
紫花含笑适宜生境的保护空缺与人类干扰分析
2019-09-26 02:36刘慧明高吉喜宋创业于胜祥
中国环境科学 2019年9期
刘慧明,高吉喜*,宋创业,于胜祥
紫花含笑适宜生境的保护空缺与人类干扰分析
刘慧明1,高吉喜1*,宋创业2,于胜祥2
(1.生态环境部卫星环境应用中心,北京 100094;2.中国科学院植物研究所,北京 100093)
以紫花含笑地理分布点数据为基础,结合地形和气候数据,使用最大熵(MaxEnt)模型,对紫花含笑的生境适宜度进行了评估,并基于阈值法,将紫花含笑生境分为适宜区和最适宜区,分析了紫花含笑生境适宜区和最适宜区在我国的地理分布、保护现状以及人类干扰状况,结果表明:年均降雨量对紫花含笑的分布影响最为重要,地形因子和温度因子对紫花含笑的分布影响相对较小;紫花含笑的生境最适宜区面积为13.7万km2,适宜区为27.0万km2,分布在22°~30°N, 107°~120°E之间.广东、福建、广西、江西和湖南等5个省是紫花含笑生境最适宜分布区面积最广的地区;位于国家级自然保护区内的生境最适宜区和适宜区面积约占总面积的1.9 %,大部分适宜生境未得到有效保护;紫花含笑生境适宜区和最适宜区内,林地是最大的土地利用类型,但是各项人类活动也占有相当大的比例.
生境适宜度;最大熵模型;ROC曲线;土地利用;自然保护区;人类干扰
紫花含笑是木兰科含笑属中极其少有的开红花的树种,为木兰科的珍品[1],但紫花含笑天然种群正逐渐减少[2].在“中国生物多样性红色名录”中[3],紫花含笑被认定为“濒危”植物,亟需保护.然而,目前关于紫花含笑生境适宜度及潜在生境分布、保护现状等未见报道,这会影响对紫花含笑开展有效的原地和迁地保护.
物种潜在生境的了解及其适宜生境的空间分布预测是进行物种保护和资源开发利用策略的基础[4-5],对开展受胁物种的生境恢复和有效保护具有重要的指导意义.物种分布模型是以生态位理论为基础,建立物种分布与环境因子之间的关系,并对其潜在分布或者生境进行预测[6]的一种方法.常用的物种分布模型有生物气候包络模型[7]、生态位因子分析[8]、分类与回归树[9]、最大熵[10]、基于规则集的遗传算法[11]等.最大熵模型以最大熵理论为基础,将物种与环境因子视为一个系统,计算系统具有最大熵时的状态参数确定物种和环境之间的稳定关系,以此估计物种的地理分布[10].与其他模型相比,最大熵模型具有较强的拟合与预测能力[10],可以在建模样本量较少的情况下获得较好的预测结果[12-14].因此,本研究以中国数字植物标本馆收集的紫花含笑标本地理分布数据为基础,结合地形、温度和降雨等环境因子数据,采用MaxEnt模型对紫花含笑的生境适宜度进行模拟和评估,了解紫花含笑栖息地的立地条件,并结合自然保护区分布数据和土地利用数据,评价紫花含笑的保护空缺和适宜生境的人类干扰状况,为该物种的有效保护和合理利用提供参考.
1 数据和方法
1.1 紫花含笑地理分布数据
紫花含笑地理分布数据来源于中国数字标本馆(http://www.cvh.ac.cn/),共计192条记录,记录信息包括物种名、经度和纬度,地理空间分布见图1.
1.2 环境变量数据
从世界气候数据库(WorldClim v2.0, http://www.worldclim.org/)中,提取研究区的19个气候指标数据(表1).该数据库利用全球各地气象站记录的气象信息,通过整合插值生成全球气候栅格数据(1950~2000年平均值),空间分辨率为2.5¢(约4km)[15].地形数据来源于数字高程模型,空间分辨率2.5¢.在ArcGIS中利用Spatial Analyst Tools工具中Surface模块的Slope和Aspect算法提取坡度和坡向数据,其中坡向设定以正北为0°,往南逐渐增大,正南为180°.为了降低环境变量之间的相关性对模拟结果的影响,本研究采用Pearson相关分析计算了各个环境变量之间的相关系数(表1).以0.7为阈值[16],2个变量之间的相关系数高于0.7的,取其中一个变量参加模型运算.最终选择海拔、坡向、坡度、bio2、bio4、bio6、bio8、bio12、bio15和bio18等10个变量参与模型计算.
