
食源性致病菌群体感应信号分子的检测
2019-09-28 02:20何伟佳岳思远王翔孙天妹董庆利
生物工程学报 2019年9期
何伟佳,岳思远,王翔,孙天妹,董庆利
食源性致病菌群体感应信号分子的检测
何伟佳,岳思远,王翔,孙天妹,董庆利
上海理工大学 医疗器械与食品学院,上海 200093
群体感应 (Quorum sensing,QS) 在食物中毒导致的食源性疾病暴发机制和食物腐败变质中起主要作用,QS影响致病菌的细胞被膜形成和致病性。文中通过深入了解食源性致病菌的QS信号分子,综述了革兰氏阴性和革兰氏阳性菌产生的信号分子类型,同时介绍了检测QS信号分子的不同技术,并根据QS机制在食品中的影响提出了思考和建议,为监控食源性致病菌提供依据。
食源性致病菌,群体感应,检测技术,信号分子
近年来,人们越来越关注群体感应(Quorum sensing,QS) 信号分子对食品质量与安全的影响。许多研究表明,QS在生物被膜形成、食物中毒导致的食源性疾病暴发机制和食物腐败变质中起主要作用。目前已在牛奶、肉类和蔬菜等不同食品类别中检测到多种信号分子[1],而产生信号分子的嗜冷菌是导致动物源食品腐败变质的主要因素[2]。同时,食源性致病菌大肠杆菌O157:H7、单增李斯特菌及铜绿假单胞菌[3]等易在食品表面和食品接触设备上形成生物被膜,进而导致消费者产生严重的健康问题和经济损失[4]。因此,为加强食品安全,降低食源性疾病的暴发水平和减少消费者的经济损失,需深入研究QS对食源性致病菌细胞个体和细胞间的影响。
实际生活中有很多控制物质可以解决食品设备上生物被膜的形成,如各种抗菌精油[3]及中草药[5]等群体感应抑制剂[6]。自1994年Fuqua等首次提出“群体感应”这一术语,人们对QS系统及其信号分子的研究逐渐增多。QS是指细胞与细胞之间的交流过程,该过程通过产生和感知某些自诱导小分子物质的局部浓度,来进一步调节基因表达产物(如酶或毒性因子) 的产生[7]。自诱导物质也称为信号分子,它的浓度随种群数量的增加而增加,在达到临界阈值后,会引发一种调节反应,进而导致整个细菌群体中与QS相关的目标基因协调表达或抑制[8-9]。研究发现许多致病菌可以释放QS信号分子,它们在食品中发挥着重要的作用,因此通过新型检测技术对QS信号分子进行监测,并预防食源性疾病的发生显得尤为重要。
本文综述了食源性致病菌信号分子的分类及其机制对食品的影响,同时介绍了食源性致病菌信号分子的检测方法,并根据当前QS研究的不足之处,提出了思考和建议,为监控食品环境中致病菌的生长提供了理论依据。
1 信号分子的分类
QS信号分子按化学结构的不同主要分为4个类别:自诱导剂-1 (AI-1)、自诱导剂-2 (AI-2)、自诱导剂-3 (AI-3) 和自诱导多肽(AIP)[10]。表1为部分食源性致病菌的QS信号分子及调控的表现型,除此之外,在肺炎克雷伯菌、志贺氏菌、阴沟肠杆菌等致病菌中也发现AI-3,但它们的信号分子合成途径、结构和调控功能尚不清楚。
