张彩虹1,梁革梅1
(1中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室,北京 100193;2河南农业大学植物保护学院,郑州 450002)
【】研究棉铃虫()中肠蛋白ABCC1(HaABCC1)与Cry1Ac的结合特性及对Cry1Ac毒力的影响,明确HaABCC1在Cry1Ac杀虫机制中的作用。分析基因序列,设计引物,通过原核表达得到HaABCC1两个跨膜区片段的蛋白,与Cry1Ac进行Ligand blot试验,验证其与Cry1Ac的体外结合特性;利用RNAi技术干扰棉铃虫幼虫的,在3龄幼虫腹部注射siABCC1,比较的表达量及Cry1Ac处理后棉铃虫死亡率的变化;通过细胞转染将ABCC1导入Sf9细胞系中,确定pAc-ABCC1重组质粒转入Sf9细胞后,用细胞生物测定的方法比较Cry1Ac处理后细胞死亡率的变化;比较敏感品系(96S)和Cry1Ac抗性品系(BtR)棉铃虫的基因全长序列,并通过荧光定量RT-PCR检测在抗、感棉铃虫中的表达量。HaABCC1跨膜区TMD1和TMD2在BL21(DE3)感受态细胞中成功表达,两个HaABCC1跨膜区片段蛋白均能与活化的Cry1Ac在体外结合;棉铃虫注射siABCC1后,的表达量显著下降,与未注射的棉铃虫、注射DEPC水和siEGFP的棉铃虫相比,用活化的Cry1Ac蛋白处理被干扰的棉铃虫,其幼虫死亡率显著降低,表明棉铃虫幼虫的被干扰后,能显著降低Cry1Ac对棉铃虫的毒力;用活化的Cry1Ac蛋白处理成功转入的Sf9细胞,与对照Sf9细胞相比,细胞的死亡率明显上升,表明将导入Sf9后能显著提高Cry1Ac处理后的细胞死亡率;抗性品系(BtR)与敏感品系(96S)棉铃虫的氨基酸序列没有差别,但抗性品系BtR棉铃虫的表达量显著降低。HaABCC1是Cry1Ac的特异性结合蛋白,可能是Cry1Ac的功能受体蛋白,并可能参与对Cry1Ac的抗性机制。
棉铃虫;ABCC1;苏云金芽孢杆菌;Cry1Ac;结合蛋白;毒力
0 引言
【研究意义】苏云金芽孢杆菌(,Bt)产生的晶体蛋白对鳞翅目、双翅目、鞘翅目等多种昆虫和线虫具有特异性毒性,但对人畜无害[1-4]。Bt能够产生具有杀虫活性的伴孢晶体,Cry蛋白是伴孢晶体中的重要一类,目前已发现约有300多种可以分为40个不同的小类Cry蛋白,Cry1Ac蛋白是其中最早被应用于害虫防治的[1-4]。表达Bt蛋白的抗虫转基因农作物已在美国、巴西、印度、中国等多个国家广泛种植,这不仅可以有效降低相关农业害虫的危害,还可以减少由于化学农药过度使用带来的环境污染[4-5]。但是随着转Bt基因农作物种植面积的扩大,害虫对Bt蛋白的抗性[3,6-8]问题也日益严重。因此,研究昆虫对Bt蛋白产生抗性的机理对于延迟抗性产生至关重要。【前人研究进展】当昆虫取食Bt蛋白后,Bt蛋白会在昆虫中肠碱性环境中被溶解、活化,并与中肠上皮细胞上的特异性受体结合,随后造成中肠膜穿孔,细胞溶解,最终导致昆虫死亡[3]。Bt蛋白与昆虫中肠受体结合是其发挥杀虫作用的关键,主要受体蛋白表达量的变化或基因突变是引起昆虫对Bt蛋白产生抗性的重要原因。目前报道的主要受体蛋白有钙黏蛋白(cadherin,CAD)、氨肽酶N(aminopeptidase N,APN)、碱性磷酸酯酶(alkaline phosphatase,ALP)、腺苷三磷酸结合转运蛋白(ATP-binding cassette transporter,ABC)等[9-15]。ABC转运蛋白家族蛋白包含4个核心结构域:两个跨膜结构域(TMD),每个结构域由6个跨膜螺旋组成;两个位于胞质侧的核苷酸结合区(NBD),与两个跨膜结构域交替。ABC转运蛋白的功能与细胞物质转运和相关物质的代谢有关[16-18],已有报道表明,ABC转运蛋白不仅是Bt的受体,而且ABC基因的突变或表达量的改变与昆虫对Bt产生抗性相关[14-16]。【本研究切入点】前期研究表明,对Bt蛋白敏感棉铃虫()的发生突变,造成翻译的蛋白序列提前终止,HaABCC2蛋白功能丧失,会导致棉铃虫对Cry1Ac蛋白抗性的产生[14]。ABC转运蛋白是一个庞大的家族,且在细胞膜上以多种ABC的形式混合存在,因此研究其他ABC在Cry1Ac杀虫机制及棉铃虫对Cry1Ac抗性机制中的作用很有必要。【拟解决的关键问题】明确棉铃虫HaABCC1是否为Cry1Ac的功能性受体蛋白,表达量的变化是否与棉铃虫对Cry1Ac的抗性有关,进一步揭示Cry1Ac的杀虫机制及棉铃虫产生抗性的分子机制,为合理应用Bt及转基因抗虫作物、制定合理的抗性治理策略提供理论依据。
