
融合黄瓜光质需求的设施光环境智能调控模型
2019-10-10 02:50:16荆昊男李远方张仲雄张海辉
农业机械学报 2019年9期
胡 瑾 荆昊男 高 攀 李远方 张仲雄,3 张海辉,3
(1.西北农林科技大学机械与电子工程学院, 陕西杨凌 712100;2.农业农村部农业物联网重点实验室, 陕西杨凌 712100;3.陕西省农业信息感知与智能服务重点实验室, 陕西杨凌 712100)
0 引言
黄瓜是一种重要的设施农业作物,光环境是影响黄瓜生长发育的重要因素,其不仅与光照强度有关[1-6],也与光质比(红光占总光强的比例)密切相关。研究证明,合理调节光质比能显著提升作物品质[7-8]。KHATTAK等[9]研究表明金鱼草植物高度、节间长度和叶面积受光质影响较大。MIAO等[10]证明蓝光比红光更有利于促进黄瓜叶片光合电子传递。王君等[11]证明采用不同红蓝配比光处理的生菜其光能利用效率存在明显差异。刘彤等[12]研究发现在不同条件下,植物光合作用所需的红蓝光强度比例也存在不同。但是目前大多数光环境调控的模型和系统研究只涉及到固定的光质比调控[13-15],近期也出现了考虑环境对光质影响的研究,其一定程度上提高了调控效果[16],但由于设置的试验条件不足,以及算法限制等问题,并没有进一步进行光质动态连续调控模型和方法的研究,造成现有设施光环境调控模型通常只考虑了固定光质比下的光强精准调控,尚未引入环境动态变化下的融合光质比和光强的协同调控方法,影响光环境调控效果。所以,建立融合作物光质需求的设施光环境智能调控模型,是设施农业环境调控急需解决的问题之一。
本文拟构建融合黄瓜光质需求的设施光环境智能调控模型。首先基于试验数据,利用回归型支持向量机(Support vector regression algorithm, SVR)算法建立融合黄瓜光质需求的光合速率预测模型,然后利用粒子群算法寻找特定温度下最大光合速率点对应的光照强度和光质比,最后基于寻找的数据建立光合速率调控目标值模型。
1 材料与方法
1.1 试验材料
试验于2019年4月在西北农林科技大学北校区农业物联网重点实验室进行,供试的黄瓜品种为“金胚98-1F1”,试验期间,对试验材料在培养箱进行正常的培育,不喷洒任何农药和激素。本试验主要设计了不同光质比、光强和温度耦合的嵌套试验,选取长势优良的黄瓜植株60株,采用LI-6800型便携式光合速率测试仪测定样本净光合速率。为了避免因天气状况不同和植株“午休”对试验结果造成影响,选择较为晴朗的天气,于08:30—11:30,14:30—17:30期间进行试验。