
光强和光周期对水培生菜光合及叶绿素荧光特性的影响
2019-10-08 09:10:18胡笑涛王文娥方舒玲
西南农业学报 2019年8期
刘 杰,胡笑涛,王文娥,冉 辉,方舒玲,杨 鑫
(西北农林科技大学 旱区农业水土工程教育部重点实验室,陕西 杨凌 712100)
【研究意义】每种作物都有其适宜的光环境,因为四季、昼夜、海拔、纬度、天气及环境污染等因素,许多作物生长的自然光环境已经不能满足人们生产生活的需要。植物工厂的兴起,使作物生长环境更加可控,很大程度的解决了低温寡照、病灾虫害等作物生长的限制。但植物工厂对人工光的需求很大,光强和光周期直接影响植物工厂照明的电能消耗,也关系到作物光合作用、光能利用等情况[1-2]。生菜(LactucasativaL.)作为常见的特优蔬菜,是植物工厂的主栽作物,因此,研究光强和光周期对生菜光合特性和生长的影响对减少植物工厂电能消耗,提高作物生产效率具有重要意义。【前人研究进展】近年来,有学者通过气体交换参数、叶绿素荧光手段、同位素、酶活性、气孔等方法研究了光强和光周期对植物光合特性、光能利用的情况,并得到了许多光强、光周期响应规律,揭示了相关机理[3-7]。已有研究表明一定范围内增加光强,作物的净光合速率会随之增大,但不利于提高光能利用效率,且气孔开度、叶绿素含量会降低,弱光或强光会影响破坏光系统功能,增加光系统热耗散,不利于光能吸收利用[8-12];光周期对作物光合速率和光能利用效率、色素含量等指标的影响没有形成统一规律,受物种、生育期等因素影响较大[13-15]。研究表明,植物光合对光强的响应趋势基本一致,但在不同的环境和条件下,光响应曲线的参数不尽相同,随着干旱胁迫的增强,温度过高或过低,油桐、刺槐的最大净光合速率(Pmax)及光饱和强度均降低,遮光率过高或全光照,也不利于绣球光合作用[16-18]。表明改变某一环境因子,植物的光合能力和生理特性会受之影响。【本研究切入点】长时间生长在固定的光强和光周期环境中,会影响植物光敏色素的合成,进而影响色素分子对光能的吸收效果[19]。Krol研究显示光周期会影响松树光系统Ⅱ(PSⅡ)的电子传输,其耐受性与光辐射强度和光周期都有关系[20],表明光周期与光强可以通过共同作用影响植物的光合特性。目前针对诸如温度、CO2浓度、水分、光强等单因素对作物光合特性的影响较多[21-23],但对于不同光强和光周期条件下,生菜光合特性变化规律却罕有报道。【拟解决的问题】通过阐明生菜光合参数、叶绿素荧光、叶绿素含量及产量等对光强和光周期的响应规律,旨在为植物工厂光强和光周期调控提供理论依据。
1 材料与方法
1.1 试验环境与材料
试验于2016年11月在西北农林科技大学旱区农业水土工程教育部重点实验室人工光植物工厂内进行。
实验室其他环境因素为:温度为17~23 ℃;二氧化碳浓度为1025~1350 mg/L;湿度为64 %~75 %。供试材料为“香港玻璃脆散叶生菜”,照明设备为荧光灯。营养液选用日本山崎配方营养液(表1)。
1.2 试验设计
试验采用光强×光周期双因素试验,设置了S1、S2、S3、S4四个光强,分别为:150、200、250、300 μmol·m-2·s-1,其设定值为距离灯光源垂直距离25 cm处测量值;3个光周期:D1: 8 h/d(短光周期)、D2: 12 h/d(中长光周期)、D3: 16 h/d(长光周期)。共12个处理:S1D1、S1D2、S1D3、S2D1、S2D2、S2D3、S3D1、S3D2、S3D3、S4D1、S4D2、S4D3,每个处理设3个重复。生菜于2016年11月7日定植,待生菜幼苗长至4叶1心时移栽至14 cm×33.5 cm×24 cm 的水培箱上,定植后缓苗3 d。栽培方式采用深液流法,营养液体积为10 L,每5 d更换1次营养液,每个水培箱种植4株生菜,生菜生长时间为40 d。
