
小粒野生稻渗入系抗南方水稻黑条矮缩病QTL分析及利用
2019-10-10 08:20李孝琼刘开强郭嗣斌
贵州农业科学 2019年9期
韦 宇, 李孝琼, 陈 颖, 刘开强, 郭嗣斌*
(1.广西壮族自治区农业科学院 水稻研究所/广西水稻遗传育种重点实验室, 广西 南宁 530007; 2.广西壮族自治区农业科学院, 广西 南宁 530007)
水稻病毒病是水稻三大病害之一,有水稻“癌症”之称,严重威胁着我国的粮食安全。与普通水稻黑条矮缩病不同,南方水稻黑条矮缩病是一种以白背飞虱为传播媒介的水稻病毒病,其病原病毒为南方水稻黑条矮缩病毒(Southern rice black-streaked dwarf virus, SRBSDV),是呼肠孤病毒科(Reoviridae)斐济病毒属(Fijivirus)第2组的一个新种[1],该病于2001年在广东省阳西县晚稻田中首次被发现,在短短的几年间病情蔓延迅猛,给我国南方广大稻区及越南北部水稻生产造成了极大损失,甚至绝收[2]。目前,对南方水稻黑条矮缩病毒病的防治仍以防治飞虱为主,但长期施药易使飞虱产生抗药性,导致病害再次爆发[3-4]。实践证明,发掘并利用新抗源培育抗病新品种是控制水稻病害最经济有效和最环保的措施。挖掘南方水稻黑条矮缩病抗性资源、鉴定和定位抗性基因是当前我国南方稻区水稻抗病育种的当务之急。
国际上对由SRBSDV引起的南方水稻黑条矮缩病的研究主要集中在病毒检测、分子生物学特点、发生危害及综合治理等方面[2-11],抗源鉴定与评价工作目前还处于起步阶段,仅有几篇关于抗性材料筛选的报道[12-15]。研究表明,生产上尚未普遍使用抗性品种来防治南方水稻黑条矮缩病病害,主要问题是抗性鉴定及品种培育存在技术困难,且耗时过长[16]。迄今为止,对南方水稻黑条矮缩病抗性的遗传研究及抗性基因挖掘研究仅见零星报道[17-18]。农保选等[17-18]对419个广西地方稻种资源核心种质的SRBSDV苗期抗性进行全基因组关联分析,共检测到8个抗病基因(R基因),分别位于第1、第5及第11染色体上,可能参与SRBSDV苗期抗性的调控;利用来源于普通野生稻高抗SRBSDV的导入系D4为材料,将与水稻SRBSDV苗期抗性相关的主效QTL定位于第9染色体上102.3 kb处,将其命名为qSRBSDV9。
