
闽江口龙头鱼(Harpadon nehereus)的生物学参数及资源评估
2019-09-25 11:22:12郭峻宏何雄波
广东海洋大学学报 2019年5期
郭峻宏,李 军,沈 忱,石 焱,冯 晨,何雄波,康 斌,2
闽江口龙头鱼()的生物学参数及资源评估
郭峻宏1,李 军1,沈 忱1,石 焱1,冯 晨1,何雄波1,康 斌1,2
(1. 集美大学水产学院, 福建 厦门 361021;2. 中国海洋大学水产学院, 山东 青岛 266003)
【】调查闽江口龙头鱼资源,估算其生物学参数。于 2015 年1月、5月、8月、11月在福建闽江口11个采样点(25.85°―26.29° N,119.65°―119.87° E)采集龙头鱼3 770尾,测量其体长、体质量,用FiSAT Ⅱ软件中的ELEFAN I方法,估算龙头鱼的生长、死亡参数及其开捕体长和单位补充量渔获量。闽江口渔场龙头鱼体长 () 体质量 () 关系为:=0.000 73.804 2(2=0.961 1);龙头鱼Von Bertalanffy生长方程估算的生长参数、∞、∞、0分别为0.52、28.7 cm、246.1 g和-0.317龄;由生长方程推算出龙头鱼的拐点年龄约为2.252龄,拐点体长约为21.2 cm,拐点体质量约为77.1 g;根据长度变换渔获曲线法估算龙头鱼的总死亡系数为1.67,由Pauly经验公式估算自然死亡系数为1.04,捕捞死亡系数为0.63,开发率为0.377;由动态综合模型估算出龙头鱼的开捕年龄为0.712龄,对应开捕体长为11.9 cm。闽江口龙头鱼资源处未过度开发状态。
龙头鱼;闽江口;生长参数;开发率;单位补充量渔获量
龙头鱼()隶属于硬骨鱼纲(Osteichthyes),仙女鱼目(Aulopiformes),狗母鱼科(Synodontidae),龙头鱼属()[1],属大陆架中下层鱼类[2],我国主要分布于南海、台湾海峡、东海和黄海[3]。近年来,海洋生态环境受到严重破坏,导致传统经济鱼种资源严重衰退[4-6],以龙头鱼为代表的次级经济鱼类逐步成为各渔场优势种群[4]。龙头鱼适应性较强,生物量逐年上升,作为连接闽江口海域食物网中高级消费者的重要纽带,龙头鱼在闽江口海域中占有重要位置[5, 7],研究其渔业生物学特征有重要意义。
国内有关龙头鱼的研究并不多见,主要有2005年林龙山[8]对东海区龙头鱼数量分布的分析、2008―2009年林显鹏等[5]对东海龙头鱼摄食习性的研究、2008―2009年罗海舟等[9]对东海龙头鱼生物学特性的研究、2011―2012年陈玲等[10]对舟山海域龙头鱼生长特征的研究、2012年晏磊等[11]对南海北部龙头鱼刺网捕捞性能的研究。国外的相关研究多集中在南亚海域,仅见1995―1996年Nurul Amin[12]对孟加拉国浅海区龙头鱼年龄与生长及环境因素的研究、2003―2006年Ghosh等[13]对索拉什特拉海岸龙头鱼渔业和种群动态的研究。本研究以 2015 年4个季节福建闽江口的调查数据为基础,通过研究龙头鱼体长与体质量的生长特性,用ELEFAN I方法,以体长频率估计龙头鱼的生长、死亡参数,运用Beverton-Holt动态综合模型评估种群资源的状况,并确定最适开捕规格,为龙头鱼的合理开发和资源利用提供依据。