食源性致病菌大致分为两类:革兰氏阴性和革兰氏阳性(细胞壁类型是主要区别)。其中AI-1和AI-3主要存在于革兰氏阴性菌中,AIP主要被革兰氏阳性菌使用,这3类QS信号系统主要涉及的是种内交流[19]。而AI-2在革兰氏阴性菌和革兰氏阳性菌中均可发现,其在QS信号系统中主要是用于种间通信[20-21]。革兰氏阳性菌的QS信号分子通常使用的是肽类衍生物,而革兰氏阴性菌的QS信号分子一般使用脂肪酸衍生物,最常见的是AI-1类的N-酰基-髙丝氨酸内酯(N-acyl L-homoserine lactones,AHLs)[22]。自从在研究费氏弧菌中生物发光现象时发现第一个基于AHL的QS系统以来,到目前为止,已在50多种革兰氏阴性菌中发现了含AHLs的QS系统。自诱导多肽(AIPs)则在40多个物种中已得到确认,AIPs具有体积小、稳定性高、特异性和多样性等特点[23]。
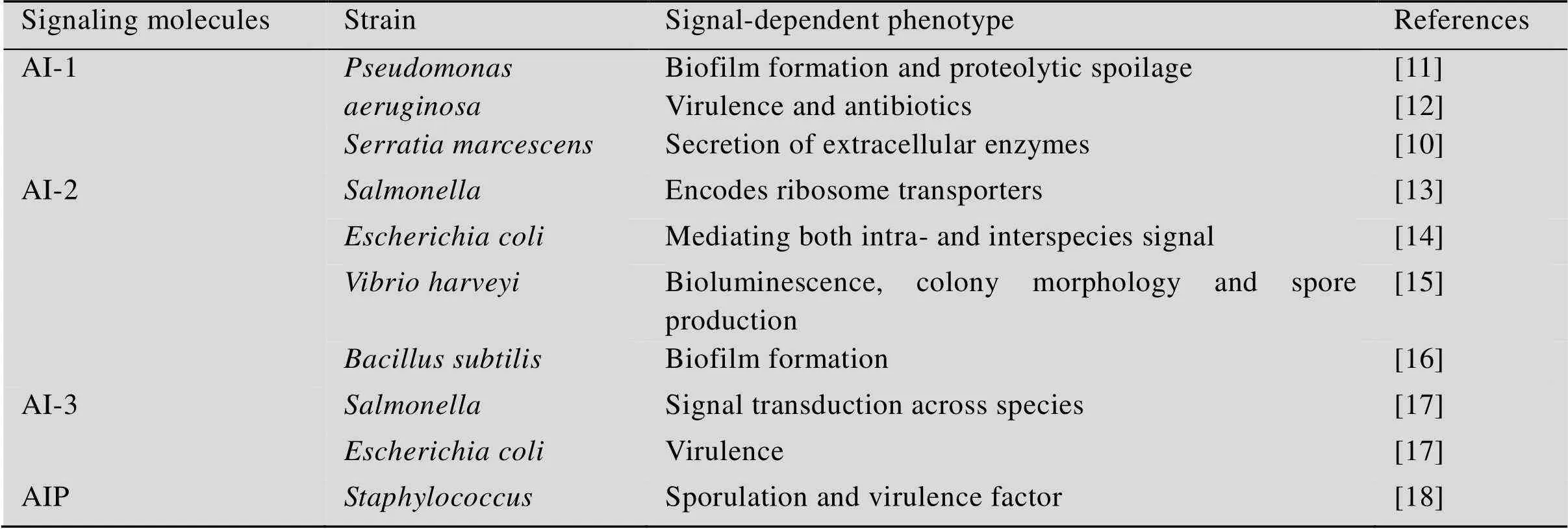
表1 部分食源性致病菌的QS信号分子及调控的表现型
对于食源性致病菌而言,QS作为一种细胞间的传播机制,它通常发生在相同或不同种类的细菌之间。当信号分子达到阈值时就会激活一些基因的转录,从而改变生物表型[11]。因此,依靠QS改变的基因大多发生在细菌生长指数期后半段或稳定期,而不是在生长初期被激活或抑制[24]。QS调控性状包括生物被膜的形成、毒力因子的产生、生物发光、耐酸胁迫、细菌素生产、粘附能力、形态转换和定向生长[23]。图1简单地列举了群体感应与信号分子的关系。
2 QS信号分子的检测
根据分子结构,QS信号分子通常可以通过生物测定或化学方法检测。
2.1 生物测定
生物传感技术是一种选择性和敏感性较高的技术,在该技术中,分析物被结合到生物成分(如组织、酶、蛋白质等) 中,使信号被转换成可分析和可测量的输出量[25]。通常使用的生物传感器自身不产生信号分子,但拥有它们的同源受体,其检测是通过不同的表型,包括色素、光(生物发光)、绿色荧光蛋白和β-半乳糖苷酶来实现的。生物传感技术因其灵敏度高和可检测多种AHL型系统/自诱导分子而得到广泛应用[26]。
使用生物传感器也有一定缺陷。菌株产生的AHLs浓度通常较低且低于大多数生物传感器的检测极限,同时,生物传感器检测信号分子生产的能力受培养基组分、食品组分及食品中添加剂的影响,因此,尽管AHLs分子存在,生物传感器仍可能会产生假阴性结果[27-28]。虽然每个生物传感器适用于AHLs化合物的范围有限,但生物传感器已被应用于一些食品中QS信号分子的检测。Blana等[29]和Almasoud等[30]为测试精油、乳酸和苹果酸类抗菌物质对细菌的信号分子活性的影响,使用生物传感技术筛选菌株并测试微生物培养液中AI-2活性,从而证明植物精油和有机酸可有效抑制细菌的QS能力。在不久的将来,生物传感器有可能发展成为仪器,为更精确和灵敏的测定提供方向。
2.2 化学测定
2.2.1 薄层色谱法(Thin layer chromatography,TLC)

图1 群体感应调控图[11]
TLC是跟踪有机化学反应进展、分析植物化学和生物技术中有机化合物纯度的最有效工具之一。与所有的色谱方法一样,它利用分析物与流动相和固定相的不同亲和力,实现了有机分子复杂混合物的分离[31]。在QS检测中,一般是当细菌生长到指数后期或晚期时,用二氯甲烷、乙酸乙酯或氯仿等有机溶剂提取培养上清液中的AHLs,然后通过TLC在C18反相层析板上分离细菌提取物,最后覆盖一层含传感菌的琼脂培养基后进行培养[32-33]。