1 材料与方法
试验于2017—2018年在中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室完成。
1.1 供试昆虫
棉铃虫新乡敏感品系(96S):1996年6月采自河南省新乡县棉田,在实验室内用人工饲料[19]饲养至今,未接触过任何杀虫剂。敏感棉铃虫品系在昆虫饲养间以温度(27±2)℃,光周期14L﹕10D,相对湿度(75±10)%的条件下饲养。成虫在产卵笼中饲养并用10%蜂蜜水补充营养。
棉铃虫Cry1Ac抗性品系(BtR):室内用3.6 mg·mL-1Cry1Ac原蛋白对96S敏感品系筛选了210代,抗性倍数为3 600倍[20]。
1.2 Cry1Ac蛋白
活化的Cry1Ac蛋白购于北京乐士宁科技有限公司(Envirologix)。生物素标记试剂盒购于武汉伊莱瑞特生物科技股份有限公司(Elabscience)。用生物素标记活化的Cry1Ac蛋白,标记过程参考试剂盒说明书。
1.3 取样方法
收集敏感品系(96S)与抗性品系(BtR)棉铃虫5龄幼虫各10头,在冰上解剖,去除食物截取中肠,用4℃预冷的0.7% NaCl溶液将内含物冲洗干净,再用滤纸吸干水分,-80℃保存备用。
1.4 总RNA的提取与cDNA的合成
按照Invitrogen操作说明分别提取各样品总RNA,RT-PCR和荧光定量RT-PCR所用的cDNA模板的合成参照Tiangen公司Fast QuantRT kit说明书和SuperRealPreMix(Probe)说明书。所有样品设3次生物学重复。
1.5 原核表达及Ligand blot分析
利用Primer 5.0软件分别设计两个跨膜区(TMD)携带酶切位点RV和dⅢ的引物(表1),名称分别为TM1-RV-F、TM1-dIII-R;TM2-RV-F、TM2-dIII-R。把PCR合成的目的基因转入克隆载体,取测序正确的质粒和表达载体pET32a进行双酶切,用T4-DNA连接酶将目的片段和表达载体构建为重组质粒。将测序正确的重组质粒转入BL21(DE3)感受态细胞中并转接500 ml LB培养基中培养,加入0.8 mmol·L-1IPTG并在25℃过夜诱导表达。高速低温离心收集菌体并超声破碎,经SDS-PAGE电泳并切取目的片段送至北京华大蛋白质研发中心进行质谱检测。并用购于生工生物工程有限公司的PAGE胶蛋白微量回收试剂盒回收目的蛋白。
用Ligand blot检测原核表达得到的HaABCC1两个跨膜区片段蛋白能否与活化的Cry1Ac在体外结合。将纯化蛋白经过电泳转到PVDF膜上;加入30 ml封闭液到自封袋中并放入PVDF膜,置于摇床上80 r/min室温孵育2 h;重新加入新的30 ml脱脂牛奶再加入5 μl生物素标记的活化Cry1Ac,置于摇床上80 r/min孵育1 h;重新加入20 ml脱脂牛奶再加入2 μl二抗(HRP标记的山羊抗小鼠抗体)到自封袋中,室温孵育1 h,PBST洗膜3次,每次10 min;用ECL方法进行曝光,所用的试剂盒为EASYsee底物发光液。
1.6 RNAi及生物测定
利用RNAi干扰敏感品系(96S)棉铃虫的。为了提高RNAi干扰的特异性,从高特异性的跨膜区片段设计siRNA序列。并用加强荧光蛋白基因()作为对照。siRNA由生工生物工程有限公司合成(表1),名称分别为siRNA-ABCC1和siRNA-EGFP。
用5 μl微量注射器往3龄初棉铃虫幼虫的腹部注射2 μg·μl-1的siABCC1 1 μL。注射点用凡士林封闭,防止体液过多流失。用不被注射的棉铃虫以及注射siEGFP和DEPC水的棉铃虫作为对照。每个处理24头棉铃虫,进行3次重复。干扰后48 h利用荧光定量RT-PCR检测不同处理棉铃虫的表达量。