1 材料与方法
1.1 样品采集及鉴定
本次采样分四次进行,分别在2015年1月(冬季)、5月(春季)、8月(夏季)、11月(秋季)于福建闽江口海域设置的11个采样点,进行底拖网作业采样(图1)。调查渔船网具尺寸为7.5 m×3 m×45 m,网口网目为4.5 cm,囊网网目为2.5 cm,据调查对象的游泳能力和调查船性能,每个站位的平均拖速为3.1 ~ 4.6 kn(约5.74 ~ 8.52 km/h),每个站点作业时间约为30 min。
图1 采样站位点
Fig. 1 Sampling sites
样品采集与处理参照GB/T 12763.6–2007[14]进行。样品分类及鉴定参考《中国鱼类系统检索》[15]和《福建鱼类志》[3]。渔获样本在现场处理后冰冻保存,航次结束后带回实验室进行分类、测定并记录。生物学测定包括体长和体质量,体长测定精确到0.1 cm,体质量测定精确到0.1 g。
1.2 方法
1.2.1 体长与体质量关系 用幂函数关系式拟合鱼类的体长与体质量关系:
=aL[16]。
式中,为鱼类的体长(cm),为相对应的体质量(g);、两常数为待定参数,并称为生长的条件因子,为幂指数系数[17]。
1.2.2 组距的选择 ELEFAN I方法克服了在年龄推算过程中存在的主观性误差[18],相比于传统的年龄鉴定方法,长度频率法使得鱼类生长参数的估计更加便捷、准确[19]。
体长组距则对龙头鱼的生长参数估算起决定性影响,选择合适组距可提高体长频率分析的可信度[20]。参考生长性能指标(Growth performance index)估算不同组距的可信度:=lg+2lg∞[21],并结合以ELEFAN I方法计算出的拟合优度选取最终组距。
通过Snedecor和Sturges[20]两个体长组距选择的公式分别为
=R/(4 R /σ),
=R/(1 +3.322 lg)。
式中,为组距,为体长的极差,为体长的标准差,为个体数(尾)。由不同组距得到对应的生长速率()和渐进体长(∞),并计算生长性能指标,结合拟合优度选择最佳组距。
1.2.3 Von Bertalanffy生长方程 以4个季度闽江口龙头鱼的体长频率为数据,使用ELEFAN I (electronic length frequency analysis I)[18]方法拟合龙头鱼的生长曲线,并估算其生长参数(∞为渐近体长,为生长速率[17])。
拟合出生长曲线后,选取拟合优度最大且最合理时所对应的∞和作为龙头鱼生长参数的估计值[22]。运用Von Bertalanffy生长方程[17]拟合龙头鱼的生长,其生长过程的特征变化采用生长速度、生长加速度及生长拐点的方式来描述。描述生长过程的方程为[17]