TLC是文献中普遍报道的一种技术,可用于所有信号分子的检测和表征[26]。为判断信号分子对细菌动态生长行为的影响,Chorianopoulos等[34]和Dourou等[35]通过TLC分析发现,部分细菌(例如哈夫尼菌属、耶尔森菌属等) 的AHLs会抑制其他细菌的代谢活性和生物被膜的生长。TLC虽操作繁琐,但因不需要昂贵的硬件设备,且其结合生物传感技术分析AHLs较为敏感可靠,现已作为一种有效分析方法并被广泛采用。
2.2.2 高效液相色谱-质谱(High-performance liquid chromatography-MS,HPLC-MS)
HPLC-MS集成了高效液相色谱和质谱扫描,可以在检测产AHLs离子质量的同时,在单个质谱图中以高分辨率和高质量精度生成全质谱-色谱运行,该方法可筛选多种革兰氏阴性菌中的AHLs产生,如铜绿假单胞菌、洋葱伯克霍尔德菌、欧文氏菌等[36]。高效液相色谱(HPLC) 被证明是一种有效的生物液体、中药物和代谢产物的定量测定技术。对于AIPs,尤其是与食品相关的细菌素,检测方法主要是通过琼脂平板做抑制试验,基于HPLC技术测定指示菌株的抑制圈以及测量指示菌株的透明区域[37]。
綦国红等[38]证明,在没有标样的情况下,样品经过处理后可应用HPLC-MS对样品中任意的AHLs分子进行定性分析。HPLC-MS不仅对检测的AHLs化合物分离能力强,而且对AHLs的分析范围较广。随着技术的进步,为满足信号分子的定性及定量分析,HPLC-MS已逐渐被HPLC- MS/MS替代并被广泛使用。
2.2.3 气相色谱-质谱(Gas chromatography- MS,GC-MS)
因为可检测到酰基链的变化,GC-MS已被证明是定性和定量测定AHL的一种简便方法。GC-MS可直接、有效地反映细菌培养液上清中存在的信号分子[39]。GC-MS适用于沸点低于400 ℃、挥发性强、极性小的化合物[26]。
Cataldi等[40]利用GC-MS明确地鉴定了沙门氏菌、铜绿假单胞菌、结肠炎耶尔森菌和荧光假单胞菌提取物中的信号分子,并在沙门氏菌无细胞培养上清液的提取物中,发现了几种新型的AHLs。Davenport等[41]通过GC-MS对铜绿假单胞菌的代谢产物进行分析,显示约三分之一的代谢产物(包括三羧酸循环中间体、氨基酸和脂肪酸的浓度变化) 均受QS的干扰。一般采用GC-MS对低分子量化合物进行优选分析,用HPLC-MS对体积较大、极性较大的化合物进行分析。
除了上述方法,还可通过串联质谱法(Tandem mass spectrometry,MS/MS)、核磁共振光谱(Nuclear magnetic resonance spectroscopy,NMR)、红外光谱法(Infrared spectroscopy) 等分析方法检测和定量AHLs[10]。质谱、气相色谱和液相色谱的结合为精确地测定分析物提供了有力的工具。根据不同的分子类型选择相应的检测方法,有助于食品中信号分子的定量检测,因此应提高QS信号分子检测的灵敏度,保障食品安全。
3 群体感应机制对食品的潜在影响
一个生物体可拥有多个具有不同功能的QS系统,不同QS系统之间相互合作,共同调节生物体表型[42]。以铜绿假单胞菌为例,它有3个经典的QS系统,分别为Las、Rhl和PQS系统。Las系统转录调节因子和合成蛋白酶;Rhl系统产生合成酶并控制鼠李糖脂的产生,以及参与铜绿假单胞菌毒素蛋白向宿主细胞细胞质的释放;而假单胞菌PQS系统则与一些基因的产生和信号分子经外膜囊泡的转运有关[43]。此外,假单胞菌PQS系统还诱导鼠李糖脂和其他分子参与铜绿假单胞菌中生物被膜的形成[44]。各系统相互合作且分工明确,调节细胞内部和细胞之间的信息交流,帮助细胞更快适应周围环境并接收外界信息。铜绿假单胞菌的QS通路图见图2。