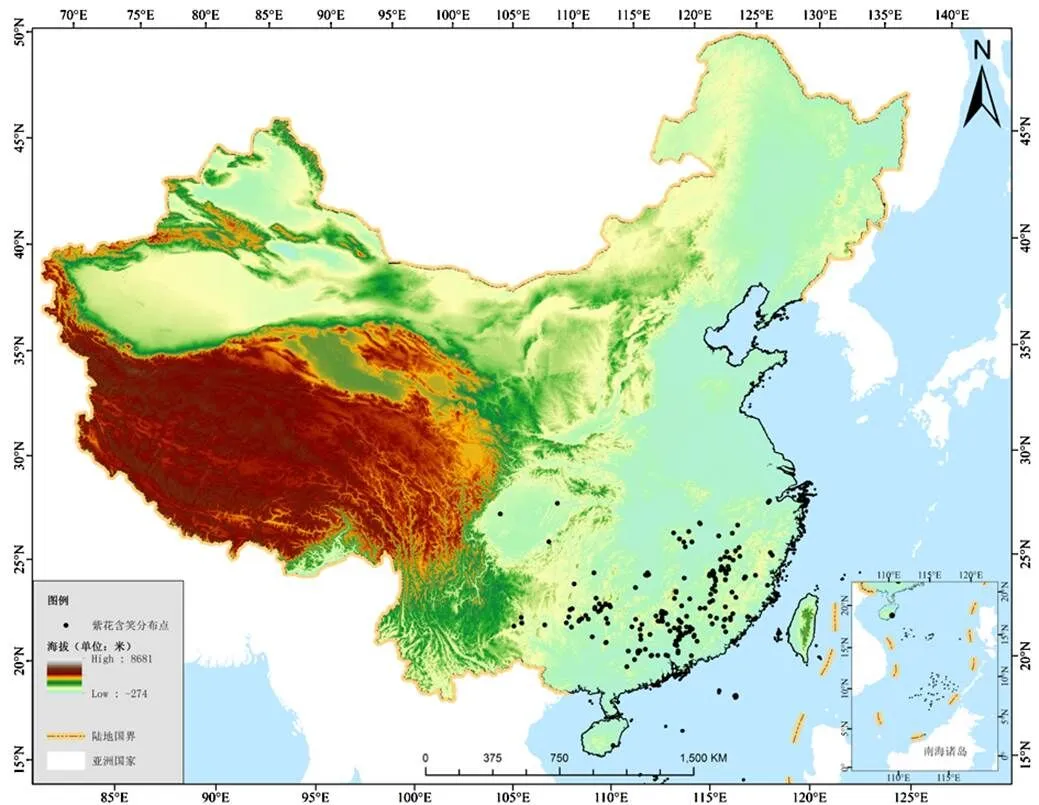
图1 紫花含笑样点地理空间分布

表1 环境变量之间的相关系数
1.3 自然保护区空间分布数据和土地利用数据
自然保护区空间分布数据由生态环境部南京环境科学研究所提供,2015年土地利用数据来源于中国科学院资源环境数据中心(http://www.resdc. cn/),空间分辨率为1km.
1.4 物种分布模型
MaxEnt模型是以最大熵理论为基础的物种分布预测模型[10],其在物种生境模拟方面表现出了优越的性能,广泛应用于物种潜在分布和生境适宜性评价[17-18].在计算中,随机选取25%的紫花含笑地理分布点作为测试集,剩余75%的分布点作为训练集.其他参数设置均为软件默认值.模型输出格式为ASCII栅格图层,图层中每个栅格的值代表紫花含笑在该区域对环境的适应情况,值域为0~1.在ArcGIS软件中加载MaxEnt的运算结果,进行生境等级划分和可视化表达.
在Maxent模型中,计算了环境因子对模型的贡献和重要性,同时采用刀切法判断环境因子的重要性,具体计算方法请参见文献[10,19].另外,绘制生境适宜度对重要环境因子的响应曲线[20].
1.5 模型精度验证
采用受试者工作特征曲线(ROC)进行模型精度检验.ROC曲线以真阳性率为纵坐标(敏感性,实际存在且被预测为存在的比率),以假阳性率(1-特异性,实际不存在但被预测为存在的比率)为横坐标,AUC值指ROC曲线与横坐标围成的面积值,值域为0~1,AUC值越大表示与随机分布相距越远,环境变量与预测的物种地理分布之间的相关性越大,即模型预测效果越好,反之说明模型预测效果越差.AUC值在0.5~0.6,0.6~0.7,0.7~0.8,0.8~0.9,0.9~1分别表示模拟效果失败、较差、一般、好、非常好[21].
1.6 生境适宜度阈值选择
MaxEnt模型给出的结果是栅格数据,值域范围为0~1.为了获取明确的紫花含笑适宜生境,需要选择阈值将连续性的概率数据转换为二元数据.基于MaxEnt的阈值选择方法较多,如0.5[22],10%训练存在逻辑阈值[23];ROC曲线敏感度为0.95时的阈值[24]; 0.8[25],20%训练存在逻辑阈值[26].
2 研究结果
2.1 模型精度验证和生境适宜度阈值选择
本研究中,训练数据和检验数据的AUC值均在0.9以上,模拟效果非常好(图2).