L=∞,
m=∞。
式中,为年龄;∞为渐近体长;L为龄时的体长;∞为渐近体质量;m为龄时的体质量;为生长速率;0为理论生长初始年龄,表示理论上体长和体质量等于零时的年龄。
理论生长初始年龄(0)则根据Pauly提出的经验公式估算[23]:
lg (–0) =–0.392 2–0.275 lg∞–1.038 lg。
1.2.4 生长速度及加速度 研究鱼类的生长速度、生长加速度(一般情况下均指体质量)及生长拐点,对于合理利用渔业资源有重要意义[17]。根据Von Bertalanffy生长模型,可通过鱼类生长方程推算出鱼类的生长速度、生长加速度和生长拐点年龄[17]。
鱼类生长速度、生长加速度、生长拐点年龄的计算公式:
,
,
tp=(ln)/+0。
式中,tp为生长拐点年龄,即生长速度达到最大值时或生长加速度为0时所对应的年龄。
1.2.5 死亡系数及开发率 运用长度变换渔获曲线法[17]估算总死亡系数():
lnN=ln0–
式中,为总死亡系数,N为龄时龙头鱼资源量,0为0龄(理论生长初始年龄)时龙头鱼资源量。
采用Pauly提出的经验公式[24]估算自然死亡系数():
lg=–0.0066–0.279lg∞+0.6543lg+0.4634lg。
式中,为自然死亡系数;为闽江口采样点龙头鱼栖息环境的年平均温度,采样时测得并计算平均温度为21.6℃。
捕捞死亡系数()和开发率()则根据总死亡系数和自然死亡系数计算[17],公式分别为
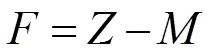
=(–) /=/。
式中,为捕捞死亡系数,为开发率。
1.2.6 开捕体长 根据体长渔获曲线拟合出的直线方程,推算线性回归中未被使用的各点相对应的期望值ln (/Δ),并计算各点的观测值与期望值的比,以及这些点的比率累积率,选取累积率达到50%的点,其所对应的体长即为平均选择体长的估计量(c),也称为开捕体长[25]。
1.2.7 单位补充量渔获量 Beverton和Holt[26]提出了Beverton和Holt稳定状态模型,并提出一套公式来表达渔获量与各种鱼群参数之间的复杂关系。当公式中的任一参数发生改变时,可观察到渔获量()或单位补充量渔获量()也会受到影响而发生改变,这就是动态综合模型(dynamic pooled model),也称Beverton和Holt稳定状态模型(简称B-H模型)[17]。
该模型假定捕捞格局(主要指开捕年龄和捕捞强度)长时间不变,所有个体一经补充均有被捕的可能性,该模型经过Pauly和Soriano[27]的调整,在渔业研究中被用来预测相对单位补充量渔获量()和相对单位补充量生物量()。在FiSAT Ⅱ软件中,模型和模型以刀刃式选择假设模型(knife-edge model)[28]建立。其中,开发率()分为以下3种水平:1)0.1,指边际增长减少10%时的开发率;2)0.5,指资源量下降到原始水平50%时的开发率;3)max,指获得最大渔获量的开发率[29]。