由QS机制可以看出,细菌在信号分子的调控下,可改变自身的代谢速率、表现型以及酶的产生等,从而对食品造成潜在的影响。研究发现,牛奶的变质主要与蛋白水解性嗜冷菌有关[45-46],Pinto等[1]从生乳中分离到的几种蛋白水解性嗜冷菌(铜绿假单胞菌、沙雷氏菌、大肠杆菌、哈氏弧菌) 均产生不同的AHLs,结果表明,QS可能在牛奶和乳制品的变质过程中发挥重要作用。Bruhn等[47]通过TLC检测发现,肉及肉制品在储存和变质过程中一直存在AHLs,其中,肉类样品中产AHLs的细菌主要为肠杆菌科(占91%),其次是假单胞菌。Dogan等[48]、Ammor等[49]和Skandamis等[50]等研究证实与食物变质相关的几种细菌降解酶的活性均受QS调节,包括蛋白水解、脂质水解、几丁质水解和果胶水解,这表明这种细胞间通讯在食物变质中可能发挥作用。
QS不仅参与许多食物的腐败和发酵[23],还参与细菌的致病性,增加食品的致病机率。Kariminik等[12]发现铜绿假单胞菌作为一种条件致病菌,可以利用QS来调控其基因表达,影响宿主的免疫应答反应。Silagyi等[51]确定大肠杆菌O157:H7在QS的影响下,会促进细菌的生物被膜形成和粘附性,从而增加致病菌与食品接触表面交叉污染的机率。种种迹象都表明QS在食品变质过程和细菌致病性中发挥了重要作用。

图2 铜绿假单胞菌的QS通路图[12]
4 结论与展望
越来越多的报道证明QS不仅与食品的腐败变质有关,还与食源性致病菌的毒性和致病机制有关。虽然在变质的食品中可检测到一定量的信号分子,但食品基质对信号分子的影响仍不清楚。例如,它们如何影响其他种类的细菌,它们对腐败变质的影响(如果有的话) 或食品成分如何影响QS分子(例如食品基质中的抑制剂) 的释放和稳定性。因此,还需要进一步研究,以了解信号分子在食品基质中的潜在作用。为了解复杂的微生物群落中相互交流的信息,深入研究QS系统,以积极改善食品质量和保障食品安全,提出以下几点建议:1) 确定食品环境中细菌相互作用的机制以及食品成分对信号分子的释放和稳定性的影响。2) 更新信号分子检测技术,根据致病毒素或食品中信号分子的浓度提高QS检测灵敏度。3) 明确能改变细胞表型的信号分子阈值,对信号分子定量建模,提前预防食品变质和安全问题。4) 开发QS抑制剂,以减小QS对食品的负面影响。
[1] Pinto UM, de Souza Viana E, Martins ML, et al. Detection of acylated homoserine lactones in gram-negative proteolytic psychrotrophic bacteria isolated from cooled raw milk. Food Control, 2007, 18(10): 1322–1327.
[2] de Oliveira GB, Favarin L, Luchese RH, et al. Psychrotrophic bacteria in milk: how much do we really know? Braz J Microbiol, 2015, 46(2): 313–321.
