
海带钒依赖溴过氧化物酶基因甄别和表达分析*
2019-10-12 02:08:40丁海燕李晓捷杨官品
中国海洋大学学报(自然科学版) 2019年11期
丁海燕,李晓捷,郭 栗,杨官品
(1.中国海洋大学教育部海洋遗传育种重点实验室,山东 青岛 266003;2.中国海洋大学海洋生命学院,山东 青岛 266003;3.国家海藻海参工程科学与技术研发中心 山东 烟台 264003;4.海藻遗传改良和高效栽培山东省重点实验室,山东 烟台 264003;5.山东东方海洋科技股份有限责任公司,山东 烟台 264003;6.中国海洋大学海洋生物多样性与进化研究所,山东 青岛 266003)
不等鞭毛总门的褐藻与后鞭毛生物(如动物和真菌)和植物(植物、绿藻和红藻等)的进化存在平行性,两类生物有相似的先天免疫机制和胁迫防御反应机制[1-3]。先天免疫机制不同于胁迫防御应答机制。防御机制可经受体引发对病原体的抵抗[4-5]。最近,在模式褐藻长襄水云基因组中就找到了防御有关的受体[6]。过氧化物爆发导致局部活性氧簇(Radical oxygen species,ROS)浓度急剧升高,是防御应答机制之一。这一机制在人[7]、植物[8]和褐藻[2]中都存在。病原体感染导致膜释放十八碳(植物)和二十碳(动物)脂肪酸,用于合成氧脂素(Oxylipin)。海带同时存在这两类氧脂素生物合成途径[9-14]。
海带(Saccharinajaponica)属于囊泡藻界,不等鞭毛总门,褐藻纲,海带目,海带科,海带属。上1920—1930年代日本引入中国栽培。海带是孢子体世代和配子体世代交替型生活史,从幼嫩孢子体长为成熟孢子体的过程中面临诸多环境胁迫。中国海带栽培主要是“浮绳网藻体倒置避夏”栽培模式。海带的生长区直接面对强紫外线、强阳光、高温度和频繁的光照和温度波动,而藻体的梢部却指向弱光、相对低温的海底。海带栽培集约化程度高,面积大,藻体密集,经常与动物养殖空间重叠。大尺度、大范围的环境温度波动和海水酸化,海水富营养化也逐步演变成不可忽略的环境影响因子。这些因素单独或组合作用,导致海带孢子体处于持续性的免疫和防御状态。有报道称L.digitata受环境刺激后可识别其细胞壁多糖的分解碎片,触发局部过氧化物爆发[15-16],防御相关基因的表达,释放游离脂肪酸[17]、加强卤素代谢[18]。
褐藻是唯一利用无机碘抵抗细胞外氧化胁迫的生物。碘合成可高效去除过氧化物爆发产生的过量过氧化氢和过氧根离子[19]。催化卤素合成的钒依赖卤素过氧化物酶(Vanadium-dependent halogenperoxidase,vHPO)是一类过氧化物酶,依其对卤素离子催化效率的不同,分为钒依赖氯过氧化物酶(vCPO)、钒依赖溴过氧化物酶(vBPO)和钒依赖碘过氧化物酶(vIPO)三类。3种过氧化物酶中,vCPO只存在于陆地植物中,而另外两种存在于海洋藻类,尤其是褐藻中。研究发现vBPO基因能催化碘离子与过氧化氢反应生成次碘酸,而次碘酸进一步将碘离子氧化为碘分子。反应过程为:I-+ H2O2→ HIO + OH-;HIO + I-+ H+→ I2+ H2O。
vBPO在海带免疫应答和防御中发挥重要作用[2]。为弄清海带vBPO基因的表达模式随海带生长发育的变化关系及其与藻体碘含量的关系,探究海带在生长发育过程中免疫防御反应的变化,本研究通过转录组分析研究了正常栽培海带不同生长发育时期vBPO基因的表达模式。
1 材料和方法
1.1 RNA提取和转录组测序
本研究所用的海带孢子体为东方7号[20],分别在4个发育时期,即薄嫩期(Mushroom stage)、厚成期(Adult stage)、成熟期(Mature stage)和衰老期(Aging stage)采集两株,取其生长点组织,用无菌海水彻底清洗表面粘液后,在液氮中研磨成粉末,用除多糖多酚的RNA提取试剂盒提取RNA,溶于DEPC处理过的重蒸水,用NanoDrop测定浓度,经1%琼脂糖凝胶电泳检测RNA完整度和可能的基因组DNA污染。合格RNA迅速置于-80℃冰箱中保存。