2 结果
2.1 资源状况
进行生物学测定的闽江口龙头鱼样本共3 770尾,累计质量为75 032.0 g。从龙头鱼季节分布的情况看,秋季捕获数量最多,春季最少;从龙头鱼的体长、体质量来看,冬季的平均体长和平均体质量均最大,秋季的平均体长和平均体质量均最小;从龙头鱼的资源密度来看,秋季的数量密度最大,春季的数量密度最小,而生物量密度的最大季节和最小季节分别为夏季和春季(表1)。
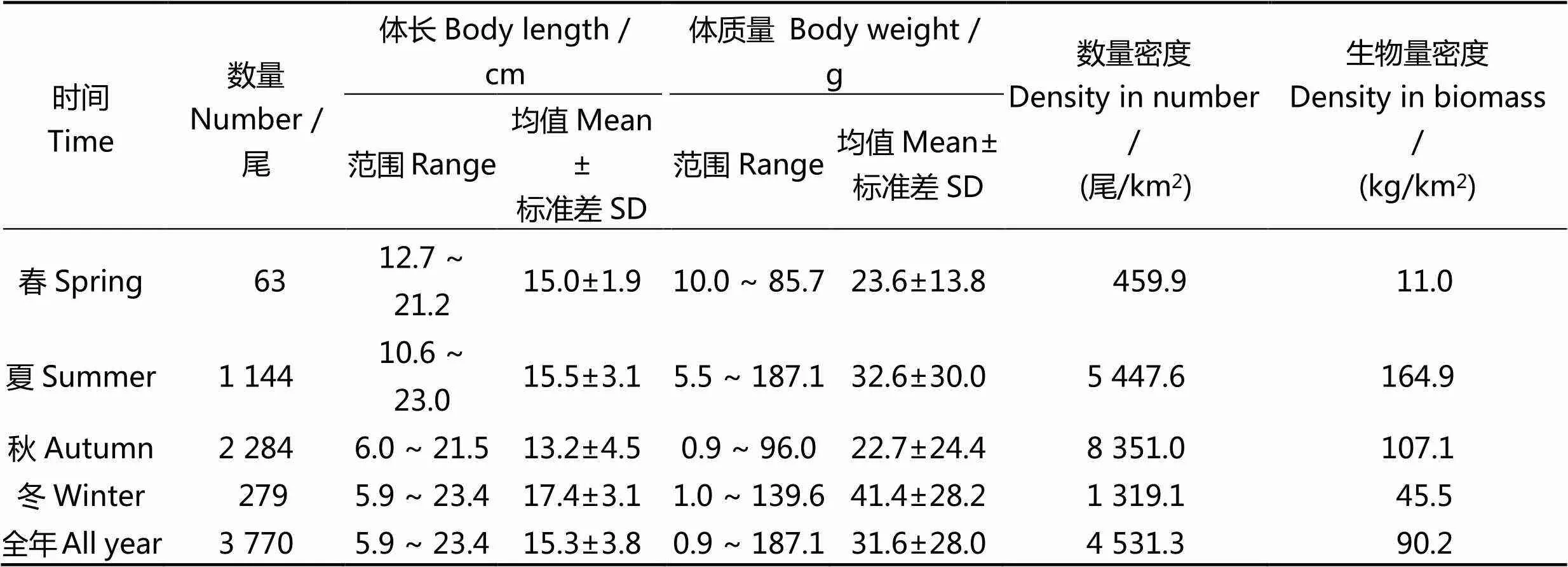
表1 2015年闽江口龙头鱼的资源概况
2.2 组距的选择
通过Snedecor和Sturges[20]两个体长组距选择的公式计算出最佳组距分别为0.9、1.8 cm。当组距为0.9 cm时,拟合优度和生长性能指标的值在7组结果中均较低;当组距为1.8 cm时,拟合优度为0.409,生长性能指标为2.606,均位于中等。在Snedecor和Sturges两结果均不合适时,选取二者均值或其附近的值为最佳体长组距[20]。当组距为1.5 cm时,拟合优度最大,为0.434,此时拟合效果最佳,但其对应的生长性能指标仅为2.434,在7组结果中较低;当组距为1.2 cm时,拟合优度和生长性能指标值在7组结果中均较高,可信度较高。综合考虑,将1.2 cm选为最佳体长组距(表2)。
2.3 体长与体质量关系
根据所测得体长和体质量数据,拟合出其体长与体质量的回归关系曲线,两者的关系呈幂函数:
0.00073.8042(20.961 1)。
其中,幂指数系数>3,表明闽江口龙头鱼呈正异速生长(图2)。
2.4 Von Bertalanffy生长方程
运用ELEFAN I方法来拟合闽江口龙头鱼的Von Bertalanffy生长曲线,并估算龙头鱼生长参数,分别=0.52,∞28.7 cm。根据Pauly提出的经验公式可估算出理论生长的初始年龄0=-0.317。根据体长体质量关系公式,可由∞计算出∞∞=246.1g。

表2 2015年闽江口龙头鱼不同体长组距的生长参数估算
龙头鱼的体长和体质量的生长方程分别为:
L28.7 [1–e-0.52 (t+0.317)],
m246.1 [1–e-0.52 (t+0.317)]3.804 2。
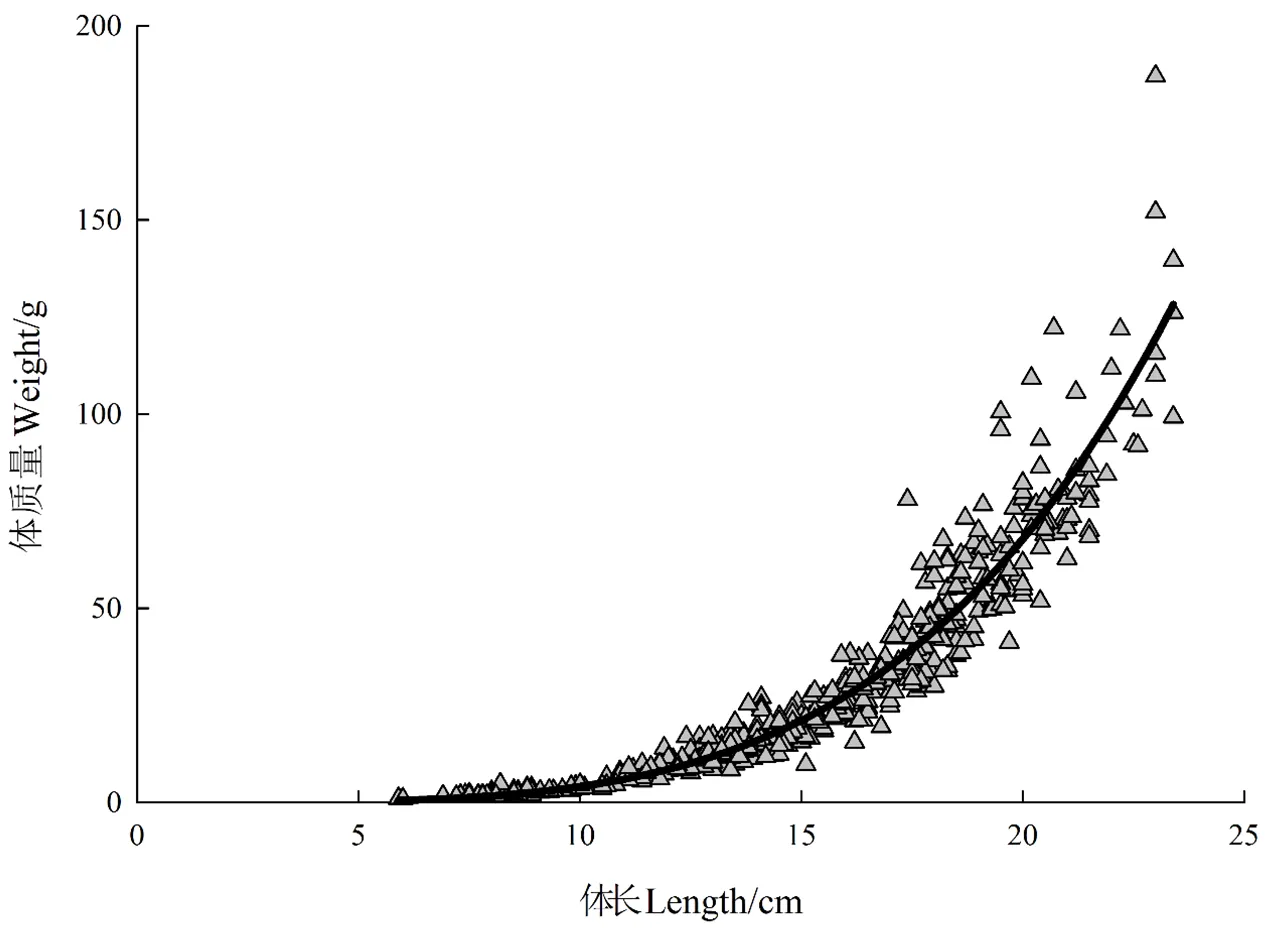
图2 龙头鱼的体长与体质量关系
2.5 生长速度及加速度
在体质量生长速度达到最大值时或生长加速度为0时,所对应的年龄即为体质量生长拐点年龄,估算出其生长拐点年龄tp约为2.252龄,此时可通过龙头鱼的生长方程估算出其对应的拐点体长约为21.2 cm,拐点体质量约为77.1 g。