1.3 测定及分析方法
于2016年12月8日测定3次,用Li-6400便携式光合测定仪测定生菜光合参数,每处理随机选取3株,测定第5片真叶,测定的光合参数包括:净光合速率(Pn)、胞间二氧化碳浓度(Ci),并计算气孔限制值(Ls=1-Ci/Ca)、叶肉导度(gm=Pn/Ci),其中,Ca为空气中的二氧化碳浓度。
采用捷克PSⅠ公司生产的FluorCam便携式荧光仪测定叶绿素荧光参数。测定叶片与光合一致,生菜暗处理15 min后测定PSII最大光化学效率(Fv/Fm)、光化学猝灭系数(qP)和非光化学猝灭系数(NPQ)。叶绿素荧光测定完成后,立即用同一叶片测定光合色素,采用Arnon修正的方法测定[24]。最后,将选取叶片的植株采摘测定叶鲜重及生物量,并计算光量子利用效率。
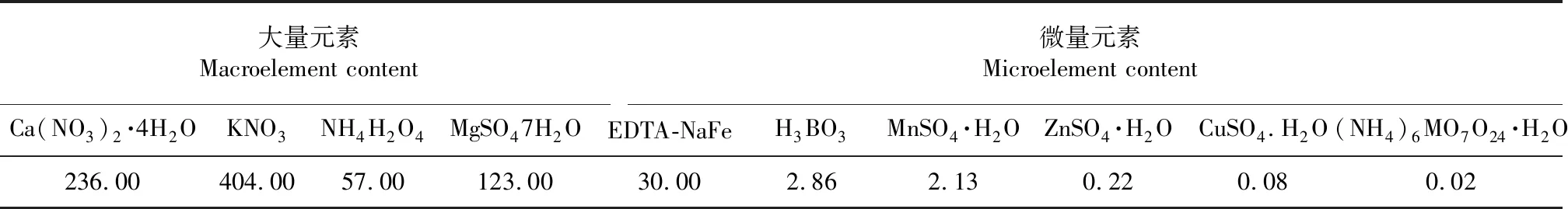
表1 所用营养液中大量元素和微量元素配方
式中:p为光量子利用效率即每mol光量子生产的生菜干重,g/mol;Dw为收获时的干重,g/株;Th为种植密度,为85株/m2;t为种植时间,40 d;DLI为日积累光量子,mol·m-2·d-1;PAR为光强,μmol·m-2·s-1;T为光周期,h/d。
1.4 试验数据处理
采用Excel计算和处理数据,采用Sigmaplot 12.0绘图,差异显著性分析采用SPSS软件。
2 结果与分析
2.1 光强和光周期对生菜色素含量的影响
如图1所示,在相同光强条件下,延长光周期,叶绿素a(Chl a)、叶绿素b(Chl b)、叶绿素(Chl a+b)含量增多,但光周期对Chl a、Chl a+b含量的影响并不显著(P>0.05);相同光周期条件下增大光强,以上色素含量却降低,且光强对Chl a、Chl b和Chl a+b含量有显著性影响(P<0.05),特别的是,150 μmol·m-2·s-1光强下的Chl a、Chl b和Chl a+b含量均显著高于300 μmol·m-2·s-1光强,但200和250 μmol·m-2·s-1与其余2 个光强处理间无显著性差异(P>0.05),光强和光周期对Chl a、Chl b、Chl a+b含量影响交互作用不显著。相同光周期条件下,增加光强有利于提高类胡萝卜素(Caro)含量(300>250>200>150 μmol·m-2·s-1),且Caro含量在300 μmol·m-2·s-1光强时显著大于(22.8 %~28.7 %)150 μmol·m-2·s-1光强的处理,在相同光强条件下,光周期对Caro含量无显著性影响(P>0.05),光强和光周期对Caro含量交互作用显著(P<0.05)。
2.2 光强和光周期对生菜气体交换参数的影响
如图2所示,在相同的光强条件下,延长光周期,水培生菜的净光合速率(Pn)、叶肉导度(gm)、气孔限制值(Ls)均增大。在相同光周期条件下,Pn、gm、Ls均随光强增大而增大,光强和光周期对Pn和gm影响均显著(P<0.05),而对Ls影响不显著(P>0.05),表明增加光强和延长光周期有利于CO2在叶肉中的传输和促进光化学反应。总体来看,光强和光周期对胞间二氧化碳浓度(Ci)的影响不显著;随着光强增大和光周期延长呈下降趋势,且在相同光强条件下,Ci值在16 h/d处理下显著低于8 h/d处理(P<0.05)。一般情况下,若Pn降低,Ci降低,Ls增大,则光合速率降低的原因是 气孔因素;相反,Ci增大,Ls降低,则是非气孔因素,由图可知,生菜在试验条件下Pn值主要受非气孔因素影响,表明Pn主要受其他因素(如酶活性)影响。

不同大写字母表示相同光强下不同光周期之间差异显著(P<0.05),不同小写字母表示相同光周期下不同光强之间差异显著(P<0.05)图1 光强和光周期对色素含量的影响Fig.1 Effect of light intensity and photoperiod on pigment content