[3] Tapia-Rodriguez MR, Hernandez-Mendoza A, Gonzalez-Aguilar GA, et al. Carvacrol as potentialinhibitor ofand biofilm production on stainless steel surfaces. Food Control, 2017, 75: 255–261.
[4] de Kievit TR, Iglewski BH. Bacterial quorum sensing in pathogenic relationships. Infect Immun, 2000, 68(9): 4839–4849.
[5] Li XP, Chen GF, Wang JX, et al. Synergistic effect of cortex mouton extract combined with tea polyphenols on quality of cold-stored turbot fillets based on the inhibition of quorum sensing. Food Sci, 2017, 38(19): 223–229 (in Chinese).李学鹏, 陈桂芳, 王金厢, 等. 基于群体感应抑制的丹皮提取物和茶多酚对冷藏大菱鲆的协同保鲜作用. 食品科学, 2017, 38(19): 223–229.
[6] Ellappan K, Thirumalaswamy K, Harish BN, et al. Inhibition of quorum sensing-controlled biofilm formation inby quorum-sensing inhibitors. Microb Pathog, 2017, 111: 99–107.
[7] Fuqua WC, Winans SC, Greenberg EP. Quorum sensing in bacteria: the LuxR-LuxI family of cell density-responsive transcriptional regulators. J Bacteriol, 1994, 176(2): 269–275.
[8] Bassler BL. Small talk: cell-to-cell communication in bacteria. Cell, 2002, 109(4): 421–424.
[9] Williams P. Quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology, 2007, 153(12): 3923–3938.
[10] Bai AJ, Rai VR. Bacterial quorum sensing and food industry. Compr Rev Food Sci Food Saf, 2011, 10(3): 183–193.
[11] Pérez-Velázquez J, Gölgeli M, García-Contreras R. Mathematical modelling of bacterial quorum sensing: a review. Bull Math Biol, 2016, 78(8): 1585–1639.
[12] Kariminik A, Baseri-Salehi M, Kheirkhah B.quorum sensing modulates immune responses: an updated review article. Immunol Lett, 2017, 190: 1–6.
[13] Taga ME, Semmelhack JL, Bassler BL. The LuxS-dependent autoinducer AI-2 controls the expression of an ABC transporter that functions in AI-2 uptake in. Mol Microbiol, 2001, 42(3): 777–793.
[14] Brito PH, Rocha EPC, Xavier KB, et al. Natural genome diversity of AI-2 quorum sensing in: conserved signal production but labile signal reception. Genome Biol Evol, 2013, 5(1): 16–30.
[15] Bassler BL, Greenberg EP, Stevens AM. Cross-species induction of luminescence in the quorum-sensing bacterium. J Bacteriol, 1997, 179(12): 4043–4045.
[16] DuanisAssaf D, Steinberg D, Chai YR, et al. The LuxS based quorum sensing governs lactose induced biofilm formation by. Front Microbiol, 2016, 6: 1517.
[17] Walters M, Sircili MP, Sperandio V. AI-3 synthesis is not dependent onin. J Bacteriol, 2006, 188(16): 5668–5681.
[18] Pollitt EJG, West SA, Crusz SA, et al. Cooperation, quorum sensing, and evolution of virulence in. Infect Immun, 2014, 82(3): 1045–1051.
[19] Smith JL, Fratamico PM, Novak JS. Quorum sensing: a primer for food microbiologists. J Food Prot, 2004, 67(5): 1053–1070.
[20] Schauder S, Shokat K, Surette MG, et al. The LuxS family of bacterial autoinducers: biosynthesis of a novel quorum-sensing signal molecule. Mol Microbiol, 2001, 41(2): 463–476.
[21] de Keersmaecker SCJ, Sonck K, Vanderleyden J. Let LuxS speak up in AI-2 signaling. Trends Microbiol, 2006, 14(3): 114–119.
[22] Ahumedo M, Drosos JC, Vivas-Reyes R. Application of molecular docking and ONIOM methods for the description of interactions between anti-quorum sensing active (AHL) analogues and thebinding site. Mol BioSyst, 2014, 10(5): 1162–1171.
[23] Johansen P, Jespersen L. Impact of quorum sensing on the quality of fermented foods. Curr Opin Food Sci, 2017, 13: 16–25.
[24] Goo E, An JH, Kang YS, et al. Control of bacterial metabolism by quorum sensing. Trends Microbiol, 2015, 23(9): 567–576.