龙头鱼的体质量生长速度曲线呈先增后减型,在生长拐点前,随着年龄的增长而增大;在生长拐点时,达到最大值;在生长拐点后,随着年龄的增长而减小并趋近于0。在生长拐点前,体质量生长加速度则随年龄的增长而先增大,至最大值后减小;达到生长拐点时,体质量生长加速度为零;生长拐点过后则为负值,至最小值后增大并趋近于0。
2.6 死亡系数及开发率
根据长度变换渔获曲线法估算闽江口龙头鱼的总死亡系数,数据点的选取以“未全面补充年龄段,以及体长接近渐近体长的年龄段不能作为回归分析的数据点”为原则,选择8个数据点进行线性回归分析(图3),并拟合总死亡系数的线性回归分析方程为
ln (/Δ) =-1.67+8.23 (2=0.859 1)。
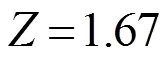

2.7 开捕体长
根据渔获量曲线拟合关系继续推算线性回归中未被使用的各个数据点的ln(/Δ),计算出各点观测值与期望值之比的累积率,选取累积率达到50%的点,其所对应的体长作为平均选择体长的估计量c,即开捕体长,c50%11.9cm,根据生长方程L28.7[1–e-0.52 (t +0.317)]求得对应的开捕年龄c0.712。
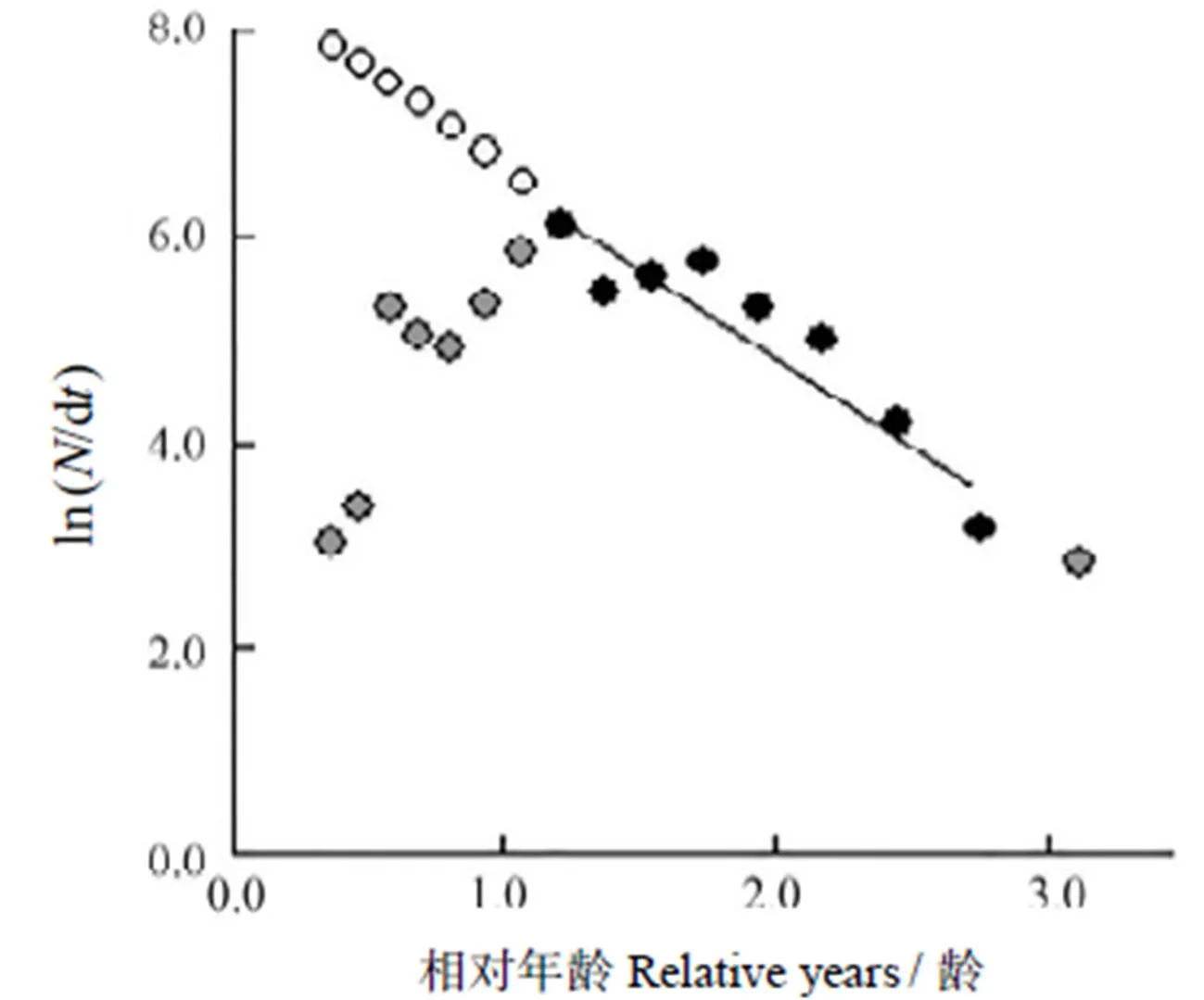
图3 据龙头鱼体长变化渔获曲线估算总死亡系数
2.8 单位补充量渔获量
运用ELEFAN I方法绘制龙头鱼相对单位补充量渔获量()与开发率和c/∞的关系图,其变化趋势用等值线表示(图4)。图4中点为采捕时捕捞状态,即=0.377,/=2.000,c/∞=0.415,=0.025;点则为逻辑上的最佳开发状态,此时,在/=2.000不变的情况下,达到最大值0.035,与此对应的约为1.000,c/∞约为0.700。
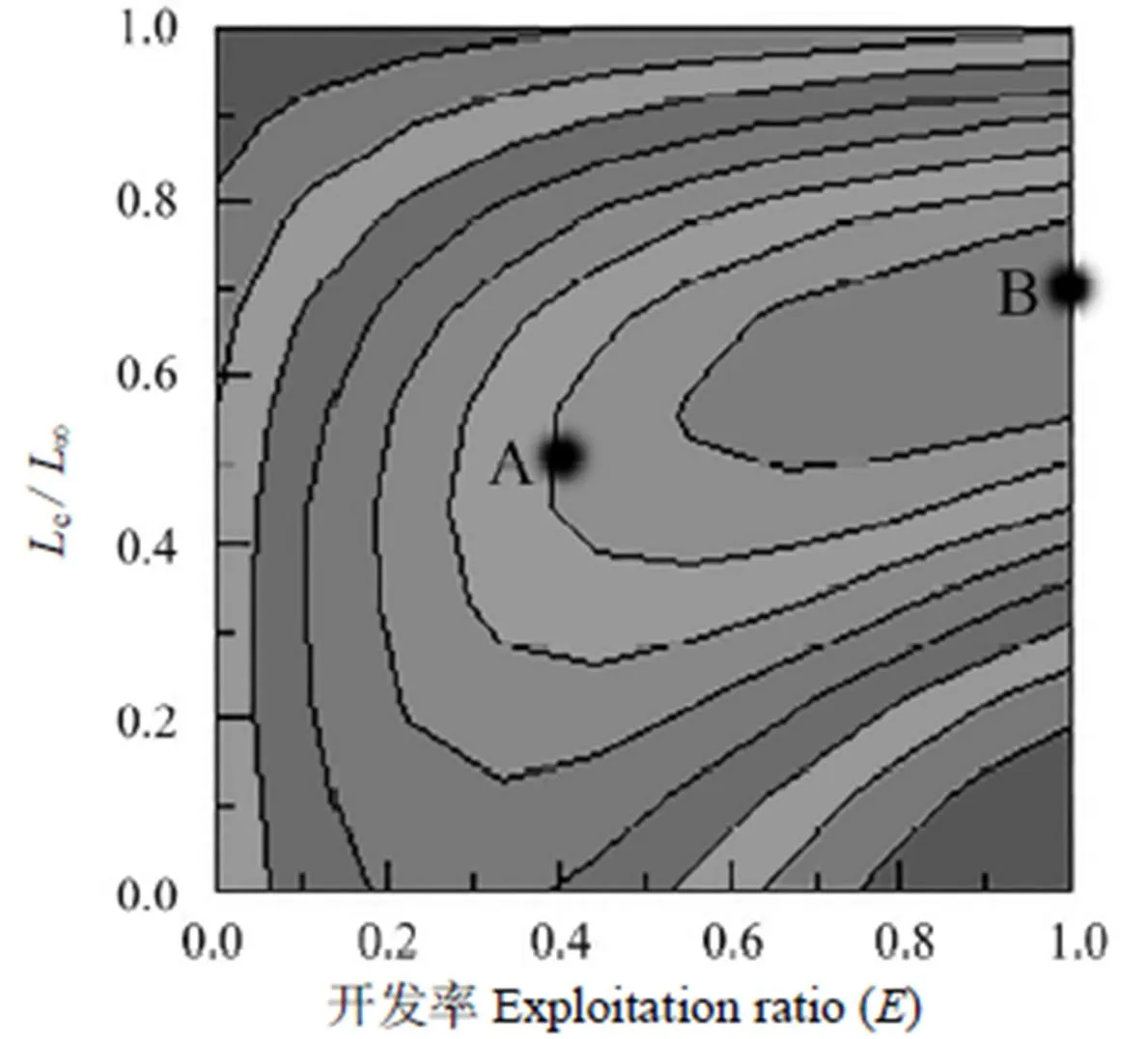
图4 2015年闽江口龙头鱼相对单位补充量渔获量与开发率和开捕体长的关系
从点调整到点,的值由0.025增加到0.035,增幅约为40%,对应的开发率和c/∞的增幅分别约为150.63%和68.67%,而开发率的增加则预示着捕捞强度的增大。当=0.377,=2.000保持稳定时,提高开捕体长c(在c/∞>0.415的前提下)的值时,会随着c/∞的增大而增大,当c/∞提升至约0.550时(c为15.8 cm,对应的c为1.219),则会达到相应的最大值0.025,当c/∞继续增大时,则会下降。相比于采捕时的开捕体长(c=11.9 cm),当开捕体长为15.8 cm时,二者所对应的并无差异,均为0.025。
相对单位补充量渔获量()与开发率()的二维分析图(图5)可以明确地显示2015年闽江口龙头鱼资源的开发程度,此时,开捕体长c=11.9 cm,0.1=0.562,0.5=0.338,max=0.662。当c=11.9 cm,<0.377时,随着开发率的增加而增加;当c=11.9 cm,>0.377时,随着开发率的增加而先增加后减小。当开发率减小至0.5(0.338)时,对应的0.023;当开发率继续增加至0.1(0.562)时,对应的值会大幅增加,即0.029;当开发率继续增加至max(0.662)时,对应的值增幅不大但达到最大值,即0.030。