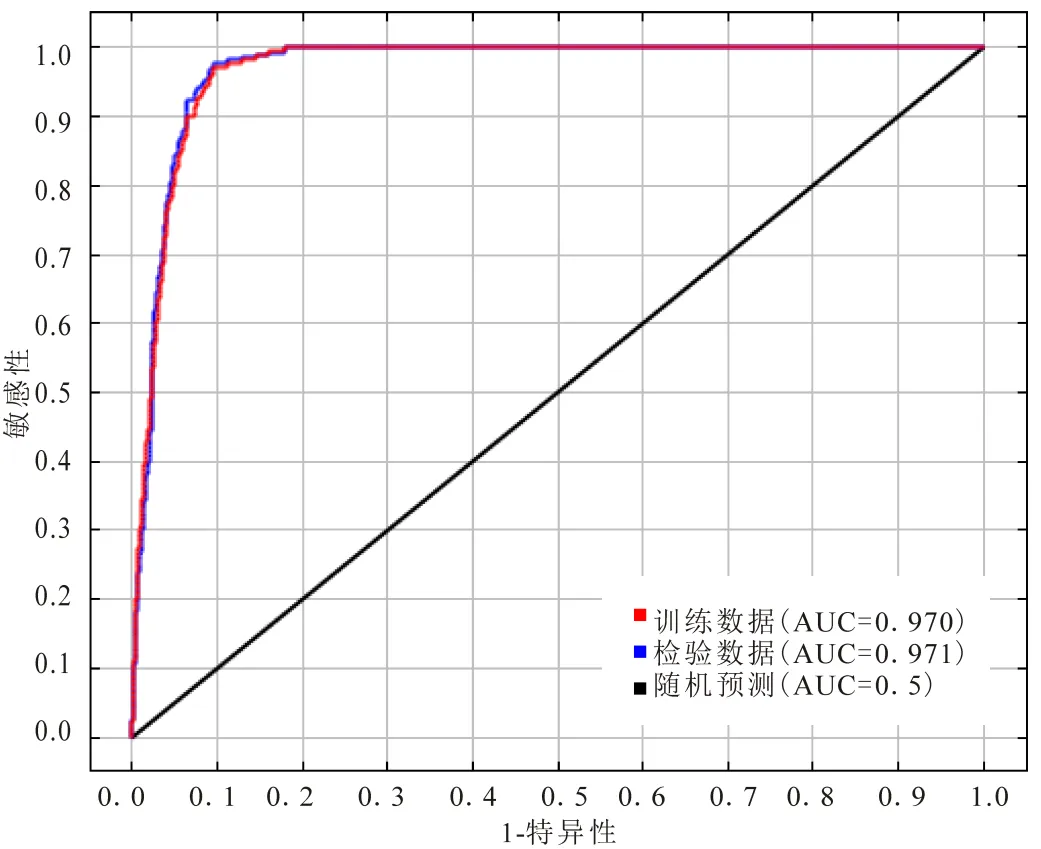
图2 MaxEnt模型的ROC验证曲线
本研究分别计算了“10%训练存在逻辑阈值”、“20%训练存在逻辑阈值”和“ROC曲线敏感度为0.95”三种方法所确定的阈值,分别为0.31、0.43和0.22.基于上述计算结果以及保护成本的考虑,本研究选择相对较为保守的阈值;以0.8作为最适宜生境的阈值,大于0.8为紫花含笑最适宜生境;0.5~0.8之间为适宜生境;0.5以下为不适宜生境,适宜区空间分布见图7.
2.2 环境因子的贡献及生境适宜度
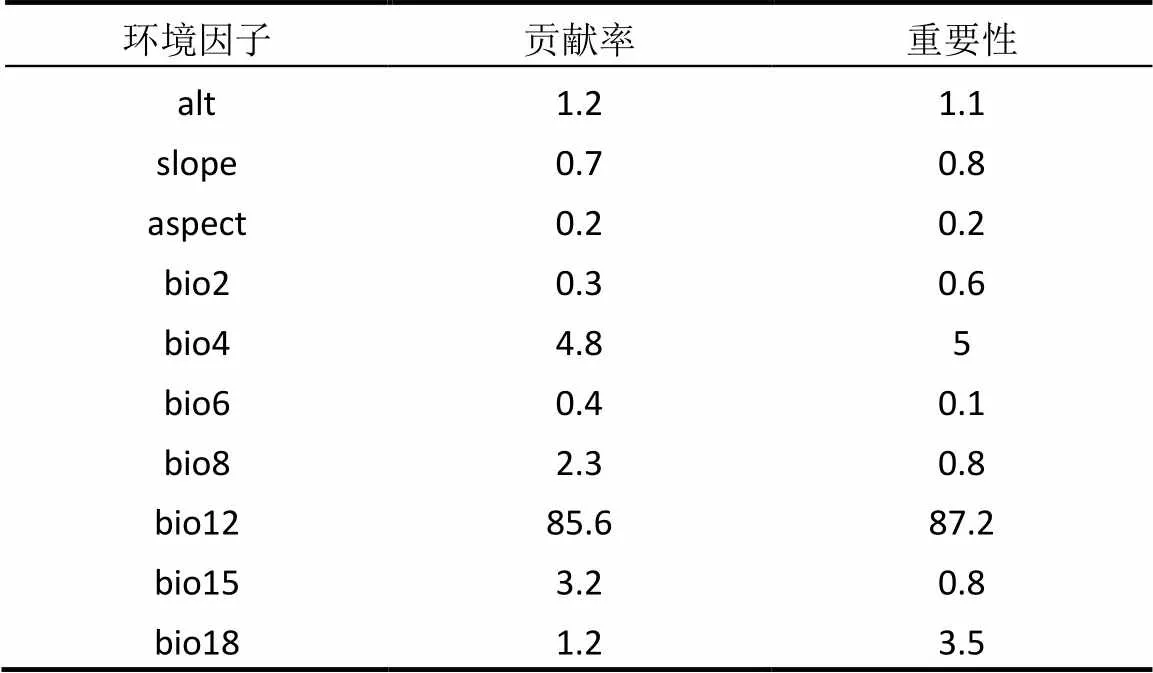
表2 环境变量贡献和重要性分析结果
环境因子的贡献和重要性分析(表2)以及基于正则化增益的刀切法(图3)对环境变量贡献度的分析结果发现,虽然两者排列顺序有一定差异,但是均显示年均降水量(bio12)的贡献度最大,这表明相对于温度和地形因素,降水量对于紫花含笑的分布影响更为重要.另外,年均降水量与其他降水量因子高度相关(表1),这也是其他降水量因子在基于刀切法的正则化训练增益较大(图3)的原因.