[25] Rodriguez-Mozaz S, de Alda MJL, Barceló D. Biosensors as useful tools for environmental analysis and monitoring. Anal Bioanal Chem, 2006, 386(4): 1025–1041.
[26] Turan NB, Chormey DS, Büyükpınar Ç, et al. Quorum sensing: little talks for an effective bacterial coordination. TrAC Trends Anal Chem, 2017, 91: 1–11.
[27] Steindler L, Venturi V. Detection of quorum-sensing-acyl homoserine lactone signal molecules by bacterial biosensors. FEMS Microbiol Lett, 2007, 266(1): 1–9.
[28] Zhao J, Quan CS, Jin LM, et al. Production, detection and application perspectives of quorum sensing autoinducer-2 in bacteria. J Biotechnol, 2018, 268: 53–60.
[29] Blana VA, Nychas GJE. Presence of quorum sensing signal molecules in minced beef stored under various temperature and packaging conditions. Int J Food Microbiol, 2014, 173: 1–8.
[30] Almasoud A, Hettiarachchy N, Rayaprolu S, et al. Inhibitory effects of lactic and malic organic acids on autoinducer type 2 (AI-2) quorum sensing ofO157:H7 andTyphimurium. LWT-Food Sci Technol, 2016, 66: 560–564.
[31] Kumar S, Jyotirmayee K, Sarangi M. Thin layer chromatography: a tool of biotechnology for isolation of bioactive compounds from medicinal plants. Int J Pharm Sci Rev Res, 2013, 18(1): 126–132.
[32] Blana V, Georgomanou A, Giaouris E. Assessing biofilm formation byserovar Typhimurium on abiotic substrata in the presence of quorum sensing signals produced by. Food Control, 2017, 80: 83–91.
[33] Zhang QQ. Study on quorum-sensing signal of spoilage bacteria in raw chicken meat[D]. Nanjing: Nanjing Agricultural University, 2014: 1–131 (in Chinese).张秋勤. 生鲜鸡肉中腐败菌群体感应信号分子研究[D]. 南京: 南京农业大学博士论文, 2014: 1–131.
[34] Chorianopoulos NG, Giaouris ED, Kourkoutas Y, et al. Inhibition of the early stage ofserovar enteritidis biofilm development on stainless steel by cell-free supernatant of aculture. Appl Environ Microbiol, 2010, 76(6): 2018–2022.
[35] Dourou D, Ammor MS, Skandamis PN, et al. Growth ofenteritidis andtyphimurium in the presence of quorum sensing signalling compounds produced by spoilage and pathogenic bacteria. Food Microbiol, 2011, 28(5): 1011–1018.
[36] Patel NM, Moore JD, Blackwell HE, et al. Identification of unanticipated and novel-Acyl L-homoserine lactones (AHLs) using a sensitive non-targeted LC-MS/MS method. PLoS ONE, 2016, 11(10): e0163469.
[37] Ge JP, Fang BZ, Wang Y, et al.enhances production of Paracin1.7, a bacteriocin produced byHD1-7, isolated from Chinese fermented cabbage. Ann Microbiol, 2014, 64(4): 1735–1743.
[38] Qi GH, Dong MS, Wu SM, et al. Determination of signal molecules N-Acyl-homoserine lactones by HPLC-MS. J Instrum Anal, 2007, 26(3): 417–419 (in Chinese).綦国红, 董明盛, 吴胜明, 等. HPLC-MS法检测N-酰基-高丝氨酸内酯类信号分子. 分析测试学报, 2007, 26(3): 417–419.
[39] Cataldi TRI, Bianco G, Frommberger M, et al. Direct analysis of selected-acyl-L-homoserine lactones by gas chromatography/mass spectrometry. Rapid Commun Mass Spectrom, 2004, 18(12): 1341–1344.
[40] Cataldi TRI, Bianco G, Palazzo L, et al. Occurrence of-acyl-L-homoserine lactones in extracts of some Gram-negative bacteria evaluated by gas chromatography-mass spectrometry. Anal Biochem, 2007, 361(2): 226–235.
[41] Davenport PW, Griffin JL, Welch M, et al. Quorum sensing is accompanied by global metabolic changes in the opportunistic human pathogen. J Bacteriol, 2015, 197(12): 2072–2082.