从维持生物量资源合理开发和利用的角度来看,当max=0.662时,仅约17.0%的生物量资源得到保持;当0.1=0.562时,仅有约26.3%的生物量资源得到保持;而当0.5=0.338时,则有约50.0%的生物量资源可得到保持。
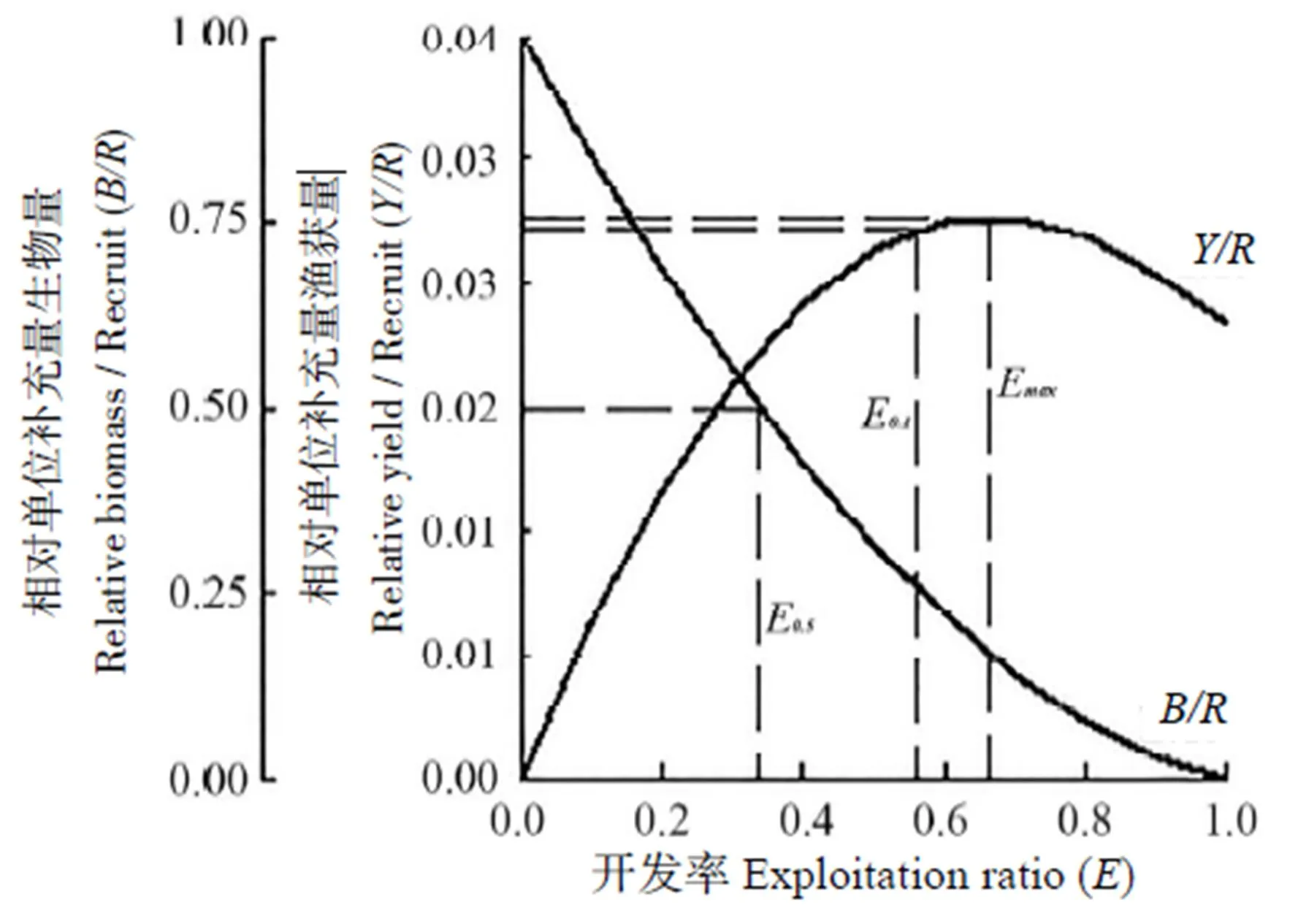
图5 2015年闽江口龙头鱼开捕体长下的Y/R和B/R二位分析曲线
3 讨论
3.1 龙头鱼的资源状况
龙头鱼有洄游习性[30],每年10月后水温下降,龙头鱼群体逐渐向外海域移动至深水区越冬,翌年的3 - 4月起,开始游向近岸;6月起,龙头鱼群体到达闽江河口区域,进行分散产卵,而7 - 9月时,成鱼及当年生幼鱼进行索饵育肥[9, 31],导致本研究中春季龙头鱼的数量较少,夏秋季节的数量较多(表1)。此外,夏季的龙头鱼生物量密度大于秋季,但数量密度却远小于秋季。
基于扫海面积法所得的龙头鱼全年的数量密度为4 531.3尾/km2,生物量密度为90.2 kg/km2(表1),2006年黄良敏[32]和王家樵等[33]的结果中,龙头鱼全年的数量密度为8 257.6尾/km2,生物量密度为103.3 kg/km2,比较而言,本研究中龙头鱼资源密度有所降低。
3.2 龙头鱼的相关参数
3.2.1 闽江口不同时间龙头鱼的参数对比 在2006年黄良敏[32]和王家樵等[33]对闽江口龙头鱼的生长状况研究中,采样地点及研究区域与本研究极为相近。他们的结果表明,2006年闽江口龙头鱼的生长速率为2.10,渐近体长∞为31.5 cm,渐近体质量∞为297.9 g,总死亡系数为2.41,幂指数系数为3.7569。本研究中闽江口龙头鱼的幂指数系数较高,生长条件因子无差异,但生长速率低,渐近体长和渐近体质量均小,体长范围区间亦小,总死亡系数低但自然死亡系数高(表3)。2006年采样地的年平均温度约为20.7 ℃,本研究中采样地的年平均温度约为21.6 ℃,增长约1 ℃。温度不同导致营养条件不同,显著影响值[16]。近年来闽江口海域人类活动频繁,闽江上游的水库[34]、电站[35]等,下游的污水排放[36],河口区域码头建设[37]以及赤潮的暴发[38],均影响闽江口渔场的水质和龙头鱼的生长及生存,龙头鱼的生长速率、渐近体长、渐近体质量、体长范围区间均有所减小,呈现个体小型化趋势。龙头鱼的总死亡系数降低但自然死亡系数增高,且自然死亡系数在总死亡系数中的比例增大。造成小型化趋势及自然死亡率增大的原因可能是水域温度的提升,也可能是人类活动对闽江口水域环境造成的破坏和过度捕捞所致[39]。
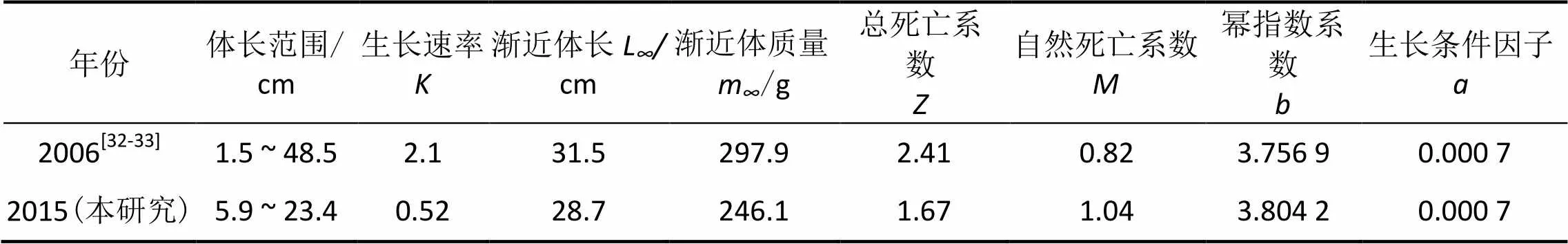