采用生物测定的方法测定Cry1Ac对干扰棉铃虫后死亡率的变化。往24孔板的每个孔中加入1 ml刚制作完成的棉铃虫人工饲料(液态)。待其凝固后往每个孔中加入75 μl活化的Cry1Ac蛋白使每个孔中蛋白浓度为120 μg·cm-2。接入干扰处理48 h后的棉铃虫,每个处理24头,3次重复。以加入等量的PBS溶液为对照处理。5 d后,统计各处理的死亡虫数,计算相应的死亡率[21]。
1.7 细胞转染及细胞生物测定
利用Primer 5.0软件设计带酶切位点I和I的全长引物ABCC1-I-F和ABCC1-I- R(表1)。经PCR测序鉴定正确的质粒通过双酶切和T4-DNA连接酶连接到pAc5.1b质粒上,构建重组质粒pAc-ABCC1。质粒购于北京华越洋生物有限公司。
将Sf9细胞分别平铺到12孔板中,每孔约9×105个细胞,放到28℃培养箱让细胞充分贴壁(2 h)。在Cellfectin(Invitrogen;8 μl/孔)作用下,每个孔里转染2 μg pAc5.1b(对照,空质粒)或目的基因质粒,孵育5 h。用1.5 ml Sf-900 II SFM培养基(Sf9细胞)置换转染液,然后放至28℃培养箱培养64 h。在每次独立的转染试验中,细胞被吹起,重新平铺到96孔板的3个孔中,每孔100 µL含细胞10 000个。充分贴壁2 h后,用活化的Cry1Ac处理转染细胞,活化的Cry1Ac浓度为18.5 µg·ml-1。每个质粒被重复3次转染到细胞中,每次独立的转染用活化的Cry1Ac进行3次重复的生物测定。对细胞生物测定后,Sf9细胞提取RNA并反转录为cDNA,用特异性引物ABCC1-F’和ABCC1-R’(表1)进行PCR验证,检验pAc-ABCC1重组质粒是否成功转入Sf9细胞中。
1.8 氨基酸序列对比分析
根据Genbank中的cDNA序列(KY796050),用Primer 5.0软件分别设计ABCC1-F和ABCC1-R引物(表1),以抗、感棉铃虫5龄中肠的cDNA为模板,进行PCR扩增,分别扩增出序列。按下列程序启动PCR:94℃预变性5 min;94℃变性30 s,55℃退火30 s,72℃延伸5 min,共35个循环;72℃保温10 min。PCR反应所用的高保真聚合酶购于赛默飞世尔科技(中国)有限公司(Thermo Fisher)。扩增完成后用1%琼脂糖凝胶电泳检测,利用AxygenDNA回收试剂盒回收目的条带,然后把回收产物克隆到Peasy-T3载体上,转化入Trans1-T1感受态细胞中,挑取阳性单克隆送北京博迈德(Biomed)进行序列测定。PCR获得基因全长并经Blast序列比对确认为后,用DNAMAN分别对比其对应的氨基酸序列。
1.9 荧光定量RT-PCR
利用TaqMan探针技术进行荧光定量RT-PCR,根据的cDNA序列设计、合成用于荧光定量RT-PCR的特异性引物和探针,以棉铃虫(GenBank:JF417983.1)和-(GenBank:EU527017.1)作为双内参基因,合成特异性引物和探针,探针5′端采用FAM标记,3′端采用BHQ标记。引物和探针在上海Invitrogen公司合成(表1)。在20 μL荧光定量RT-PCR反应体系中含有:2× SuperReal Premix 10 μL正向引物(10 μmol·L-1)0.6 μL,反向引物(10 μmol·L-1)0.6 μL,荧光探针(10 μmol·L-1)0.4 μL,cDNA模板1 μL,50×ROX ReferenceDye 0.2 μL,RNase-Free ddH2O 7.2 μL。混匀离心后在ABI 7500荧光定量RT-PCR仪上按照如下条件反应:95℃预变性15 min,然后95℃变性3 s,60℃退火/延伸32 s,共40个循环。反应中以水作为阴性对照,每个处理3个技术重复。
1.10 数据处理
对荧光定量RT-PCR结果采用2-ΔΔCt法进行计算[22]。采用SPSS软件对处理数据进行分析,处理间比较采用单因素方差分析检测,用Turkey法进行多重比较。

表1 各RT-PCR、荧光定量RT-PCR及RNAi反应所用引物
2 结果
2.1 HaABCC1跨膜区原核表达及Ligand blot分析
通过PCR技术,经过测序验证成功克隆得到HaABCC1跨膜区TMD1(924 bp)和TMD2(882 bp),并在BL21(DE3)感受态细胞中成功原核表达,蛋白片段大小分别为34和33 kD,与预期结果相符(图1-A)。经过Ligand blot检测,HaABCC1跨膜区原核表达片段均能与活化的Cry1Ac蛋白结合(图1-B)。