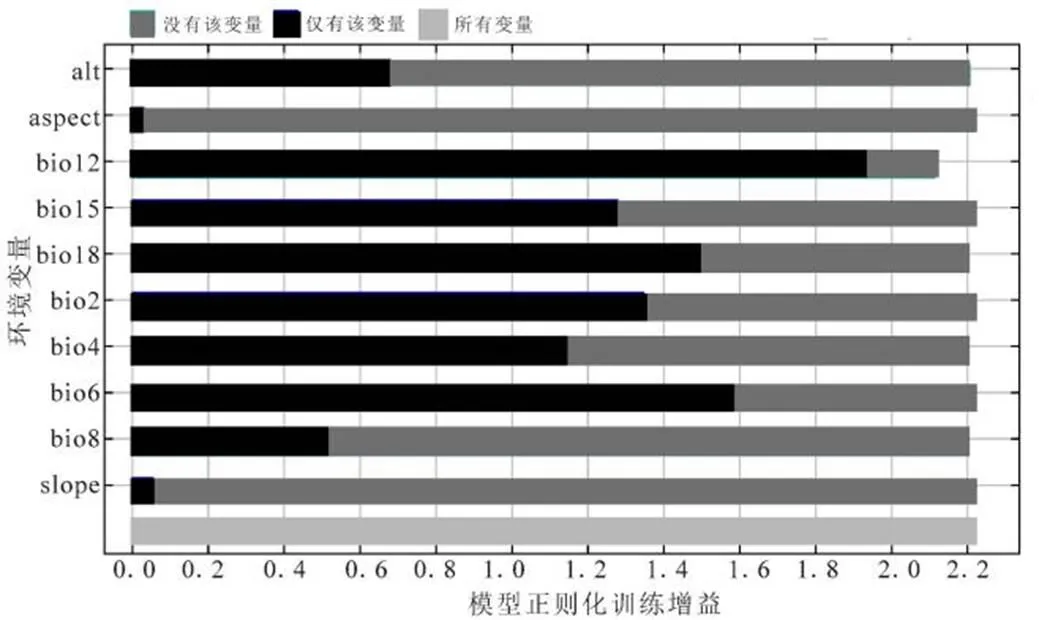
图3 基于刀切法的模型正则化训练增益
基于图3可以看到,如果在建模因子中去除年均降水量(bio12),模型的正则化训练增益的降低较多.而如果去除温度因子以及地形因子,模型的正则化训练增益则变化不大,这可能是年均降水量因子的贡献和重要性远高于其他环境因子的原因.
2.3 生境适宜度与海拔、年均降水量和年均温度的关系
海拔、年均降水量和年均温度是描述物种生境最为常用的环境因子,此处给出生境适宜度与海拔、年均降水量和年均温度的关系曲线,以便于确定紫花含笑的适宜生境.如图4生境适宜度与海拔、年均降水量和年均温度的关系均呈单峰模式,紫花含笑最适宜生境的海拔范围约在300~700m之间,年均降水量1700~1900mm,年均温度17~21℃.

2.4 紫花含笑生境适宜区空间分布
紫花含笑的生境适宜区与最适宜区主要分布在中亚热带东部地区,北纬22°~30°之间,东经107°~ 120°之间(图5).广东、福建、广西、江西和湖南等5个省份是紫花含笑生境最适宜区面积最广的地区,其次是浙江(表3).