利用光合速率测试仪自带的多个子模块设置不同的环境条件进行试验,其中,CO2浓度控制在400 μmol/mol,控温模块设定18、21、24、27、30、33℃共6个温度梯度,LED光质比设定为0.1、0.3、0.4、0.5、0.6、0.7、0.9共7个梯度,LED光源设定为0、30、75、150、300、500、650、800、1 000、1 200、1 400、1 600、1 800 μmol/(m2·s)共13个光照强度梯度,获取以温度、光质比、光照强度为输入,黄瓜净光合速率为输出的546组样本数据。
1.2 融合光质需求的光合速率预测模型建立
智能算法可有效提升模型的预测效率[17-19], 相关研究发现BP神经网络和随机森林算法的运算效率和预测精度一般不及支持向量机算法[20],因此本文采用回归型支持向量机进行模型构建。该模型以温度、光质比、光照强度为输入,黄瓜净光合速率为输出,选用高斯核函数进行数据与特征空间之间的映射,最后在特征空间构建最优回归平面,完成模型建立,其流程图如图1所示。
(1)由于不同样本数据之间的差距比较大,量纲也有所不同,故先对全体样本数据进行归一化处理,将数据统一到同一个区间[0,1]之内,所用的归一化计算式为
y′=(x-xmin)/(xmax-xmin)
(1)
式中y′——归一化之后的数据
xmin——同一量纲数据中的最小值
xmax——同一量纲数据中的最大值
(2)从归一化后的546组数据中,随机选取437组数据(占样本总数据的80%)作为训练集数据构建SVR模型,其余的109组数据(占样本总数据的20%)作为测试集数据以验证模型的泛化能力。
(3)本研究样本数据之间存在明显的非线性关系。对于非线性支持向量机问题来说,首先需选取合适的核函数,将数据映射到特征空间,然后在特征空间里利用线性支持向量机方法寻找最优回归平面。本文采用高斯核函数进行数据的映射变换,并采用网格迭代寻优方法进行惩罚因子c和核函数参数g优化。其中,c控制对超出误差样本的惩罚程度,g决定了数据映射到新的特征空间后的分布,与函数的宽度参数有关。本文经过迭代寻优得到最优的c为17,g为5。其中高斯核函数的表达式为
(2)
式中δ——函数宽度参数,控制函数的径向作用范围
x——训练集函数的自变量
xi——核函数中心
映射之后在特征空间内利用拉格朗日函数求解回归平面,所得最优回归平面为
(3)

(4)
(4)以测试集实测净光合速率为实测值,模型计算值为预测值,进行光合速率模型泛化能力和相关性比较。
1.3 基于粒子群算法的最优光照强度、光质比获取