2.2 HaABCC1功能验证
被干扰后,相较于未注射的棉铃虫以及注射DEPC水和siEGFP的棉铃虫,的表达量分别下降了54.0%、49.4%和52.2%(<0.05)(图2-A)。被干扰后的棉铃虫,除表达量明显降低外,幼虫对Cry1Ac的敏感性也发生显著改变。用活化的Cry1Ac蛋白处理被干扰的棉铃虫,与未注射的棉铃虫以及注射DEPC水和siEGFP的棉铃虫相比,其幼虫死亡率分别下降了65.9%、60.0%和62.2%(<0.05)(图2-B)。

A:Marker:蛋白标准品protein marker;1:超声破碎处理后TMD1片段TMD1 fragment after ultrasound treatment;2:超声破碎处理后TMD2片段TMD2 fragment after ultrasound treatment。B:1、4:蛋白标准品protein marker;3、6:BSA蛋白,作为对照BSA, as a control;2:TMD1片段与活化的Cry1Ac蛋白结合TMD1 fragment bound with Cry1Ac;5:TMD2片段与活化的Cry1Ac蛋白结合TMD2 fragment bound with Cry1Ac
与转入pAc空质粒的Sf9细胞相比,可以看到棉铃虫pAc-ABCC1被成功导入Sf9细胞中(图3-A)。用18.5 µg·ml-1的活化Cry1Ac蛋白处理成功转入的Sf9细胞,结果发现细胞的死亡率显著上升(<0.05)(图3-B),为33.8%,显著高于未转入的Sf9细胞的细胞死亡率(2.0%)。
2.3 抗、敏品系棉铃虫HaABCC1的cDNA及氨基酸序列对比分析
比较Cry1Ac抗性棉铃虫与敏感棉铃虫的全长,发现在Cry1Ac抗性品系(BtR)和敏感品系(96S)棉铃虫只有少数碱基的差异,在氨基酸水平上没有任何差异。
2.4 抗、敏品系棉铃虫HaABCC1的mRNA表达量分析
进一步比较了在抗性品系(BtR)和敏感品系(96S)棉铃虫中表达量的差异,发现在抗性品系棉铃虫中的表达量显著降低(<0.05),敏感品系的表达量是抗性品系的4.19倍(图4)。
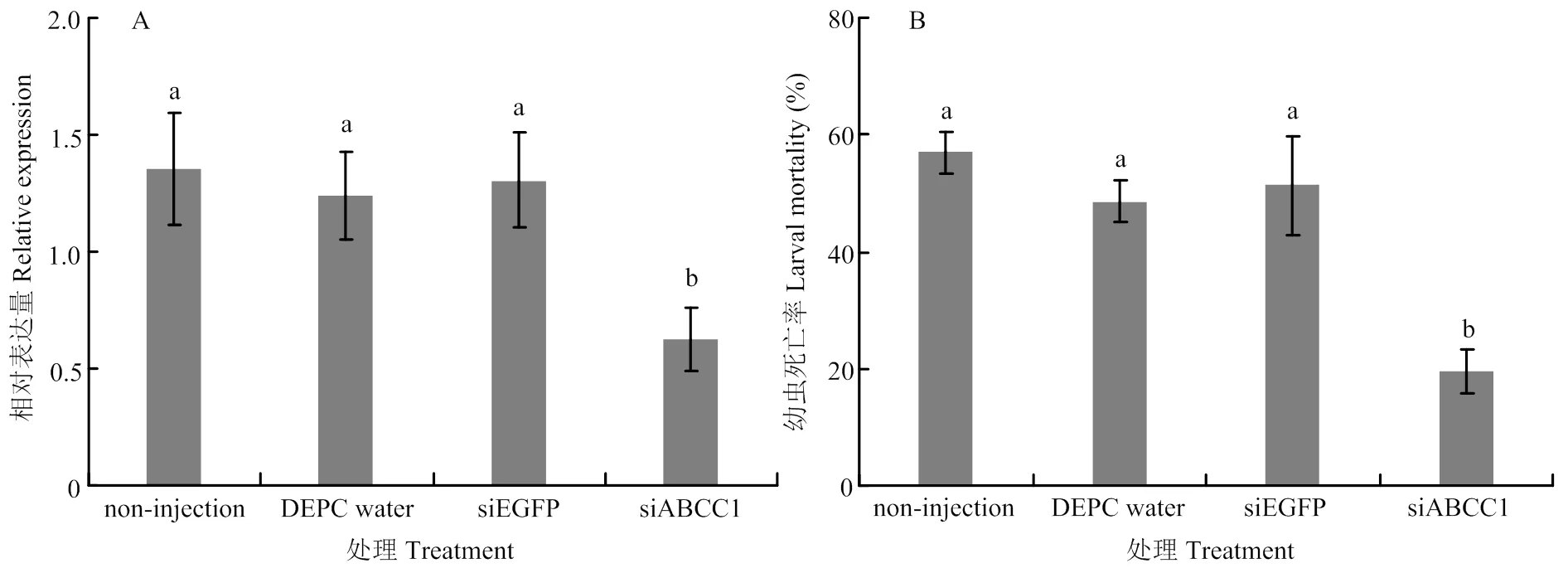
柱上不同小写字母表示处理间差异显著(P<0.05)。下同
3 讨论
昆虫中肠上皮细胞刷状缘膜囊泡BBMV上的受体与活化的Bt蛋白特异性结合在Bt杀虫机制中起着关键作用[23-24]。已经有相关报道证实ABC转运蛋白是Bt蛋白的受体。例如,Zhou等[15]通过蛋白质体外结合试验,发现Cry1Ac能与ABCC2结合;TanakA等[25]在昆虫细胞内分别表达突变型(酪氨酸插入)和野生型家蚕的,通过分析细胞对Bt的敏感性变化发现参与了Cry1Ab、Cry1Ac、Cry1Fa和Cry8Ca的杀虫过程;Chen等[26]通过在昆虫细胞系表达斜纹夜蛾(Sl-HP),证实其为Cry1Ac的功能受体。本研究选择在HaABCC1转运蛋白中特异性较高的两个跨膜结构域,通过在大肠杆菌中原核表达得到了两个HaABCC1跨膜结构域蛋白,通过Ligand blot分析确定这两个跨膜区片段蛋白均能与Cry1Ac在体外结合。被干扰后棉铃虫对Cry1Ac的敏感性显著降低,而且在Sf9细胞中表达能够提高Cry1Ac处理后的细胞死亡率。这些结果表明HaABCC1是Cry1Ac的特异性结合蛋白,可能是Cry1Ac的功能性受体蛋白。

A:PCR检测pAc-ABCC1质粒转入Sf9细胞Confirmation of HaABCC1 expression in cells transfected with pAc-ABCC1 by PCR。1—9:转入pAc-ABCC1质粒的Sf9细胞(3个生物学重复× 3个技术重复)pAc-ABCC1 plasmid transfected into the Sf9 cell line (3 biological replicates × 3 technical replicates);10—18:转入pAc空质粒的Sf9细胞(3个生物学重复× 3个技术重复)pAc empty plasmid transfected into the Sf9 cell line (3 biological replicates × 3 technical replicates)。B:用活化的Cry1Ac蛋白处理转入pAc-ABCC1质粒和转入pAc空质粒的Sf9细胞死亡率The mortality of Sf9 cells exposed to Cry1Ac after transfected with the pAc-ABCC1 or pAc empty plasmid