不同大写字母表示相同光强下不同光周期之间差异显著(P<0.05),不同小写字母表示相同光周期下不同光强之间差异显著(P<0.05)图2 光强和光周期对叶片气体交换参数的影响Fig.2 Effect of light intensity and photoperiod on leaf gas exchange parameters
2.3 光强和光周期对生菜叶绿素荧光参数的影响
由表2可知,在相同光周期下,增大光强,Fv/Fm均先增大后减小,表明在150~300 μmol·m-2·s-1光强范围内,光强偏强和偏弱对生菜均能产生光抑制。光强相同时,16 h/d下的Fv/Fm大于8、12 h/d,表明中短光照对生菜的胁迫影响要大于长光照。方差分析表明,光强和光周期对Fv/Fm无显著性影响(P>0.05),但交互作用显著(P<0.05)。非光化学荧光猝灭(NPQ)随光强增大和光周期的延长而增大,表明增加光强和延长光周期增加了PSII天线色素吸收光能以热的形式耗散的比例,弱光和短光周期提高了光能利用效率。整体来看,光强、光周期对生菜NPQ无显著影响,交互影响也不显著(P>0.05)。12 和16 h/d光周期下增大光强,光化学猝灭(qP)逐渐减小,在8 h/d时,qP随光强的增加而先增大后减小,光强和光周期交互作用对qP影响显著(P<0.05)。表明在中长光周期下,增大光强降低了PSII天线色素吸收光能用于光化学电子传递的份额,但在低光周期条件下,弱光时增大光强此比例却增大。
2.4 光强和光周期对叶鲜重的影响及光量子利用效率分析
由图3可知,相同光强下,生菜叶鲜重随光周期延长而增大,在8和12 h/d光周期时,光强增大时,叶鲜重也随之增大,但16 h/d光周期时,光强增大,叶鲜重先增大后减小(250 μmol·m-2·s-1时最大)。分析变化趋势,光周期在各光强下对叶鲜重的促进作用均很明显,增加光强对叶鲜重的增效随光周期的增加而减弱,直至到16 h/d时,300 μmol·m-2·s-1光强时叶鲜重反而比减少250 μmol·m-2·s-1减少了10.3 g/株。相同光强下,光周期越大,光量子利用效率(P)越大,随着光强增大,光周期之间的P值差异越来越小,在300 μmol·m-2·s-1时达到近相等水平。在8 h/d光周期时,P随试验光强范围内的光强增大而增大,但增幅很小,12 和16 h/d光周期时,P随光强增大而先增大后减小,均在200 μmol·m-2·s-1时P最大,表明低光强下延长光周期可以提高光量子利用效率,但随着光强增大,光周期的作用逐渐降低。由图4可知,P随日积累光量子先增大后减小,表明在本试验条件下,一定范围内提高的光量子数(≤12.6 mol·m-2·d-1)能提高生产效率,但过高的光能摄入会降低光能利用效率,所以在植物工厂生产中应注意控制总光量子的摄入。

表2 光强和光周期对叶绿素荧光的影响
注:不同大写字母表示相同光强下不同光周期之间差异显著(P<0.05);不同小写字母表示相同光周期下不同光强之间差异显著(P<0.05)。
Note: Different capital letters indicate significant differences among three photoperiod under the same light intensity (P< 0.05); Different small letters indicate significant differences among light intensity of the same photoperiod (P< 0.05).