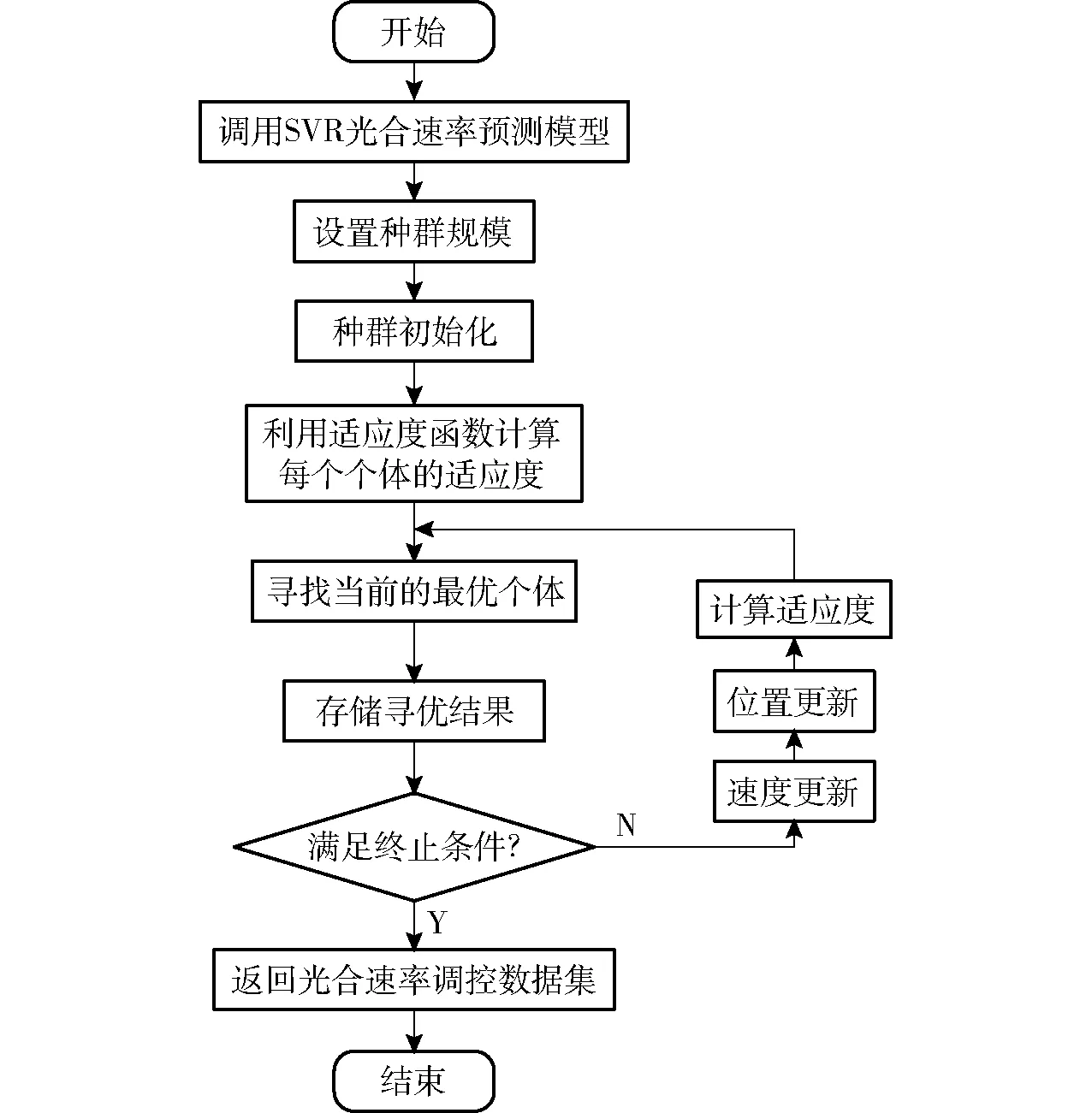
图2 粒子群算法寻优流程图Fig.2 Flowchart of particle swarm optimization algorithm
为了寻找特定温度条件下的光饱和点和最佳光质比,本文采用粒子群算法进行寻优。粒子群算法是一种基于迭代的优化算法,具有操作方法简单,寻优速度快的特点[21]。该算法基于群体中个体位置信息和食物位置信息连续发送的机制,利用信息共享完成个体移动方向和移动速度调整,从而实现群体不断地从杂乱无序的运动变为有序的活动,完成从随机解出发寻找最优解的过程。基于上述思想,本文采用光合速率预测模型实例化的方法建立不同温度下的寻优适应度函数,利用粒子群算法获取不同温度下净光合速率最大点,以及对应的光照强度和光质比,其流程图如图2所示,寻优具体过程包括以下步骤:
(1)进行特定温度下的光合速率预测模型实例化,并以此构建寻优目标值函数。
(2)确定模型参数,其中种群规模(即算法所需粒子的数量)为60,算法最大迭代次数为300,粒子的位置范围为[0,1],速度范围为[-0.005,0.005],粒子的个体学习因子为0.5,社会学习因子为1.5,惯性权重为0.5。
(3)初始化每个粒子的位置和速度,并以每一点对应的光合速率预测值作为适应度函数进行计算。
(4)通过比较每个粒子的适应度寻找初始化后群体中的最优位置及其适应度值,在此基础上进行迭代寻优,使每个个体都向最优解的方向运动,直到寻找到最优解。
(5)当满足寻优结束条件,即运行次数达到该算法设置的最大迭代次数后自动停止,并记录和保存所寻找到的最优解结果。
1.4 光合速率调控目标值模型建立
粒子群寻优算法寻找了特定温度下光合速率最优点对应的光质比和光照强度,基于此寻优结果,利用偏最小二乘回归法构建黄瓜的光合速率调控目标值模型,主要包括以下步骤:
(1)基于粒子群算法寻找到的特定温度下最大光合速率对应的光质比和光照强度,分别计算对应的红、蓝光的光照强度,计算式为
Pr=PL
(5)
Pb=P(1-L)
(6)
式中Pr——最优点红光光照强度
Pb——最优点蓝光光照强度
L——最优光质比
P——最优总光照强度
以温度为输入,对应的红光和蓝光的光强为输出,采用最高阶次为三次的偏最小二乘回归法,进行模型拟合,得到的调控模型表达式为
y=at3+bt2+ct+d
(7)
式中a、b、c、d——调控曲线系数
t——环境温度
y——红光或蓝光需求量
2 试验结果与分析
2.1 试验结果