图4 HaABCC1在不同品系中的表达量
前期研究发现,在棉铃虫不同发育阶段均有表达,在4龄、5龄幼虫中表达量最高,在成虫中最低;在不同组织中均有表达,在马氏管中表达量最高,在前肠中最低。这可能与ABC转运蛋白家族的功能相关,如在哺乳类动物中ABC转运蛋白参与离子运输和毒素分泌的过程,能够将不同的外源物质、重金属等运输到体外,因此具有抵御抗体、化学治疗药物和农药的功能[27]。近年来,关于ABC转运蛋白的变化与昆虫对Bt的抗性关系也受到普遍关注。如鳞翅目昆虫的ABCB、ABCD、ABCE、ABCF之间存在一定的同源性且与昆虫对Bt的抗性有关[28];敏感棉铃虫的发生突变,造成其翻译的蛋白序列提前终止,HaABCC2蛋白功能丧失,这会导致棉铃虫对Cry1Ac蛋白抗性的产生[14];小菜蛾()突变会引起其对Cry1Ac蛋白抗性的产生[29];小菜蛾被干扰后,会显著提高其对Cry1Ac蛋白的抗性[30];棉铃虫的突变会引起棉铃虫对Cry2Ab蛋白的抗性[31]。本研究发现,虽然Cry1Ac抗性棉铃虫编码的氨基酸序列没有发生变化,但与敏感品系相比,抗性品系棉铃虫的表达量显著降低,推测表达量的降低可能与棉铃虫对Cry1Ac抗性的产生相关。
由于棉铃虫HaABCC1蛋白具有与其他ABC蛋白家族相似的结构,含有2个跨膜结构域和2个膜外区域,含有14个N-糖基化位点和16个O-糖基化位点。尤其是2个富含糖基化位点的跨膜结构域可能与Bt杀虫作用及昆虫对Bt的抗性相关。有关ABCC2在Bt杀虫机理中的作用,Gahan等[32]推测Cry蛋白与ABCC2结合可能是形成寡聚体的基础,并把穿孔前的结构拉近到跨膜区以便于穿孔;Tanaka等[25]研究发现,在细胞内共表达钙黏蛋白(CaLP)和ABCC2,共表达后的细胞比仅表达ABCC2时更大;类似地,在Sf9细胞中共表达HevABCC2()和HevCaLP,对细胞有更高的毒性;而Tay等推测ABCA2可以独立参与Cry2Ab的作用模型,来完成结合和穿孔[31];真核表达抗、感家蚕,结果表明该基因协助穿孔[33];的修饰和敲除可能阻碍了Bt蛋白作用机制的最后一步[24]。基于不同ABC转运蛋白之间相似的结构,棉铃虫ABCC1也可能与其他ABC转运蛋白一样,两个跨膜结构域的变化可能与昆虫对Bt的抗性相关。但本研究并没有发现HaABCC1两个跨膜结构域的氨基酸序列发生改变,全长氨基酸序列也没有变化,仅表达量发生了改变。由于ABC蛋白是一个庞大的家族,且在细胞膜上也混合存在,有可能是很多种蛋白共同起作用。因此,笔者推测ABCC1可能和其他ABC家族蛋白或与其他受体蛋白共同在Bt杀虫机制及抗性机制中起作用。
Tay等[31]利用EPIC(exon-primedinton-crossing)技术和目的基因的测序技术,通过双向遗传连锁分析了抗Cry2Ab棉铃虫,发现其抗性的产生是由于ATP结合盒转运体基因上的3个与抗性相关的INDEL的突变而引起,且和的突变与棉铃虫对Cry2Ab的抗性相关;Xiao等[14]也报道了的突变引起棉铃虫对Cry2Ab的抗性。前期研究发现被干扰后,棉铃虫对Cry2Ab的敏感性显著降低,推测ABCC1也是Cry2Ab的功能性受体蛋白[34]。上述结果表明,ABC转运蛋白可能不仅是Cry1A的受体蛋白,也是Cry2A的受体,这些共同的受体是否与Cry1A和Cry2A类蛋白间的交互抗性相关,有待进一步研究。
4 结论
原核表达的两个棉铃虫HaABCC1跨膜区片段蛋白均能与Cry1Ac在体外结合;被干扰后,能显著降低Cry1Ac对棉铃虫的毒力;将导入Sf9细胞后能显著提高Cry1Ac处理后的死亡率;与敏感品系相比,Cry1Ac抗性品系表达量显著降低。因此,HaABCC1可能是Cry1Ac的功能受体蛋白,并可能参与对Cry1Ac的抗性机制。