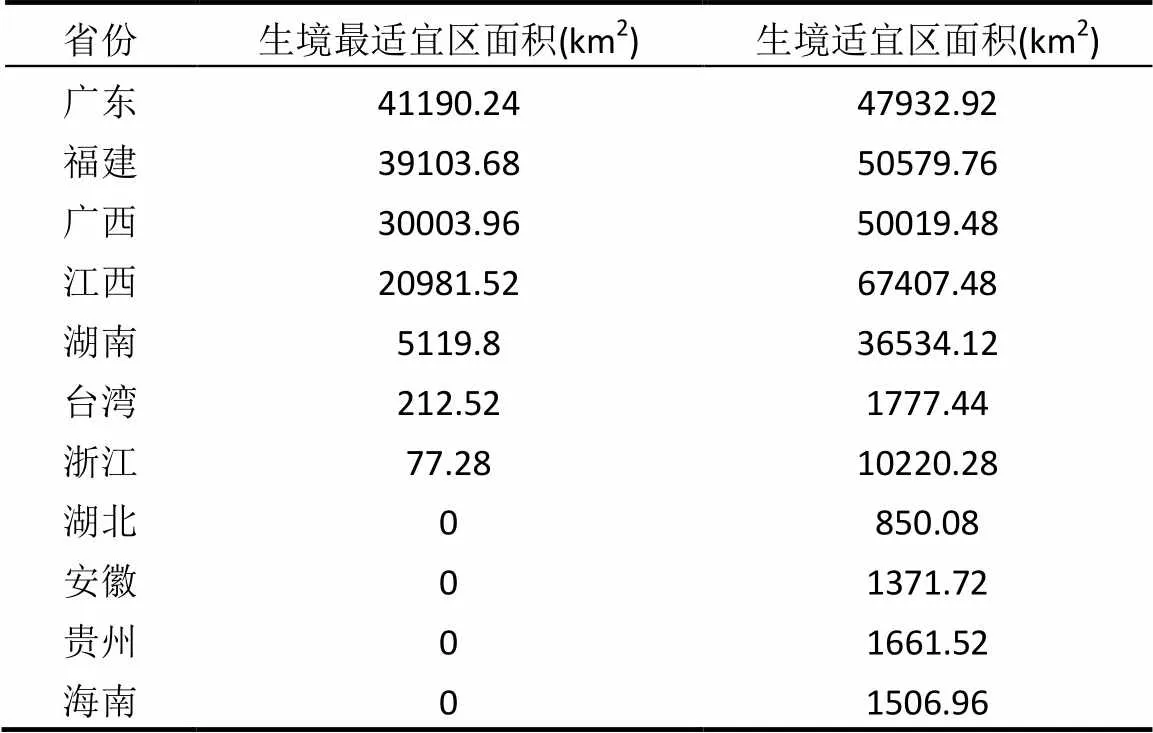
表3 紫花含笑生境最适宜区和适宜区面积分省统计结果
土地利用为2015年数据
2.5 紫花含笑生境适宜区保护和人类干扰分析
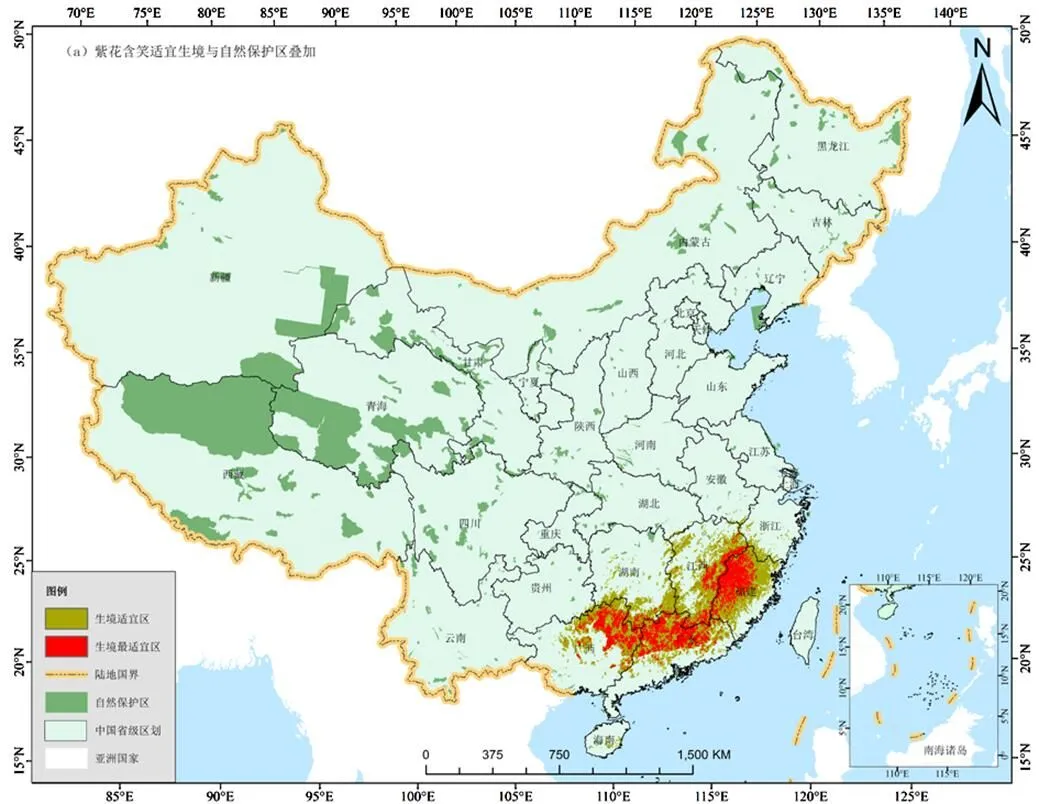
叠加分析紫花含笑生境最适宜区、适宜区与国家级自然保护区空间分布数据,如图5所示,发现处在保护区内的面积为7670km2,约占生境最适宜区和适宜区总面积的1.9%,绝大多数的生境适宜区和最适宜区未得到有效保护.