在前期研究中,广西壮族自治区农业科学院水稻研究所利用1份抗南方水稻黑条矮缩病的小粒野生稻(Oryzaminuta)基因渗入系桂恢1561和高感病品种桂1025杂交构建F2为作图群体,绘制了遗传连锁图谱,并完成了重组自交系(Recombinant inbred lines, RILs)的构建。为南方水稻黑条矮缩病的抗性育种提供新的基因资源,笔者等对桂1025/桂恢1561的重组自交系群体进行南方水稻黑条矮缩病抗性鉴定,对其抗性进行遗传分析,并利用QTLNetwork-2.2软件对RILs的南方水稻黑条矮缩病抗性 QTL 进行定位,旨在为南方水稻黑条矮缩病抗性育种提供基础材料和理论依据。
1材料与方法
1.1试验材料
桂恢1561来自小粒野生稻基因渗入系,抗南方水稻黑条矮缩病;桂1025为三系杂交稻恢复系,高感南方水稻黑条矮缩病。2010年晚季广西壮族自治区农业科学院水稻研究所以桂恢1561(抗病亲本)为父本,以桂1025(感病亲本)为母本,经杂交、自交,获得F2遗传分离群体,并构建了包含210份个体的重组自交系群体(F8)。重组自交系种植于专用病圃进行抗性鉴定,感病对照 Taichung Native (TN1) 由中国水稻研究所提供,广西农业科学院水稻研究所繁殖和保存。抗病对照中浙优8号,由浙江勿忘农种业股份有限公司生产。
1.2试验方法
1.2.1南方水稻黑条矮缩病抗性鉴定南方水稻黑条矮缩病是以白背飞虱为传播媒介的水稻病毒病,其病原病毒为南方水稻黑条矮缩病毒。选择广西壮族自治区兴安县湘漓镇麦源村的病圃(东经110°14′,北纬25°17′)为鉴定圃,该病圃位于我国白背飞虱迁飞的重要通道,虽然白背飞虱发生程度年度间有差异,但各年均有发生[20-21]。鉴定圃的白背飞虱量越大,其南方水稻黑条矮缩病抗性鉴定越有效,鉴定效果越可靠。抗性鉴定参照于文娟等[15]的鉴定方法,于2016年4月26日播种感病对照品种TN1、抗性对照中浙优8号及210份重组自交系,秧田较常规管理增施尿素75 kg/hm2,苗期不使用杀虫剂和杀菌剂。
6月6日移栽,将TN1(感病对照品种)作诱发南方水稻黑条矮缩病材料,TN1种植小区每行单本移栽6株(株距 15 cm、行距18 cm)。重组自交系穿插于TN1小区之间种植,每个重组自交系种植5行,每行10株(株距 15 cm、行距18 cm),随机区组排列,重复3次。抗病对照中浙优8号每个重复种植1个小区,种植规格同重组自交系。大田管理较常规肥水管理增施尿素150 kg/hm2,除拔节初期用杀虫双防治1次稻纵卷叶螟外,其他生长期不使用杀虫剂和杀菌剂。