[1] Comas C, Lumbierres B, Pons X, Albajes R. No effects ofmaize on nontarget organisms in the field in southern Europe: a meta-analysis of 26 arthropod taxa., 2014, 23(1): 135-143.
[2] Nicolia A, Manzo A, Veronesi F, Rosellini D. An overview of the last 10 years of genetically engineered crop safety research., 2014, 34(1): 77-88.
[3] Pardo-López L, Soberón M, Bravo A.insecticidal three-domain Cry toxins: mode of action, insect resistance and consequences for crop protection., 2013, 37(1): 3-22.
[4] Sanahuja G, Banakar R, Twyman R M, Capell T, Christou P.: a century of research, development and commercial applications., 2011, 9(3): 283-300.
[5] ISAAA. Brief 52: Global status of commercialized biotech/GM crops//Uthaca, NY, 2016.
[6] Jakka S R K, Gong L, Hasler J, Banerjee R, Sheets J J, Narva K, Blanco C A, Jurat-Fuentes J L. Field-evolved mode 1 resistance of the fall armyworm to transgenic cry1fa- expressing corn associated with reduced cry1fa toxin binding and midgut alkaline phosphatase expression., 2016, 82(4): 1023-1034.
[7] Jin L, Zhang H, Lu Y, Yang Y, Wu K, Tabashnik B E, Wu Y. Large-scale test of the natural refuge strategy for delaying insect resistance to transgenic Bt crops., 2015, 33(2): 169-174.
[8] Tabashnik B E, Carriere Y. Surge in insect resistance to transgenic crops and prospects for sustainability., 2017, 35(10): 926-935.
[9] Caccia S, Moar W J, Chandrashekhar J, Oppert C, Anilkumar K J, Jurat-Fuentes J L, Ferre J. Association of Cry1Ac toxin resistance in(Boddie) with increased alkaline phosphatase levels in the midgut lumen., 2012, 78(16): 5690-5698.
[10] Ocelotl J, Sanchez J, Gomez I, Tabashnik B E, Bravo A, Soberon M. ABCC2 is associated withCry1Ac toxin oligomerization and membrane insertion in diamondback moth., 2017, 7(1): 2386.