紫花含笑生境适宜区与最适宜区与土地利用数据叠加分析结果见图5.从图中可以看出,紫花含笑生境适宜区和最适宜区内,林地是最大的土地利用类型,耕地和城乡、工矿和居民用地也占有相当大的比例.
3 讨论
根据现有文献,紫花含笑为我国中亚热带东部特有,主要分布于广西东北部、广东北部、湖南南部、江西省中东部、贵州南部和东南部,分布区海拔在300~1000m[2,27].本研究中,基于Maxent的预测结果,广东北部、湖南南部、广西东北部、江西东部、福建西部等地区为紫花含笑的主要生境适宜区和最适宜区.除去福建以外,本研究中生境适宜区与中国植物志中的记载基本一致.而在福建,亦有紫花含笑栽培的报道[28],因此,可以推断在福建应存在适合紫花含笑生存的生境.对于紫花含笑分布的海拔范围,根据生境适宜度与海拔响应曲线(图4)推测,紫花含笑最适宜生境的海拔范围在300~700m之间,这在传统的紫花含笑分布区海拔范围之内.因此,本研究基于Maxent预测的紫花含笑的适宜生境分布是合理、可靠的.
自然保护区是野生物种和种子资源最为重要和有效的保护场所.然而,本研究中,紫花含笑与国家级自然保护区的叠加分析发现只有1.9%的生境最适宜区和适宜区位于国家级自然保护区内.本研究中只有国家级自然区的数据,而缺乏省、市和县等级别的保护区数据.不过,根据已经发表的数据,我国自然保护区的总面积约147万km2,国家级自然保护区面积和其他等级的保护区面积分别为96和50.5万km2[29].参考国家级自然保护区所覆盖的紫花含笑适宜生境的面积,其他级别的保护区所覆盖的紫花含笑生境适宜区的面积不会超过2%,那么受到保护紫花含笑生境适宜区和最适宜区不会超过总面积的5%,绝大多数的适宜生境处在零保护状态,这可能也是造成野生紫花含笑“濒危”的主要原因.
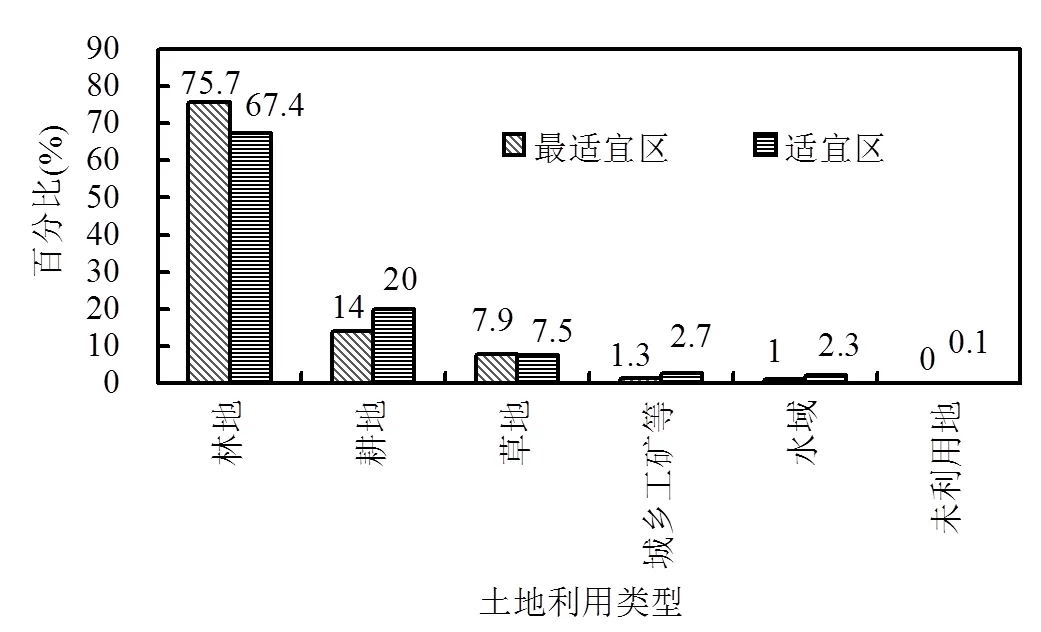
图6 紫花含笑生境最适宜区和适宜区内土地利用状况(2015年)
林地是紫花含笑生境适宜区和最适宜区内最重要的土地覆被类型(图6),这说明紫花含笑生境适宜区和最适宜区内的植被状况总体上较好.但是,在林地中,还有部分人工林和苗圃及各类园地(果园、桑园、茶园、热作林园等),而在土地利用数据中,人工林未单独分类,所以难以了解人工林在林地中的比例.此外,在生境适宜区和最适宜区内,耕地和城乡、工矿和居民用地还占有相当大的比例,这说明紫花含笑的适宜生境内还存在较大的人类活动干扰.另外,在紫花含笑生境适宜区和最适宜区内,还有部分草地的存在(7%左右),这部分草地可能是林地被砍伐以后形成的.因此,综合考虑耕地和城乡、工矿和居民用地、草地以及林地中的人工林、苗圃等,估计紫花含笑生境适宜区和最适宜区内约有30%的比例受到严重的人类干扰.