分析了温度、光照强度、光质比与净光合速率的关系,得到部分试验曲线如图3所示。由图3可知,在相同温度下,随着光照强度的增大,黄瓜净光合速率总体趋势增大,当光照强度较低时,黄瓜的净光合速率随光照强度增长而快速上升,到达一定值后增长速度变缓,甚至会出现下降。而且光质比对黄瓜净光合速率也有显著影响,在不同温度下其最佳光质比不一致,整体来说,随着光质比的增加,光合速率呈先增后减的趋势。因此,建立光质比动态调控可以在一定程度上优化调控效果。
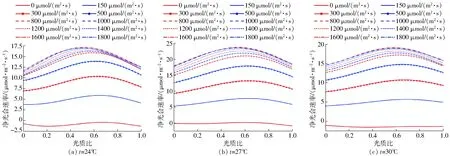
图3 不同温度下黄瓜净光合速率与光照强度、光质比的关系Fig.3 Relationships between net photosynthetic rate,light intensity and light quality of cucumbers
2.2 光合速率预测模型验证
为了验证SVR光合速率预测模型的性能,同时选取BP神经网络算法、随机森林算法(Random forest, RF)建模进行对比,3种模型的评价指标对比如表1所示。

表1 3种模型的拟合度和均方根误差对比Tab.1 Comparisons of fitness and root mean square errors of three models
由表1可知,利用SVR算法建立的模型训练集数据和测试集数据的拟合度分别为0.997 1与0.996 9,均高于BP神经网络算法和RF算法建立的模型;进一步对比均方根误差可知,随机森林算法在模型建立过程中出现过拟合,虽然训练集的均方根误差最小,但测试集的均方根误差较大。综合考虑,选用SVR算法能更好地反映数据之间的关系。
为了观察SVR预测模型的泛化能力,选用测试集的109组样本数据对模型的泛化能力进行验证。实测值和计算值的相关性如图4所示。结果表明,测试集拟合度为0.996 9,实测值和计算值两组数据之间具有较高的相关性。
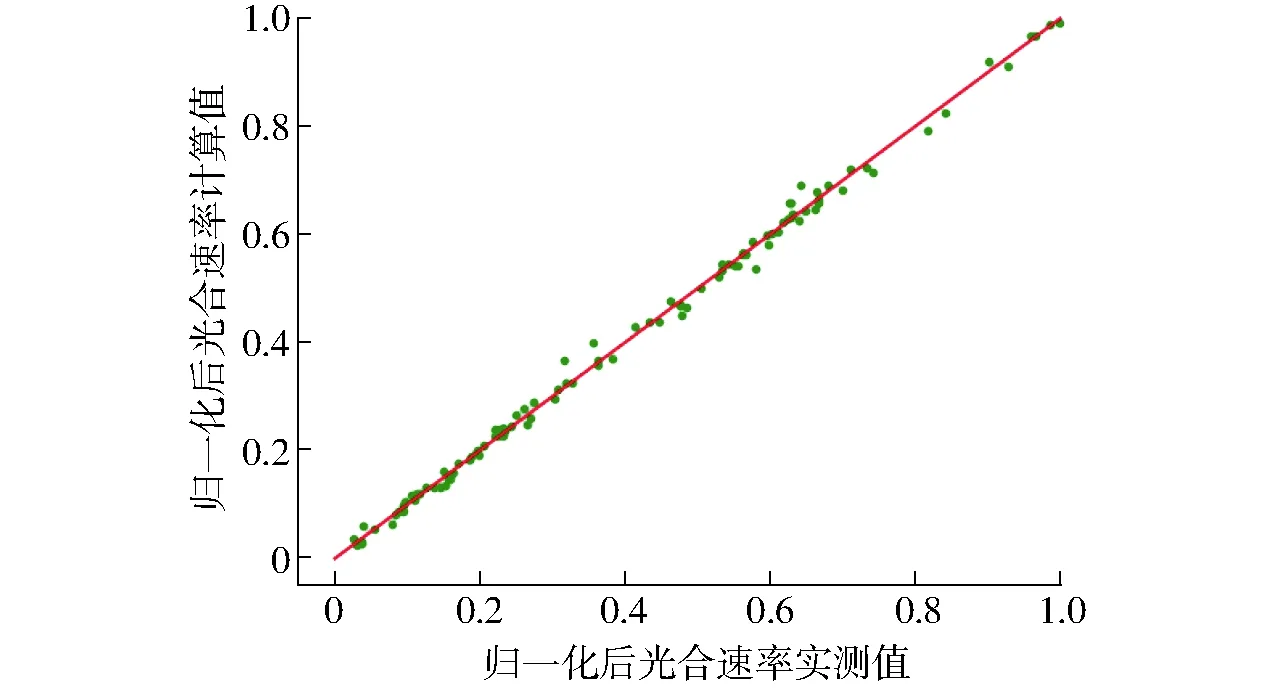
图4 光合速率预测模型相关性分析Fig.4 Correlation analysis of photosynthetic rate prediction model
2.3 粒子群寻优结果
利用粒子群算法完成了特定温度下净光合速率最大点的寻优,其中18、21、24、27、30、33℃条件下的寻优迭代过程如图5所示。由图5可知,粒子群算法寻优过程中无振荡,很快便可以收敛到最优点,证明该方法对于此类样本寻优适合。得到最优点数据如表2所示。
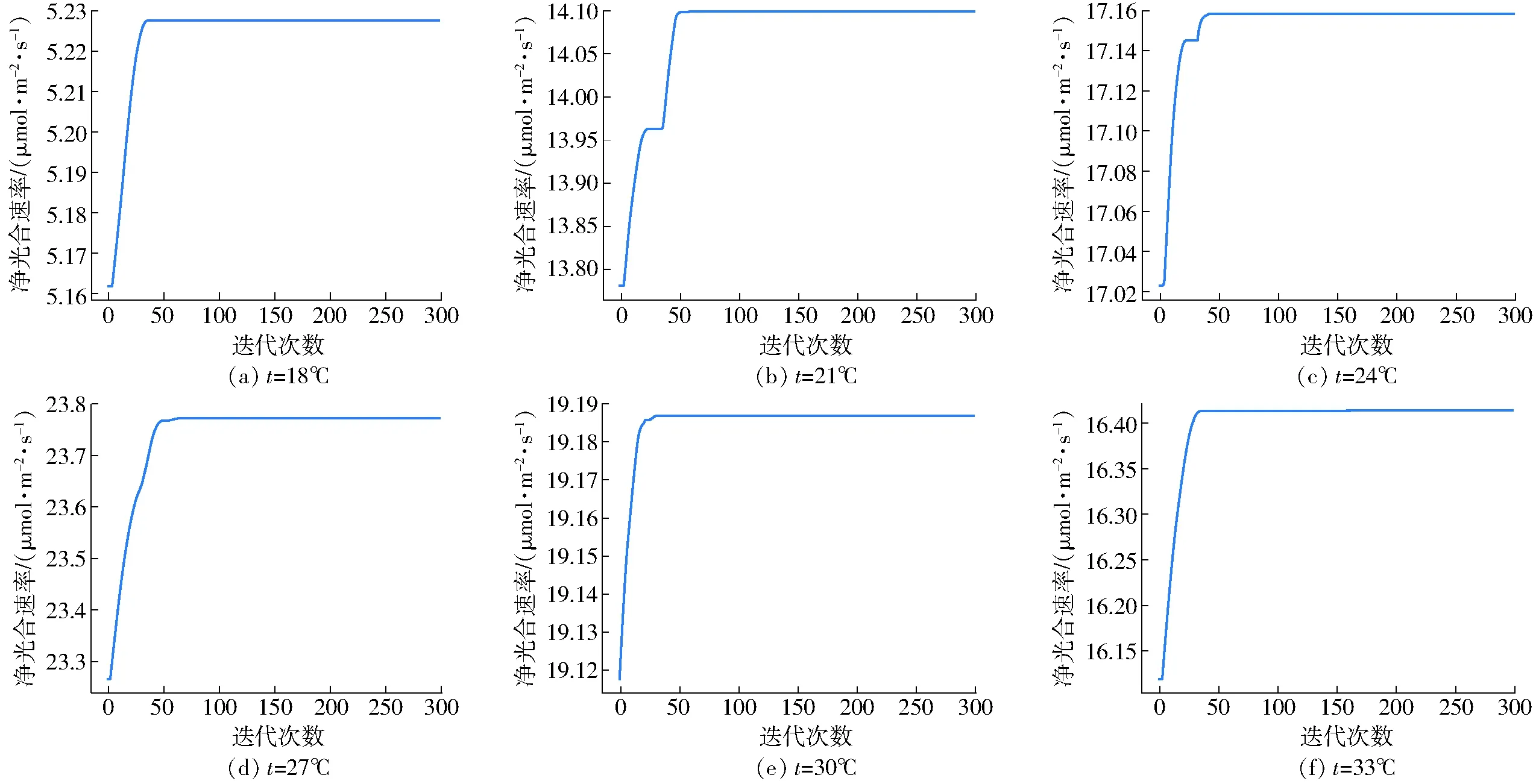