[11] Tanaka S, Miyamoto K, Noda H, Endo H, Kikuta S, Sato R. Single amino acid insertions in extracellular loop 2 ofABCC2 disrupt its receptor function forCry1Ab and Cry1Ac but not Cry1Aa toxins., 2016, 78: 99-108.
[12] Wang J, Zhang H, Wang H, Zhao S, Zuo Y, Yang Y, Wu Y. Functional validation of cadherin as a receptor of Bt toxin Cry1Ac inutilizing the CRISPR/Cas9 system., 2016, 76: 11-17.
[13] Wang L Y, Gu S H, Nangong Z Y, Song P, Wang Q Y. Aminopeptidase N5 (APN5) as a putative functional receptor of Cry1Ac toxin in the larvae of., 2017, 74(4): 455-459.
[14] Xiao Y, Zhang T, Liu C, Heckel D G, Li X, Tabashnik B E, Wu K. Mis-splicing of the ABCC2 gene linked with Bt toxin resistance in., 2014, 4: 6184.
[15] Zhou Z, Wang Z, Liu Y, Liang G, Shu C, Song F, Zhou X, Bravo A, Soberon M, Zhang J. Identification of ABCC2 as a binding protein of Cry1Ac on brush border membrane vesicles fromby an improved pull-down assay., 2016, 5(4): 659-669.
[16] Dassa E, Bouige P. The ABC of ABCS: a phylogenetic and functional classification of ABC systems in living organisms., 2001, 152(3/4): 211-229.
[17] Linton K J. Structure and function of ABC transporters., 2007, 22: 122-130.
[18] 戚伟平, 马小丽, 何玮毅, 陈玮, 董人富, 尤民生. 节肢动物ABC转运蛋白及其介导的杀虫剂抗性. 昆虫学报, 2014, 57(6): 729-736.
Qi W P, Ma X L, He W Y, Chen W, Dong R F, You M S. ATP-binding cassette transporters and their mediated resistance to insecticides in arthropods., 2014, 57(6): 729-736. (in Chinese)
[19] 梁革梅, 谭维嘉, 郭予元. 人工饲养棉铃虫技术的改进. 植物保护, 1999, 25(2): 15-17.
Liang G M, Tan W J, Guo Y Y. An improvement in the technique of artificial rearing cotton bollworm., 1999, 25(2): 15-17. (in Chinese)
[20] Xiao Y, Dai Q, Hu R, Pacheco S, Yang Y, Liang G, Soberon M, Bravo A, Liu K, Wu K. A single point mutation resulting in cadherin mislocalization underpins resistance againsttoxin in cotton bollworm., 2017, 292(7): 2933-2943.
[21] 周慧丹, 杨亦桦, 吴益东. RNAi介导的棉铃虫氨肽酶N基因和钙粘蛋白基因沉默对Cry1Ac毒力的影响. 昆虫学报, 2010, 53(10): 1097-1103.
Zhou H D, Yang Y H, Wu Y D. Effects of RNAi-mediated silencing of an aminopeptidase N geneand a cadherin geneon Cry1Ac toxicity against(Lepidoptera: Noctuidae)., 2010, 53(10): 1097-1103. (in Chinese)
[22] Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta C(T)) method., 2001, 25(4): 402-408.
[23] Bravo A, Likitvivatanavong S, Gill S S, Soberon M.: A story of a successful bioinsecticide., 2011, 41(7): 423-431.
[24] Heckel D G. Learning the ABCs of Bt: ABC transporters and insect resistance toprovide clues to a crucial step in toxin mode of action., 2012, 104(2): 103-110.
[25] TanakA S, Miyamoto K, Noda H, Jurat-Fuentes J L, Yoshizawa Y, Endo H, Sato R. The ATP-binding cassette transporter subfamily c member 2 inlarvae is a functional receptor for Cry toxins from., 2013, 280(8): 1782-1794.
[26] Chen Z, He F, Xiao Y, Liu C, Li J, Yang Y, Ai H, Peng J, Hong H, Liu K. Endogenous expression of a Bt toxin receptor in the Cry1Ac-susceptible insect cell line and its synergistic effect with cadherin on cytotoxicity of activated Cry1Ac., 2015, 59: 1-17.
[27] Dean M, Hamon Y, Chimini G. The human ATP binding cassette (ABC) transporter superfamily., 2001, 42: 1007-1017.
[28] Dermauw W, Van Leeuwen T. The ABC gene family in arthropods: Comparative genomics and role in insecticide transport and resistance., 2014, 45: 89-110.
[29] Guo Z, Kang S, Zhu X, Xia J, Wu Q, Wang S, Xie W, Zhang Y. Down-regulation of a novel ABC transporter gene () is associated with Cry1Ac resistance in the diamondback moth,(L.)., 2015, 59: 30-40.