移栽50 d后采用盘拍法调查对照及重组自交系材料的白背飞虱虫量。在黄熟期,将明显矮缩株和具高位分蘖的单株记为病株,调查各小区内50丛水稻的死亡株和存活的病株数。由于病圃内褐飞虱发生较重而引起部分植株枯死,为排除褐飞虱引起的枯死株影响,计算时以存活单株的矮缩株除以存活株总数计算南方水稻黑条矮缩病发病率。发病率= (存活病株数/存活株数) ×100%。以3个重复的发病率平均值作为重组自交系各株系的抗南方水稻黑条矮缩病表型值。抗性分级标准:1 级,发病率0.1%~5.0%,高抗(HR);2级,发病率5.1%~15.0%,中抗(MR);3级,发病率15.1%~30.0%,中感(MS);4级,发病率30.1%~50.0%,感(S);5级,发病率>50.0%,高感(HS)。依照上述方法,2017年4月进行重复试验,但由于当年自然灾害的原因,导致试验失败,因此,2018年4月再次进行重复鉴定,获得2年重组自交系对南方水稻黑条矮缩病的抗性表型数据。
1.2.2重组自交系基因型数据分析2015年9月至2016年12月,提取210份重组自交系材料的DNA,获得其基因型。DNA提取、PCR反应、电泳和银染检测方法与前期研究相同[22]。选取在双亲间具有多态的202对均匀分布于水稻12条染色体上的SSR引物,对桂1025/桂恢1561的重组自交系210个株系材料进行全基因组标记分析,用JoinMap 4.0作遗传连锁图。
1.2.3南方水稻黑条矮缩病抗性QTL分析结合重组自交系的表型数据和基因型数据,参照YANG等[22]的方法利用软件QTLNetwork-2.2对RILs的南方水稻黑条矮缩病抗性进行QTL定位,分析各个抗性QTL的遗传效应,预测最优的基因型值。
1.2.4对鉴定出的抗性较好的株系材料进行育种评价和利用选择部分南方水稻黑条矮缩病抗性较好的株系材料作父本,分别与天丰A、丰田A和华浙2A等不育系配制杂交稻组合,对杂交组合进行试种和抗病鉴定,考查其配合力和抗病表现,评价其育种价值。同时利用抗病亲本桂恢1561和部分重组自交系抗性株系为供体亲本,以广恢998、华占和桂恢553等优良恢复系为受体,进行回交转育和分子标记辅助选择,分离世代种植于广西兴安县湘漓镇麦源村的病圃进行自然诱发鉴定,选育新的抗南方黑条矮缩病育种材料。
2结果与分析
2.1南方黑条矮缩病鉴定结果的可靠性
由于南方水稻黑条矮缩病是以白背飞虱为传播媒介的水稻病毒病,南方水稻黑条矮缩病鉴定区域白背飞虱量越大,其抗性鉴定越有效,鉴定效果越可靠。调查结果表明,2016年和2018年的水稻分蘖期感病对照TN1白背飞虱平均虫量分别为35.2头/株和43.6头/株,白背飞虱虫量充足,说明研究区域具备南方黑条矮缩病鉴定的外部条件。水稻黄熟期感病对照TN1各重复的病株率均为100%,抗病对照中浙优8号平均病株率小于5%,表现抗病,表明水稻南方黑条矮缩病田间发病充分,抗性鉴定结果可靠。