用Oligo(dT)磁珠富集mRNA,片段化后用六碱基随机引物合成第一链cDNA,然后用DNA polymerase I和RNase H合成第二链cDNA,再用AMPure XP beads纯化双链cDNA。末端修饰并加A尾后与测序接头连接,再用AMPure XP beads选择片段大小,PCR扩增,用AMPure XP beads纯化PCR产物,富集150~200 bp的cDNA形成测序文库。将文库用Qubit2.0定量,稀释至1.5 ng/μL,用Agilent 2100检测片段大小,合格后用Q-PCR方法进行准确定量(文库有效浓度>2nmol/L),在Illumina HiSeq平台上测序。
去除含接头、不确定碱基比例大于0.1%、Q≤20碱基数≥50%的初始读序(raw reads)获得净读序(cleanreads)。将净读序用Trinity[21]进行拼接(k=25,最小kmer覆盖度=2)。用 FPKM方法校正转录本丰度,消除序列长度和测序深度影响[21],实现丰度标准化。
1.2 碘含量测定
与转录组测序用海带孢子体同步采集4个生长时期的东方7号海带孢子体,每时期3株,通风处晒干后用粉碎机粉碎成≤400目的颗粒,均匀混合后用NaNO2-尿素法测量碘的含量[22]。
1.3 海带vBPO基因表达模式分析
海带4个时期转录组测序获得的组装物(unigene,后续分析转称为基因)针对Nr数据库进行注释,根据注释信息获取海带vBPO基因及其对应各时期的转录本丰度。利用OmicShare tools2.0(www.omicshare.com)绘制vBPO基因表达热图。
1.4 海带vBPO分子系统学分析和结构域预测
从海带4个生长发育时期的转录组分析数据中提取vBPO基因的序列,用Unipro UGENE软件(http://www.x64bitdownload.com/download/t-64-bit-unipro-ugene-64-bit-download-jheuyjsm.html)获得其开放阅读框(openreadingframe,ORF)并推导其氨基酸序列。在NCBI蛋白质数据库中下载不同物种的vBPO序列。将这些序列用MAFFT(http://www.softpedia.com/get/Science-CAD/MAFFT.shtml)进行对位分析,然后用MrBayes3.2(http://mrbayes.csit.fsu.edu/)构建分子系统树。最后获取的系统树是1 000次重复计算的结果。为进一步确定海带vBPO基因,用NCBI的CD-Search(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)结构域预测工具对其进行了结构域的预测。
2 结果
2.1 海带vBPO基因表达模式
海带孢子体4个生长发育时期的转录组测序共获得196 238 671条净读序,组装成133 516条不重复基因(Unigene,以下改称基因)。这些基因中,只有33.84%可针对NR数据库进行功能注释,19.25%针对SwissProt数据库进行功能注释,32.95%针对PFAM数据库进行功能注释,33.06%针对GO数据库进行功能注释。另外,可注释的基因中,绝大部分是病毒、转座子等先关蛋白。海带是非模式物种,进化上与已知模式物种亲缘关系较远。这给我们基于基因的功能注释理解海盗生长发育时期的生理学变化造成巨大困难。
幸运的是针对NR数据库的功能注释获得59条可能编码vBPO的基因(见表1)。因为海带基因组序列碎片化严重,无参转录组分析可能使转录本组装也碎片化,我们转而将133 457个基因的平均丰度作为59个vBPO基因转录本丰度的参照。我们发现海带vBPO基因转录本平均丰度是相应生长发育时期的9~19倍,平均13倍。这说明,vBPO基因在海带孢子体生长发育过程中始终高表达,间接地反映出栽培海带孢子体很可能持续性处于胁迫响应生理状态。