[30] Guo Z, Kang S, Zhu X, Xia J, Wu Q, Wang S, Xie W, Zhang Y. The novel ABC transporter ABCH1 is a potential target for RNAi-based insect pest control and resistance management., 2015, 5: 13728.
[31] Tay W T, Mahon R J, Heckel D G, Walsh T K, Downes S, James W J, Lee S F, Reineke A, Williams A K, Gordon K H J. Insect resistance totoxin Cry2Ab is conferred by mutations in an ABC transporter subfamily a protein., 2015, 11(11): e1005534.
[32] Gahan L J, Pauchet Y, Vogel H, Heckel D G. An ABC transporter mutation is correlated with insect resistance toCry1Ac toxin., 2010, 6(12): e1001248.
[33] Atsumi S, Miyamoto K, Yamamoto K, Narukawa J, Kawai S, Sezutsu H, Kobayashi I, Uchino K, Tamura T, MitaK, Kadono-Okuda K, Wada S, Kanda K, Goldsmith M R, Noda H. Single amino acid mutation in an ATP-binding cassette transporter gene causes resistance to Bt toxin Cry1Ab in the silkworm,., 2012, 109(25): E1591-E1598.
[34] Chen L, Wei J, Liu C, Zhang W, Wang B, Niu L, Liang G. Specific binding protein ABCC1 is associated with Cry2Ab toxicity in., 2018, 9: 745.
Effect of Midgut Specific Binding Protein ABCC1 on Cry1Ac Toxicity against
CHEN Lin1, WEI JiZhen2, LIU Chen1, NIU LinLin1, ZHANG CaiHong1, LIANG GeMei1
(1State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193;2College of Plant Protection, Henan Agricultural University, Zhengzhou 450002)
【】The objective of this study is to research the bindingcharacteristics of midgut protein ABCC1 from(HaABCC1) to Cry1Ac and the effect of ABCC1 on toxicity of Cry1Ac, and to clarify the role of HaABCC1 in the insecticidal mechanism of Cry1Ac. 【】To verify the binding ability of HaABCC1 with Cry1Ac, the sequence ofwas analyzed and the primers were designed, two heterologously transmembrane protein fragments of HaABCC1 were expressed, and then Ligand blot experiment was conducted. The changes of expression level ofand mortality of larvae exposed to Cry1Ac were tested after silencedby microinjected siRNA into the abdomen of 3rd instar larvae. The cell mortality changes of sf9 insect cell line treated by Cry1Ac were compared using cell bioassay method, after transfectedinto sf9 and confirmed the pAc-ABCC1 recombinant plasmid was successfully transferred into sf9. The difference of full-lengthbetween Cry1Ac-resistant strain (BtR) and susceptible strain (96S) was compared, the expression of【】The two HaABCC1 transmembrane fragments, TMD1 and TMD2, were successfully expressed inBL21 (DE3) cells, these two heterologously expressed HaABCC1 fragments could bind to actived-Cry1Ac. The expression level ofsignificantly reduced after injection ofsiABCC1 into abdomen of thelarvae. Compared with non-treated, DEPC water- and siEGFP-injected larvae, the mortality oflarvae which were knocked downsignificantly decreased after treated with actived-Cry1Ac. It was showed the toxicity of Cry1Ac againstlarvae significantly reduced after thesilence. Compared with sf9 control, the mortality of cell line whichexpressed in sf9 obviously increased after treated by actived-Cry1Ac. The result indicated that transfectioninto sf9 could significantly increase the cell mortality after Cry1Ac treatment. There was no difference in the cDNA sequence and amino acid sequence ofbetween susceptible strain (96S) and Cry1Ac-resistant strain (BtR), but the expression level ofsignificantly reduced in BtR resistant strain compared with 96S susceptible strain.【】HaABCC1 ofis a specific binding protein and may be a functional receptor protein of Cry1Ac, it may involve in the resistance ofto Cry1Ac.
; ABCC1;; Cry1Ac; binding protein; toxicity
10.3864/j.issn.0578-1752.2019.19.005

2019-04-23;
2019-05-27
国家转基因生物新品种培育重大专项(2016ZX08011-002)
陈琳,E-mail:chenlincaas@126.com。
梁革梅,E-mail:gmliang@ippcaas.cn
(责任编辑 岳梅)
猜你喜欢
贵州畜牧兽医(2023年3期)2023-06-29 07:07:28
农技服务(2023年2期)2023-03-15 00:43:08
园林科技(2020年2期)2020-01-18 03:28:18
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
新疆农垦科技(2016年2期)2016-08-21 13:50:22
现代农业(2016年5期)2016-02-28 18:42:35
现代农业(2016年4期)2016-02-28 18:42:09
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
现代农业(2015年5期)2015-02-28 18:40:42