4 结论
广东北部、湖南南部、广西东北部、江西东部、福建西部等地区为紫花含笑的主要生境适宜区和最适宜区;在国家、省、市、县级自然保护区中受到保护的紫花含笑生境适宜区和最适宜区不足其总面积的5%,绝大多数的适宜生境处在零保护状态;林地是紫花含笑生境适宜区和最适宜区内最主要的土地覆被类型,但耕地和城乡、工矿和居民用地还占有相当大的比例,这说明紫花含笑的适宜生境内还存在较大的人类活动干扰.
[1] 柴弋霞,蔡梦颖,金晓玲,等.紫花含笑传粉生物学初探 [J]. 广西植物, 2017,37(10):1322-1329. Chai Y X, Cai M Y, Jin X L, et al. Preliminary study on pollination biology of[J]. Guihaia, 2017,37(10):1322-1329.
[2] 曹展波,刘仁林.江西九连山紫花含笑栖息地植物群落特征研究 [J]. 江西科学, 2013,31(4):461-464. Cao Zhanbo, Liu Renlin. Characteristics of plant communities in thehabitat of Jiulian Mountain in Jiangxi Province [J]. Jiangxi Science, 2013,31(4):461-464.
[3] 环境保护部,中国科学院.中国生物多样性红色名录[M]. 2013. Ministry of Environmental Protection, Chinese Academy of Sciences. Red List of Biodiversity in China [M]. 2013.
[4] Graham B (2001). Neutral macroecology [J]. Science, 2001,293, 2413-2418.
[5] Zhang Y B, Ma K P. Geographic distribution patterns and status assessment of threatened plants in China [J]. Biodiversity and Conservation, 2008,17:1783-1798.
[6] Guisan A, Thuiller W. Predicting species distribution:offering more than simple habitat models [J]. Ecology Letters, 2005,8:993-1009. Hampe A (2004). Bioclimate envelope models: what they detect and what they hide [J]. Global Ecology and Biogeography, 2004,13:469-471.
[7] Calenge C. The package “adehabitat” for the R software: a tool for the analysis of space and habitat use by animals [J]. Ecological Modelling, 2006,197:516-519.
[8] Vayssieres M P, Plant R E, Allen-Diaz B H. Classification trees: an alternative non-parametric approach for predicting species distribution [J]. Journal of Vegetation science, 2000,11:679-694.
[9] Phillips S J, Anderson R P, Schapire R E. Maximum entropy modeling of species geographic distributions [J]. Ecological Model, 2006,190: 231-259.
[10] Stockwell D, Peters D. The GARP modeling system:problems and solutions to automated spatial prediction [J]. International Journal of Geographical Information Science, 1999,13:143-158.
[11] Van P A, Sosef M, Wieringa J J, Raes N(2015). Minimum required number of specimen records to develop accurate species distribution models [J]. Ecography, 2015,39:542-552.
[12] Saupe E E, Qiao H J, Hendricks J R, et al. Niche breadth and geographic range size as determinants of species survival on geological time scales [J]. Global Ecology & Biogeography, 2015,24:1159-1169.
[13] Elith J, Graham C H, Anderson R P, et al. Nevel methods improve prediction of specie’s distribution from occurrence data [J]. Ecography, 2006,29:129-151.
[14] Hijmans R J, Cameron S E, Parra J L, et al. Very high resolution interpolated climate surfaces for global land areas [J]. International Journal of Climatology, 2005,25:1965-1978.
[15] Dormann C F, Elith J, Bacher S, et al. Collinearity:a review of methods to deal with it and a simulation study evaluating their performance [J]. Ecography, 2013,36:27-46.
[16] Yackulic C B, Chandler R, Zipkin E F, et al. Presence-only modelling using Maxent: when can we trust the inferences? [J]. Methods in Ecology and Evolution, 2013,4:236-243.