2.2重组自交系的抗性表现
对重组自交系各株系、抗性亲本桂恢1561、感病亲本桂1025的南方水稻黑条矮缩病调查结果表明,桂恢1561表现高抗,桂1025表现高感,210个重组自交系株系表现为偏正态连续性分布(图1),表明桂恢1561的南方水稻黑条矮缩病抗性可能受主基因和微效基因共同控制。根据南方水稻黑条矮缩病抗性分级标准,2016年和2018年从210份重组自交系中鉴定出抗及中抗的株系材料共计31份。
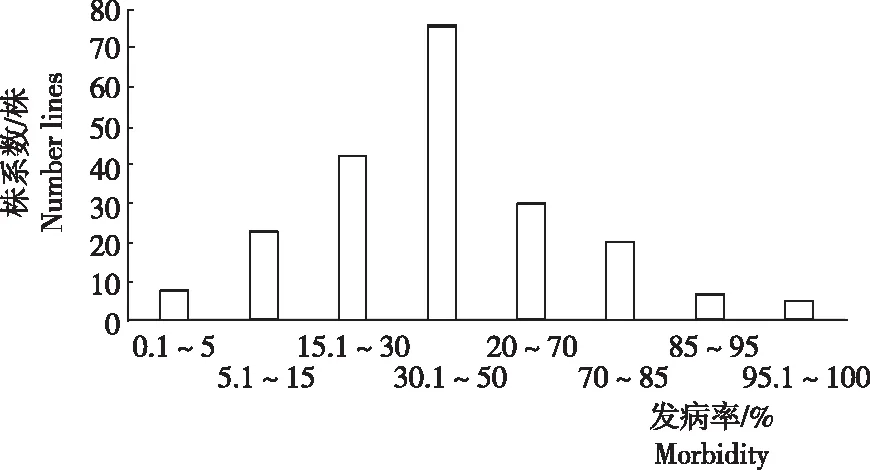
图1 重组自交系南方水稻黑条矮缩病田间抗性频次分布
Fig.1 Frequency distributions of the SRBSDV-resistance of RILs
2.3构建高密度遗传图谱
从图2看出,桂1025/桂恢1561RILs构建的水稻遗传连锁图中包括185个SSR标记,13个连锁群,覆盖了水稻全基因组1 202.5 cM的区域,标记平均间距6.5 cM。
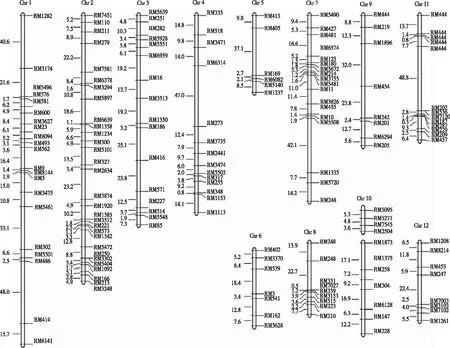
图2 桂1025/桂恢1561 RILs构建的水稻遗传连锁图
2.4QTL的分析结果
从表1看出,2016年共检测到4个抗病QTL,分布于第3、4、7和12号染色体上,表型贡献率为6.5%~36.3%;其中效应值最大的QTL是qSRBSDV4,其位于第4染色体RM3471~RM518区间。来自桂恢1561的等位基因可降低病株率19.2%,表型贡献率为36.3%,可能是主效基因。
2018年共检测到3个抗病QTL,分布于第3、4和12号染色体上,表型贡献率为8.7%~37.5%。其中效应值最大的QTL是qSRBSDV4,其位于第4染色体RM3471~RM518区间,来自桂恢1561的等位基因可降低病株率20.1%,表型贡献率为37.5%,与2016年的结果较一致。因此,初步确定qSRBSDV4为抗南方水稻黑条矮缩病的主效QTL位点。
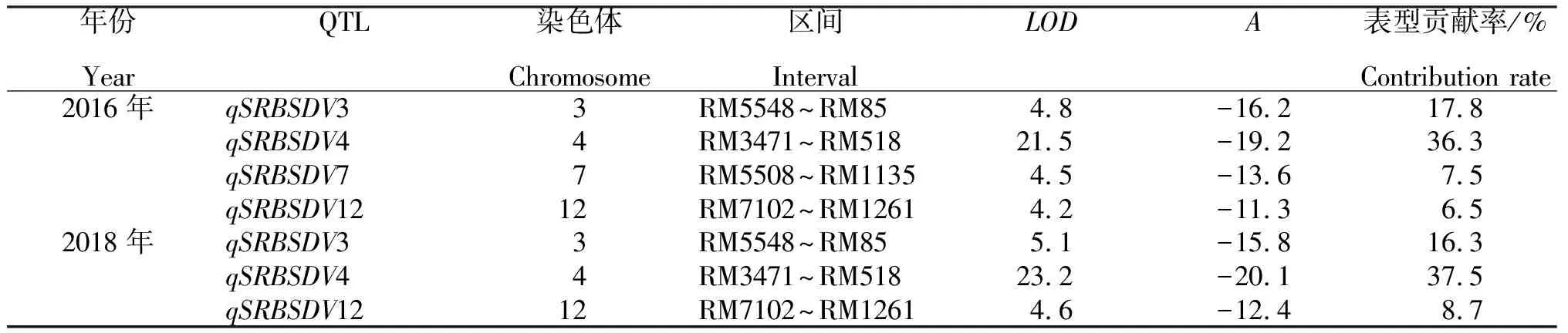
表1 2016年和2018年RILs抗南方水稻黑条矮缩病的QTL检测结果
注:表中A为被检测QTL的加性效应值,其负值表示抗性等位基因来自小粒野生稻。
Note: A is the additive effect value of QTL, and its negative value indicates that the resistance allele comes fromO.sativa.