图5 粒子群算法寻优迭代过程Fig.5 Processes of particle swarm optimization

表2 光强与光质比寻优结果Tab.2 Values of optimal points for light intensity and light quality
由表2可知,在不同温度条件下,最大光合速率、光饱和点以及最佳光质比都有所不同。随着温度的升高,3种变量的变化趋势都是先增后减,其中最大光合速率、光饱和点大约在27℃附近达到最大值,而最佳光质比在30℃附近达到最大值。进一步分析发现,温度在21~33℃范围时,最佳光质比约为0.5,但低温会使最佳光质比较大程度地偏离0.5,18℃时寻找到的最佳光质比只有0.353 9,说明在较低的温度下蓝光可能更有利于提升作物光合能力。
2.4 调控模型验证
基于光饱和点和最佳光质比寻优结果,利用式(5)、(6)分别计算特定温度下红、蓝光的需求量,然后利用偏最小二乘回归法进行拟合,得到红、蓝光的调控曲线如图6所示,得到的拟合曲线方程分别为
y=-0.138 321 296t3+6.996 413 49t2-
60.851 210 7t+176.653 100
(8)
y=0.071 543 518 5t3-3.925 050 00t2+
41.036 025 0t+1 101.351 70
(9)
红、蓝光调控模型均方根误差分别为14.635 1、10.192 0 μmol/(m2·s)。

图6 调控曲线Fig.6 Intelligent regulation model curves
由图6可知,在红、蓝光的调控模型中,红光需求量随温度的升高而先增后减,大约在28℃附近达到最大值;而蓝光需求量随温度的升高而先减后增,大约在30℃附近达到最小值。这是因为植物光合作用的主要能量来源于红光,在28℃附近,黄瓜的光合作用最强,此时所需的红光光照强度也最多。而刺激植物气孔张开是蓝光的一个重要功能,在低温和高温条件下作物会通过关闭气孔来调节光合作用,所以随着温度的升高,所需的蓝光光照强度先减小后增大,且在30℃附近,蓝光光照强度达到最小,此种变化趋势可能还与酶的活性等生理参数密切相关[22]。
为了进一步验证所得调控模型的准确性,本文进行了计算值与实测值相关性分析。通过对实际测量的9个温度下光响应曲面光合速率最大值寻优得到光饱和点和最佳光质比,并利用式(5)、(6)得到对应温度下红、蓝光实测值,同时利用式(8)、(9)获取对应温度下红、蓝光预测值,从而进行实测值与计算值的相关性分析,结果如图7所示。