表3 闽江口2006年和2015年的龙头鱼参数
3.2.2 不同地域龙头鱼的参数对比 国内其他海域龙头鱼的研究并不多,除本研究外,有关龙头鱼的研究仅有4个海域(表4),分别为罗海舟等[9]对东海海域、陈玲等[10]对舟山海域、杜晓雪等[40]对温台渔场、晏磊等[11]对南海北部的研究。由生长速率、渐近体长、渐近体质量、体长范围区间可看出,龙头鱼均有个体小型化的趋势。对比发现,本研究估算的龙头鱼生长速率0.52,为相关研究海域中生长速率最快的,东海海域的龙头鱼生长速率其次,温台渔场的龙头鱼生长速率再次,可能是由于闽江口海域为河口区域,饵料丰富,龙头鱼的生长速率快。
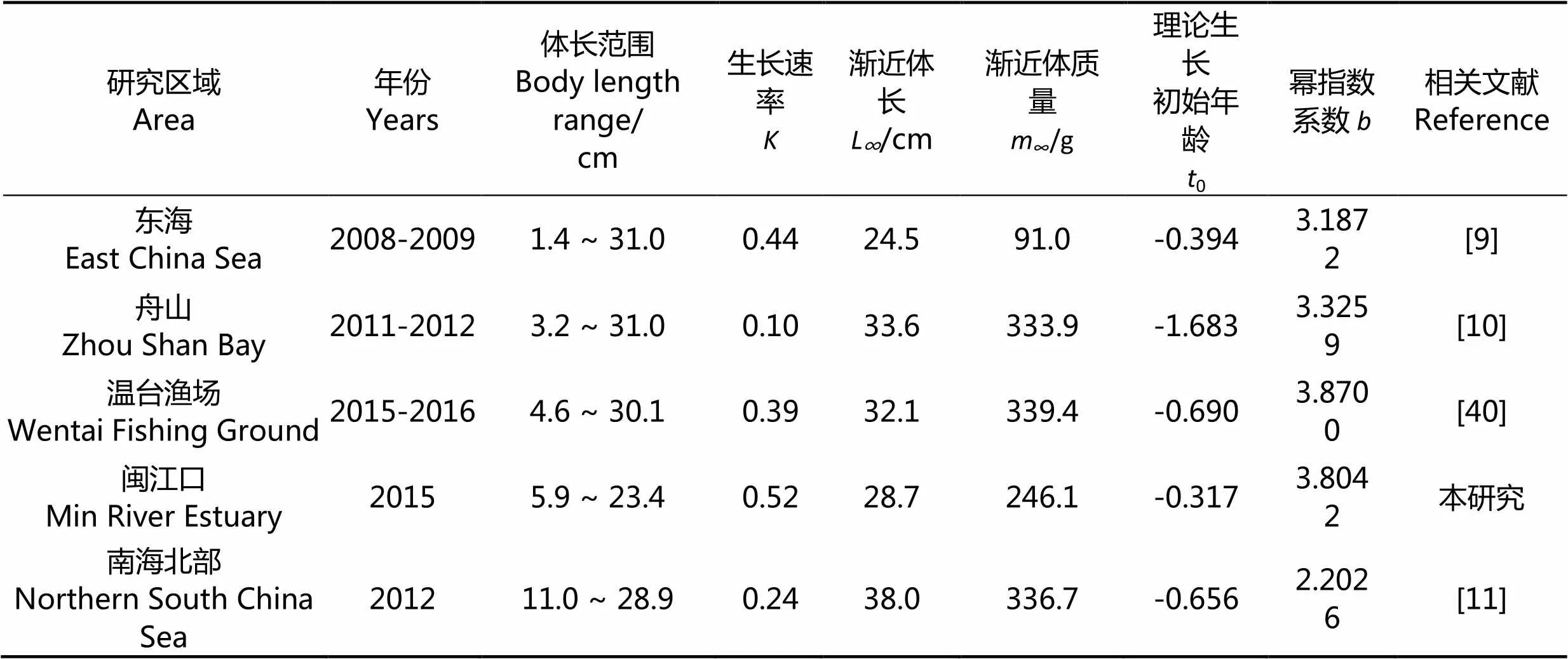
表4 国内不同海域的龙头鱼参数
对比体长范围、渐近体长和渐近体质量,东海龙头鱼的体长范围跨度最大,但渐近体长和渐近体质量均较小,表明该海域龙头鱼分布不均匀,可能是纬度较高、温度低导致龙头鱼的生长受到影响。本研究和南海北部龙头鱼的最大体长均未超过30 cm,相比于南海北部,本研究的渐近体长和渐近体质量结果均降低,表明闽江口海域的龙头鱼个体偏小,而南海北部的捕捞压力可能过大。闽江口海域隶属于东海渔场,与东海[9]研究海域渐近体长和理论生长初始年龄相仿,但样品获取年份相隔较远,使得体长范围、生长速率、渐近体质量及幂指数系数略有差异。温台渔场同样隶属于东海渔场,临近闽江口海域,且样品获取时间与本研究相近,幂指数系数的结果差异不大,但闽江口海域的体长区间较小,且生长速率等其他四项指标亦有明显差异。闽江口海域的地理位置比温台渔场偏南,因此水域温度不同导致龙头鱼新陈代谢和生长速率差异,也可能是由于两地的采捕强度和网目尺寸的不同造成闽江口龙头鱼渔获个体略小。舟山海域和南海北部的龙头鱼生长速率较慢,或因水域环境及温度等因素导致体长范围和幂指数系数稍有差异,两海域渐近体长和渐近体质量较大且差异甚微,龙头鱼成鱼个体可能普遍大于闽江口海域。
3.3 闽江口龙头鱼的资源开发
本研究估算龙头鱼的开发率为0.377,Gulland[41]认为,一般鱼类的最适开发率为0.5,并以此判断某海域渔业资源的开发程度。Mehanna[42]则认为,在相对单位补充量渔获量与开发率关系的曲线中,max点的左侧区域是某海域渔业资源得以可持续发展的安全区域,本研究中闽江口龙头鱼的开发率(0.377)点位于max(0.662)点的左侧,属于可持续发展的安全区域。二者均说明闽江口龙头鱼的渔业资源处于未过度开发状态,尚可持续开发利用。
在传统渔业资源严重衰退的情况下,龙头鱼在闽江口海域的生态地位不断上升,逐步成为该渔场的优势种群[4]。但本研究发现,随着捕捞强度的不断增大,以及捕捞过度和生态环境恶化等因素对龙头鱼的影响,闽江口渔场龙头鱼群体极可能存在小型化、低龄化的趋势。由B-H模型分析可知,当前龙头鱼的开捕年龄较小,保持当前开发率不变,增大开捕体长至15.8 cm,即开捕年龄为1.220龄时,相对单位补充量渔获量与当前状态下同为0.025,且为最佳状态;由增加当前开发率至max,对应的值增大20%。二者比较可知,提高开捕体长有利于达到提高产量和保护资源的目的。