2.5抗性株系材料的育种价值评价及利用
2017年早季从桂1025/桂恢1561的RILS中选择14份抗南方水稻黑条矮缩病的株系材料作父本,分别与天丰A、丰田A和华浙2A等不育系配制42个杂交稻组合。对42个杂交组合于2017年晚季在南宁试种,考察产量性状,并于2018年4月对42个杂交组合进行南方水稻黑条矮缩病抗性鉴定。综合抗性鉴定结果及考种结果,筛选得到7个配合力及抗性表现较好的株系材料(表4),分别为GK1、GK3、GK5、GK6、GK7、GK11和GK13。
2015-2018年,以抗病亲本桂恢1561和部分重组自交系抗性株系为供体亲本,以广恢998、华占和桂恢553等优良恢复系为受体,进行回交转育和分子标记辅助选择,分离世代种植于广西兴安县湘漓镇麦源村的病圃进行自然诱发鉴定,获得BC2F3、F4和F5等各世代株系材料187份。2018年对187份株系材料进行抗性鉴定,筛选出农艺性状基本稳定且对南方水稻黑条矮缩病抗性较好的株系材料32份。

表2 7个抗性材料所配部分杂交组合产量相关性状表现
3结论与讨论
研究从210份桂1025/桂恢1561的重组自交系株系中鉴定得到31份对南方水稻黑条矮缩病中抗以上材料,其中7份株系配合力较高;利用重组自交系定位到4个抗性QTL,qSRBSDV3位于第3染色体RM5548~RM85区间,qSRBSDV4位于第4染色体RM3471~RM518区间,qSRBSDV7位于第7染色体RM5508~RM1135区间,qSRBSDV12位于第12染色体RM7102~RM1261区间,其中qSRBSDV4为主效抗性位点。利用鉴定得到的抗性材料与部分恢复系材料进行杂交选育,获得了一批抗性育种基础材料。
野生稻长期在自然环境下生长,具有丰富的遗传多样性和对病虫害的抗性,是宝贵的水稻病虫害抗源。研究中重组自交系的亲本之一桂恢1561来源于四倍体的小粒野生稻,其所蕴含的抗逆基因已在多项研究中得到证实[23-24]。近年来,南方水稻黑条矮缩病给水稻生产造成了巨大经济损失,从小粒野生稻资源中发掘并利用新抗源培育抗性品种是防治该病的有效途径之一。
病圃所在的广西壮族自治区兴安县湘漓镇麦源村,地处水稻飞虱自华南向长江流域迁移的“湘桂走廊”,历年均有白背飞虱发生[25],于文娟等[15]研究表明,该地区也可能是SRBSDV在我国重要的毒源积累和繁殖地。在兴安病圃进行的田间自然传毒鉴定法相对于人工接种鉴定法,避免了室内人工接种抗病性鉴定中用到的白背飞虱不易捕捉、不易大批量繁殖等缺点[12],是一种简便易行且经济的鉴定方法。该病圃虫量充足,发病充分,抗性鉴定结果可靠。
目前,部分学者仅开展了抗源鉴定与评价工作,虽获得一些抗南方黑条矮缩病的材料[12-15],但关于SRBSDV抗性相关的基因或QTL挖掘方面研究不多。农保选等[17]从广西地方稻种资源核心种质中检测到8个SRBSDV苗期抗性抗病基因,分别位于第1、第5及第11染色体上;农保选等[18]从普通野生稻导入系中定位到1个水稻SRBSDV苗期抗性相关的主效QTL,将其命名为qSRBSDV9,其位于第9染色体上102.3 kb处。研究发掘出的抗南方水稻黑条矮缩病的主效QTL位点qSRBSDV4位于第4条染色体上,与目前已报道的SRBSDV抗性QTL[17-18]不在同一条染色体上,因此认为qSRBSDV4是抗南方水稻黑条矮缩病的新位点,下一步将利用SLAF-seq技术和HighMap软件对桂1025/桂恢1561的RIL开发高密度分子标签,利用该高密度遗传连锁图对主效QTL抗性位点qSRBSDV4进行精细定位,开发与目标基因紧密连锁的标记,争取实现基因的直接选择和有效聚合,大幅缩短育种年限,提高育种效率。
猜你喜欢
作物学报(2022年11期)2022-08-31
亚热带植物科学(2022年1期)2022-05-17
海峡科学(2021年6期)2021-08-04
园艺与种苗(2021年6期)2021-07-19
作物研究(2020年5期)2020-12-08
四川农业科技(2019年5期)2019-07-01
北京农学院学报(2019年1期)2019-02-22
河南农业科学(2017年4期)2017-04-12
环境昆虫学报(2016年6期)2016-12-27
农民致富之友(2016年5期)2016-10-21