由图7可知,通过相关性拟合得到的拟合度分别为0.834 5和0.853 5,红光调控模型相关直线的斜率为1.001 3,截距为1.612 2;蓝光调控模型相关直线的斜率为0.985 7,截距为9.526 0,红、蓝光调控模型的均方根误差分别为15.087 8、10.138 3 μmol/(m2·s),说明该调控模型计算值与实测值之间有较高的相关性,可满足实际红、蓝光高效调控需求。
2.5 模型调控效果分析
温度、光照强度、光质比都是影响黄瓜光合速率的重要因素,三者对黄瓜的光合作用存在明显的互作作用。为了验证耦合条件下,光质比对光合速率的影响,对比不同文献中常见的光质比与本模型得到的最佳光质比在相应光饱和点下的净光合速率,结果如表3所示。

表3 不同光质比对应的净光合速率Tab.3 Net photosynthetic rates at different light qualities μmol/(m2·s)
由表3可知,在利用粒子群寻优找到的最佳光质比下,黄瓜的净光合速率高于其他几种常见的光质配比。在这几种常用的光质比条件中,光质比为0.5时对应的净光合速率最高,当光质比超过0.5时,净光合速率随光质比的增大而减小。且由表2可知,不同温度条件下,适合黄瓜生长的最佳光质比不同,最佳光质比随温度的升高而先增大后减小。所以相比于0.5这一最优固定光质比,利用粒子群算法动态寻优得到的光质比在相同总光强条件下,其调控效果更优。
3 讨论
为了进一步分析不同光质比造成净光合速率变化的原因,对比了不同温度下光质比对气孔导度的影响,如图8所示。由图8可知,光照强度、光质比和温度对气孔导度有显著影响,相同光质比条件下,气孔导度随温度变化趋势与光合速率变化趋势保持一致,大约在27℃附近达到最大值。从图8可知,在较低温度(24℃)及较高温度(33℃)时,光质比的变化对气孔导度有显著影响,增加蓝光光强可以显著地影响气孔导度变化。27~30℃附近,在最适光质比范围内,增加蓝光光强对气孔导度的影响很小,可能是由于在此温度范围内,光质比不再是影响其变化的主导因素,此时光合速率的下降可能与酶的活性等生理参数更相关。同时,在相同温度和光照条件下,气孔导度随光质比的增加而先增后减[24],且在光质比约为0.5处达到最大值,该变化趋势与净光合速率的变化趋势一致。从而表明最佳光质比的动态变化很有可能是蓝光的气孔导度效应引起的。
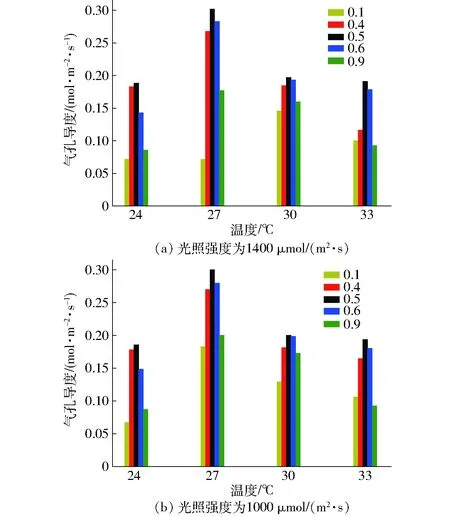
图8 不同温度下光质比对气孔导度的影响Fig.8 Effect of light quality on stomatal conductance at different temperatures