海带孢子体不同发育时期,59个vBPO基因表达模式不同(见图1)。在薄嫩期、厚成期、成熟期和衰老期,这些基因的转录本丰度(表达量)存在显著差异。相对而言,vBPO基因在成熟期和衰老期的表达模式更相似,而薄嫩期和厚成期更相似。海带孢子体不同发育时期碘含量逐期增加,薄嫩期为(0.30±0.00)‰,厚成期为(0.38±0.07)‰,成熟期为(1.57±0.05)‰,衰老期为(2.32±0.09)‰。碘含量与vBPO基因平均表达量成正相关(相关系数0.974)。

表1 海带不同生长发育时期vBPO基因的表达丰度和转录组分析组装长度Table 1 The abundance and length of the kelp vBPO gene transcripts

续表1
注:基因转录本丰度用FPKM值表示;*,小数点位数原因,计为零。 表达模式分析是基于注释信息进行的。为进一步证实这些组装物是vBPO基因,我们将59个注释为vBPO基因中包含完整开放阅读框的4个vBPO基因(GenBank存取号:MH430667、MH430668、MH430669、MH430670)逆翻译(推导)成氨基酸序列,从NCBI数据库下载16个物种的28条vBPO氨基酸序列,用这些序列构建分子系统树。分析发现海带4条vBPO氨基酸序列与掌状海带的vBPO亲缘关系近,与所有褐藻vBPO氨基酸序列聚在一起。说明这4条海带vBPO基因确实是vBPO基因(见图2)。 对4条海带vBPO基因对应的氨基酸序列进行结构域预测,发现它们的结构域相同,第339~602位氨基酸组成PAP2_like蛋白结构域,具有组氨酸磷酸酶和钒依赖卤素过氧化物酶催化活性基团,第418~426位的KWQVHRMLR序列、从第493~495位的SGH序列和从第575~584位的RSHLGVHWRMD的序列是钒结合位置(见图2)。 (①Mushroom stage;②Adult stage;③Mature stage;④Aging stage。(A)vBPO基因表达丰度热图;(B)FPKM值≥10的vBPO基因表达丰度折线图;(C)FPKM值≥1但<10的vBPO基因表达丰度折线图;(D)FPKM值>0但<1的vBPO基因表达丰度折线图。(A)The heatmap of kelpvBPOgenes;(B)The line chart of kelpvBPOgenes with FPKM values≥10;(C)The line chart of kelpvBPOgenes with FPKM values≥1 but <10;(D)The line chart of kelpvBPOgenes with FPKM values>0 but <1.) 图2 从有完整开放阅读框的海带vBPO基因推导的氨基酸序列与其他物种vBPO的分子系统学关系(上)及结构域(下)Fig.2 Phylogenetic relationship of the deduced amino acid sequences from annotated kelp vBPO genes containing the whole open reading frame with those retrieved from GenBank(above)and their structural domain (below) PAP2_like蛋白是组氨酸磷酸酶和钒依赖卤素过氧化物酶等组成的酶超家族,有磷脂酸磷酸酶、磷酸酯磷酸酶、葡萄糖-6磷酸酶、叶绿体膜脂磷酰甘油磷酸酶B、钒依赖氯/溴过氧化物酶等亚家族,有些酶是跨膜蛋白。本研究中未发现海带vBPO的跨膜区。Colin等发现第409~418位氨基酸和第475~492位氨基酸是钒结合区[23]。第409~418位氨基酸为KWQVHRMLRP,这一区域与海带vBPO第418~427位氨基酸序列完全一致。结构域分析也证实注释为海带vBPO的基因确实是vBPO。 本研究用转录组测序的方法分析了海带孢子体包括薄嫩期、厚成期、成熟期和衰老期在内4个生长发育时期vBPO基因的表达模式,并通过分子系统学分析和结构域比较验证了海带的这些基因确实是vBPO基因;发现海带vBPO基因转录本在4个发育时期的平均丰度是相应时期全部基因转录本的平均丰度的9~19倍,平均13倍,说明海带天然免疫和防御应答机制之一的碘富集过程持续性处于活跃状态;发现海带vBPO基因的表达量与海带孢子体碘含量呈正相关。