3 讨 论
在光强过低的情况下,植物合成叶绿素受阻,叶绿素(Chla+b)含量会降低,叶片发黄或泛白,秦玉芝[3]就发现马铃薯在持续弱光时叶片Chla+b含量减少;另一方面,低光强对叶片伤害减小,有利于叶绿素合成。本研究在3个光周期条件下,降低光强Chla、Chlb、Chla+b含量反而升高,说明生菜叶绿素对弱光(100 μmol·m-2·s-1)的耐受性较好,也表明在试验光强范围内(100~300 μmol·m-2·s-1)增加光强,叶绿素的响应可能是基粒减少和破坏[25]。延长光周期有助于提高Chla+b含量,这与杨振超[13]的研究结果一致,有研究表明延长光照时间可以促进叶片色素积累,但会有一个阈值,继续延长会使色素含量降低[5,26],本研究最大光周期为16 h/d,继续延长光周期能否降低Chla+b含量还需要进一步探索。光强对叶绿素含量和净光合速率影响趋势相反,表明在试验光强范围内,原初反应(光合色素吸收、传递光能)的能力不受叶绿素含量的影响。分析结果表明,影响净光合速率降低的因素为非气孔因素。增加光强,Ci降低,这表明增加光强可以显著刺激对气孔二氧化碳的吸收,低光强下对二氧化碳的同化却较低,随着光强增大,二氧化碳在细胞间隙中的富集量减少,CO2进入气孔后,叶肉细胞等阻力受到光强影响[27],虽然gm与Pn的变化趋势一致,但由于Ci的减少,表明叶肉阻力也不是影响Pn的主要因素,所以叶绿体活性与Rubisco活性及RuBP再生能力可能是影响Pn的主要原因,并且姜振升[28]也发现弱光下Pn降低与Rubisco酶活性降低有很大关系,李冬梅[15]发现短日照会促进Rubisco降解和RuBP降低。

图3 光强和光周期对叶鲜重及光量子利用效率的影响Fig.3 Effects of light intensity and photoperiod on leaf fresh weight and utilization efficiency of light quantum
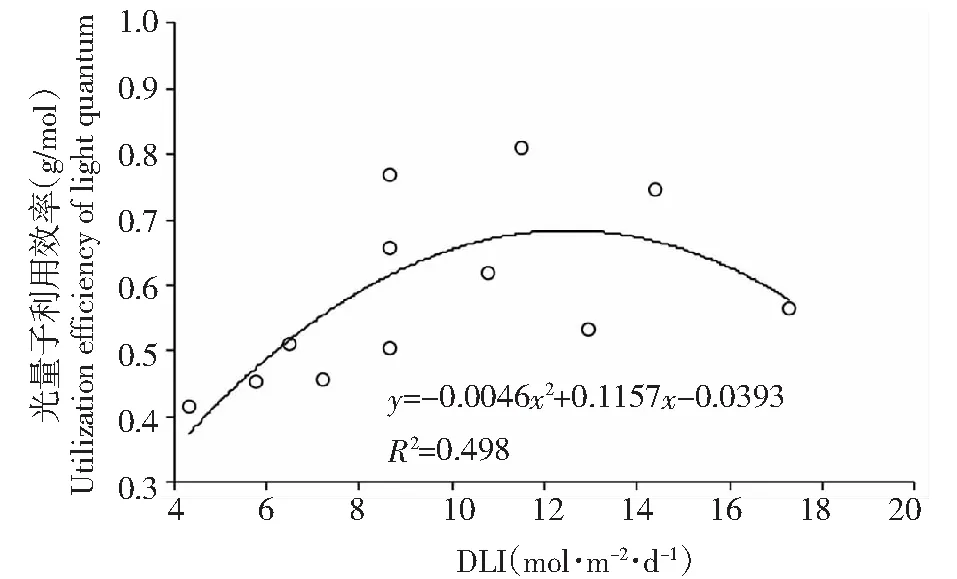
图4 日积累光量子对光量子利用效率(P)的影响Fig.4 Effect of daily light integral on utilization efficiency of light quantum