同时,光质比还会通过影响色素活性等生理参数进而影响其光合作用。相关研究发现红蓝光通过光敏色素和隐花色素共同调节光合作用。储钟稀等[25]研究了光质对黄瓜叶片光合作用特性的影响,发现红光处理的叶片具有较高的光系统Ⅱ(PSⅡ)活性和较低的光系统Ⅰ(PSⅠ)活性,蓝光处理的叶片具有较低的PSⅡ活性和较高的PSⅠ活性。而PSⅠ和PSⅡ是光合作用重要的两个过程,所以红蓝光比例过高或过低均会对植物的光合作用造成不利影响。王晓艳等[22]研究了不同比例的红蓝光对弱光下黄瓜叶绿体超微结构和相关光合特性的影响。结果显示,光质比为0.5处理组的比叶质量、净光合速率、气孔导度和光合性能指数均最高,分析得到光质比为0.5最有利于黄瓜幼苗叶绿体的发育和高光合速率的实现。其结果与表2中黄瓜的净光合速率随光质比的增大而先增后减,且在光质比约为0.5时达到最大值一致。
4 结论
(1)以温度、光质比、光照强度为输入,黄瓜净光合速率为输出,建立基于回归型支持向量机算法的融合黄瓜光质需求设施光环境智能调控模型。验证结果表明,该模型的拟合度为0.996 9,其均方根误差为0.436 7 μmol/(m2·s),明显优于BP算法和随机森林算法构建的模型,为设施光环境调控提供了重要依据。
(2)基于粒子群算法实现了不同温度下最佳光质比和光饱和点获取,其结果表明不同温度下最适合黄瓜生长的光质比存在差异,最佳光质比随温度的升高呈现先增后减的趋势,大约在30℃附近达到最大值。
(3)建立了红、蓝光目标值调控模型,其均方根误差分别为15.087 8、10.138 3 μmol/(m2·s),其调控效果明显优于传统固定的光质比,可用于指导实际应用中的光环境调控。
猜你喜欢
四川林业科技(2022年1期)2022-03-22 07:48:32
上海农业学报(2022年1期)2022-03-08 00:09:12
家庭影院技术(2019年8期)2019-08-27 02:45:04
家庭影院技术(2019年1期)2019-01-21 02:25:26
家庭影院技术(2018年10期)2018-11-02 05:35:38
新课程·下旬(2017年7期)2017-08-14 11:12:50
浙江农业学报(2017年1期)2017-05-17 06:13:36
中国果业信息(2015年12期)2015-01-24 07:31:03
西北林学院学报(2015年5期)2015-01-03 05:12:20
植物营养与肥料学报(2011年2期)2011-10-26 03:52:30