[17] Harte J, Zillio T, Conlisk E, et al. Maximum entropy and the state- variable approach to macroecology [J]. Ecology, 2008,89:2700-2711.
[18] Phillips S J, Dudik M. Modeling of species distributions with Maxent:new extensions and a comprehensive evaluation [J]. Ecography, 2008,31:161-175.
[19] Phillips S J, Anderson R P, Dudík M, et al. Opening the black box:an open-source release of Maxent [J]. Ecography, 2017,40:887-893.
[20] Swets J A. Measuring the accuracy of diagnostic systems [J]. Science, 1988,240:1285-1293.
[21] Waltari E, Guralnick R P. Ecological niche modeling of montane mammals in the Great Basin, North America: examining past and present connectivity of species across basins and ranges [J]. Journal of Biogeography, 2009,36:148-161.
[22] Brito J C, Acosta A L, Álvares F, et al. Biogeography and conservation of taxa from remote regions: An application of ecological-niche based models and GIS to North-African canids [J]. Biological Conservation, 2009,142:3020-3029.
[23] Newbold T, Gilbert F, Zalat S, et al. Climate-based models of spatial patterns of species richness in Egypt's butterfly and mammal fauna [J]. Journal of Biogeography, 2009,36:2085-2095.
[24] Ramírez-Barahona S, Torres-Miranda A, Palacios-Ríos M, et al. Historical biogeography of the Yucatan Peninsula, Mexico: a perspective from ferns (Monilophyta) and lycopods (Lycophyta) [J]. Biological Journal of the Linnean Society, 2009,98:775-786.
[25] Donegan T M & Avendaño J E. A new subspecies of mountain tanager in the Anisognathus lacrymosus complex from the Yariguíes Mountains of Colombia [J]. Bulletin of the British Ornithologists Club, 2010,130:13-32.
[26] 中国科学院中国植物志编辑委员会.中国植物志[M]. 北京:科学出版社, 1996. Chinese Flora Editorial Board of the Chinese Academy of Sciences. Chinese flora [M]. Beijing: Science Press, 1996.
[27] 陈孝丑.紫花含笑扦插繁殖试验[J]. 福建林业科技, 2014,41(3): 128-131.Chen Xiaochou. Experiment on the propagation ofcuttings [J]. Journal of Fujian Forestry Science and Technology, 2014,41(3):128-131.
[28] 王 静,孙军平,石 磊,等.中国自然保护区建设的现状、存在问题及展望[J]. 中国人口·资源与环境, 2016,26(5):270-273. Wang Jing, Sun Junping, Shi Lei et al. Current situation, existing problems and prospects of construction of nature reserves in China [J]. China Population • Resources and Environment, 2016,26(5):270-273.
Conservation status and human disturbance of the habitats of Michelia crassipes Law in China.
LIU Hui-ming1, GAO Ji-xi1*, SONG Chuang-ye2, YU Sheng-xiang2
(1.Satellite Environment Center, Ministry of Environmental Protection, Beijing 100094, China;2.The Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China)., 2019,39(9):3976~3981
This research aims to assess the habitat suitability ofLaw based on the species distribution data derived from specimen records and environmental variables through the approach of MaxEnt. The predicted habitat suitability was divided into suitable area and most suitable area according to the selected threshold. National reserve and land use data were used to conduct the analysis of conservation status and human disturbance of suitable habitat. The results show: Compared with topography variables and temperature variables, annual precipitation was the most important variables which influences the habitat suitability. The suitable habitat forLaw was mainly located in the east area of middle subtropics, between 22 and 30degrees north latitude and between 107 and 120 degrees east longitude. The area of most suitable habitat and suitable habitat were 137000km2and 270000km2. Guangdong, Guangxi, Hunan, Jiangxi and Fujian share the largest area of suitable habitat. Only 1.9% of the suitable habitats were under the protection of national reserves. Large areas of the suitable and most suitable habitats were left in a state without any protection. Woodland occupy the largest area of the suitable and most suitable habitat. However, human disturbance also occupy big area of suitable and most suitable habitat.
habitat suitability;maxent;receiver operating curve;land use;nature reserves;human disturbance
X826
A
1000-6923(2019)09-3976-06
刘慧明(1982-),女,山西临县人,正高级工程师,博士,主要从事生态遥感监管研究工作.发表论文20余篇.
2019-02-15
国家科技支撑计划项目(2012BAC01B08)
* 责任作者, 研究员, gjx@nies.org
猜你喜欢
落叶果树(2021年6期)2021-02-12
扬子江诗刊(2020年3期)2020-11-17
农民致富之友(2020年16期)2020-06-19
扬子江(2020年3期)2020-06-08
湖北农机化(2020年3期)2020-01-09
现代园艺(2018年1期)2018-03-15
周末·校园文学(2017年17期)2017-07-08
体育科技(2016年2期)2016-02-28
绿色中国(2015年12期)2016-01-05
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