这些认知对海带抗病抗逆生理机制解析具有重要意义,是海带抗病抗逆机制研究的新尝试。 海带基因组测序已经测序[24]。同源性搜索已在发表的海带基因组中找到了丰富的vBPO基因拷贝。但是已发表的海带基因组序列片段化严重,达不到参考基因组的程度,搜索结果只能说明海带基因组存在丰富的vBPO基因。如果是有参转录组分析,我们就能了解不同基因的转录特征。但是,由于已发表基因组序列的局限性,我们可能遗漏有关基因。基于这样的考虑,我们选择了无参转录组分析。分析过程中发现多数转录本组装物不完整。这又可能影响平均表达丰度的计量。因此,参考基因组质量的海带基因组序列对转录组分析具有重要的意义。然而,我们不得不等待高质量基因组序列的发表。值得庆幸的是已经有用实时单分子测序(三代测序)技术结合二代测序测定海带孢子体基因组获得参考基因组质量的海带基因组序列的工作(个人了解)。这将为深入研究海带抗病抗逆机制提供背景数据基础。 多拷贝基因的表达调控效率是比较低的,海带需要在不同胁迫条件、不同发育时期选择不同拷贝基因的表达并协调这些基因的作用。海带不同vBPO基因结构、它们的表达模式及其与生长发育时期和不同胁迫条件的关系为深入研究海带多拷贝基因的系统表达提供了一个切入点。 海带vBPO基因转录本相对丰度是与全部转录本平均丰度比较获得的相对值。我们最初想选择四个时期表达量基本恒定的基因群的平均表达量作为内参来确定vBPO基因的相对表达量,但这些基因的表达量只是相对恒定。与vBPO基因一样,无参组装获得的这些基因的序列也存在不完整性。既然都不完整,且是在相同的分析条件下比较。我们选择了全部组装物的平均丰度做对照来确定vBPO基因的相对表达丰度。海带基因的表达分析也可以参考其他物种选择内参基因。基于相似的考虑,我们没有这样做。 需要特别强调的是,本研究的表达分析针对的是多拷贝基因。基因序列高度相似。在这样的情况下,实时定量PCR方法很难设计引物来区分不同的基因拷贝。但基于测序的转录组分析可以克服这一限制,实现不同基因拷贝的区分。在方法上,本研究将为多拷贝基因的表达分析提供一种方法学借鉴。 分子系统学和结构域分析证实转录组分析注释的海带vBPO基因确实是vBPO基因。海带vBPO基因存在多个拷贝,在4个时期持续性选择性地表达,且平均表达量远高于总体基因平均表达量。这说明现行栽培模式下,海带孢子体有可能处于持续性胁迫响应状态,为解释频发的生理性病害提供了一种可能性。转录组分析组装获得多条vBPO基因序列,这与海带基因组存在众多vBPO基因拷贝对应,但这些基因的结构、表达模式及其与生长时期和胁迫环境间的关系尚需进一步研究。
Note:The abundance of genes are expressed as the FPKM value;*,due to2.2 海带vBPO分子系统学分析及结构域预测
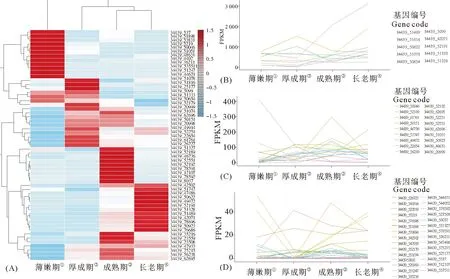
图1 海带vBPO基因在四个发育时期的表达模式
Fig.1 The expression abundance heatmap of kelpvBPOgenes during its growth and development
3 讨论
4 结语
猜你喜欢
飞碟探索(2023年3期)2023-11-23 16:08:36
飞碟探索(2023年1期)2023-05-30 22:59:34
黑龙江大学自然科学学报(2022年4期)2022-11-17 08:07:52
河北科技师范学院学报(2022年2期)2022-08-26 08:55:46
第一财经(2020年10期)2020-10-15 03:17:55
第一财经(2019年7期)2019-07-25 17:48:18
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
中国塑料(2016年3期)2016-06-15 20:30:00
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