在3个光周期条件下,随着光照强度的增加,Fv/Fm均先增加后减小,说明强光和弱光对水培生菜生长均有抑制作用,这与Hazrati S[4]的研究结果一致,在不同光环境下,发生的抑制效应不同,无论是强光抑制还是弱光抑制,都是因为PSⅡ系统捕光色素蛋白复合体受到影响,PSⅡ光合中心受到损害[29],一定程度上延长光照时间均能减小这种抑制效应。由光强和光周期对光量子利用效率(P)影响(图4)可知,随光周期延长P增大,表明光周期对生菜干物质量的影响不仅仅是时间的积累,而是可以调控光能利用效率,这与Fv/Fm随光周期的变化趋势相似,进一步说明延长光周期可以提高PSⅡ内电子传递速率和光量子的利用效率。另一方面,在12 和16 h/d时,qP随光强增加而降低,表明低光强可以促使PSⅡ重新氧化的量增大,且有利于增大PSⅡ的开放度,促进电子的传递,最终使产物还原型辅酶NADPH增多[4],从机制上看可能是在光能不足时,光系统的一种补偿[21]。NPQ随光强增大而增大,说明光强增大,生菜将光能用于热耗散的能量也越来越多,天线色素吸收光能用于光合电子传递和同化力( ATP 和 NADPH)的比例减少,但保护了PSⅡ受到光氧化破坏,但降低了PSⅡ对光能的利用效率,延长光周期这种机制会加强。类胡萝卜素的抗氧化性在叶片中的保护功能和黄体素循环有关。光强越大,细胞保护机制越强,导致Caro含量增大,昼夜更替不会产生可以提高叶片光氧化破坏的防御性的环氧化黄体素,而黄体素循环需要类胡萝卜素参与[30]。总体来看,Caro和Chla+b含量随光强的增长而变化趋势相反,前者是保护机制增强,后者是促进作用减弱,说明了水培生菜光合色素对光强较敏感,且能够通过调节含量来适应光强变化。
4 结 论
增加光强有利于水培生菜光化学反应,但减少了叶绿素含量,增加了热耗散且降低了光能利用效率,影响净光合速率降低的因素是非气孔因素。延长光周期不仅可以提高光合速率,并且有利于提高光能利用效率。在植物工厂生产中,生菜的生长光强应在250 μmol·m-2·s-1光强和16 h/d光周期附近时有利于增产和光能利用。
猜你喜欢
潍坊学院学报(2023年6期)2024-01-01 13:25:18
遗传(2023年9期)2023-09-25 09:31:44
农业科技通讯(2023年1期)2023-02-12 07:08:50
中老年保健(2022年6期)2022-08-19 01:44:10
发明与创新(2021年9期)2021-12-06 22:04:23
科学(2020年1期)2020-08-24 08:08:12
农家参谋(2020年5期)2020-06-15 05:12:28
少儿科学周刊·少年版(2017年7期)2017-09-29 08:02:41
少儿科学周刊·儿童版(2017年7期)2017-09-29 02:22:58
食品与健康(2017年3期)2017-03-15 18:07:48