[42] Williams P, Cámara M. Quorum sensing and environmental adaptation in: a tale of regulatory networks and multifunctional signal molecules. Curr Opin Microbiol, 2009, 12(2): 182–191.
[43] Wade DS, Calfee MW, Rocha ER, et al. Regulation ofquinolone signal synthesis in. J Bacteriol, 2005, 187(13): 4372–4380.
[44] Mellbye B, Schuster M. Physiological framework for the regulation of quorum sensing-dependent public goods in. J Bacteriol, 2014, 196(6): 1155–1164.
[45] Gargouri A, Hamed H, ElFeki A. Analysis of raw milk quality at reception and during cold storage: Combined effects of somatic cell counts and psychrotrophic bacteria on lipolysis. J Food Sci, 2013, 78(9): 1405–1411.
[46] Ribeiro Júnior JC, de Oliveira AM, Silva FDG, et al. The main spoilage-related psychrotrophic bacteria in refrigerated raw milk. J Dairy Sci, 2018, 101(1): 75–83.
[47] Bruhn JB, Christensen AB, Flodgaard LR, et al. Presence of acylated homoserine lactones (AHLs) and AHL-producing bacteria in meat and potential role of AHL in spoilage of meat. Appl Environ Microbiol, 2004, 70(7): 4293–4302.
[48] Dogan B, Boor KJ. Genetic diversity and spoilage potentials amongspp.isolated from fluid milk products and dairy processing plants. Appl Environ Microbiol, 2003, 69(1): 130–138.
[49] Ammor MS, Michaelidis C, Nychas GJE. Insights into the role of quorum sensing in food spoilage. J Food Prot, 2008, 71(7): 1510–1525.
[50] Skandamis PN, Nychas GJE. Quorum sensing in the context of food microbiology. Appl Environ Microbiol, 2012, 78(16): 5473–5482.
[51] Silagyi K, Kim SH, Lo YM, et al. Production of biofilm and quorum sensing byO157:H7 and its transfer from contact surfaces to meat, poultry, ready-to-eat deli, and produce products. Food Microbiol, 2009, 26(5): 514–519.
Progress in detection and modeling of quorum sensing molecules of foodborne pathogens
Weijia He, Siyuan Yue, Xiang Wang, Tianmei Sun, and Qingli Dong
School of Medical Instrument and Food Engineering, University of Shanghai for Science and Technology, Shanghai 200093, China
Quorum sensing (QS) plays a major role in the outbreak mechanism of foodborne diseases caused by food poisoning and food spoilage. QS affects the formation of cell membrane and pathogenicity ofpathogenic bacteria. Through the in-depth understanding of QS molecules of food-borne pathogens, we describe here the types of signal molecules produced by Gram-negative and Gram-positive bacteria, and the differences in QS molecules. Meanwhile, we introduce the detection of QS molecules by different technologies. According to the influence of QS on food, we propose also future research needs for the control of foodborne pathogenic bacteria.
foodborne pathogen, quorum sensing, detecting techniques, signal molecules
February 11, 2019;
May 14, 2019
National Key Research and Development Project "Research and Development of Key Technologies for Food Safety" (No. 2018YFC1602502).
Qingli Dong. Tel: +86-21-55271117; E-mail: dongqingli@126.com
国家重点研发计划“食品安全关键技术研发”重点专项(No. 2018YFC1602502)资助。
2019-05-29
http://kns.cnki.net/kcms/detail/11.1998.q.20190529.1338.001.html
何伟佳, 岳思远, 王翔, 等. 食源性致病菌群体感应信号分子的检测. 生物工程学报, 2019, 35(9): 1707–1714.
He WJ, Yue SY, Wang X, et al. Progress in detection and modeling of quorum sensing molecules of foodborne pathogens. Chin J Biotech, 2019, 35(9): 1707–1714.
(本文责编 陈宏宇)
猜你喜欢
中老年保健(2022年1期)2022-08-17
食品安全导刊(2021年21期)2021-08-30
食品安全导刊(2021年20期)2021-08-30
中老年保健(2021年6期)2021-08-24
中国现代医药杂志(2020年10期)2020-12-14
天然产物研究与开发(2019年1期)2019-03-01
中成药(2017年9期)2017-12-19
中国医药指南(2017年3期)2017-11-13
上海农业学报(2017年4期)2017-04-10
中华老年口腔医学杂志(2016年5期)2016-03-01