[1] BAILLY N, FROESE R, PAULY D, et al. Search FishBase [Z]. (2016). [2016-12-27]. https://www.fishbase. in/search.php.
[2] 朱元鼎,张春霖,成庆泰. 东海鱼类志[M]. 北京: 科学出版社, 1963: 132-133.
[3] 朱元鼎. 福建鱼类志[M]. 福州: 福建科学技术出版社, 1984: 174-175.
[4] 潘绪伟. 东海区龙头鱼渔业生物学的初步研究[D]. 上海: 上海海洋大学, 2011.
[5] 林显鹏,朱增军,李鹏飞. 东海区龙头鱼摄食习性的研究[J]. 海洋渔业, 2010, 32(3): 290-296.
[6] 郑元甲,陈雪忠,程家骅,等. 东海大陆架生物资源与环境[M]. 上海: 上海科学技术出版社, 2003: 260-278.
[7] 潘绪伟,程家骅. 长江口外海域龙头鱼营养生态学特征[J]. 中国水产科学, 2011, 18(5): 1132-1140.
[8] 林龙山. 东海区龙头鱼数量分布及其环境特征[J]. 上海海洋大学学报, 2009, 18(1): 66-71.
[9] 罗海舟. 龙头鱼主要生物学特性研究及资源状况分析[D]. 舟山: 浙江海洋学院, 2012.
[10] 陈玲,水柏年,董文霞. 龙头鱼生长特征及资源的可持续利用[J]. 中外企业家, 2012(6): 68-70.
[11] 晏磊,杨吝,杨炳忠,等. 南海北部龙头鱼刺网捕捞性能的初步研究[J]. 渔业信息与战略, 2013, 28(1): 33-38.
[12] NURUL AMIN S M. Studies on Age and Growth, VPA Analysis and Relative Condition Factor of(Ham-Buch) from the Neritic Water of Bangladesh[J]. Journal of Biological Sciences, 2001, 1(4): 192-194.
[13] GHOSH S, PILLAI N G K, DHOKIA H K. Fishery and population dynamics of Harpadon nehereus (Ham.) off the Saurashtra coast[J]. Indian Journal of Fisheries, 2009, 56(1): 13-19.
[14] 中华人民共和国国家质量监督检验检疫总局,中国国家标准化管理委员会. GB/T 12763.6–2007 海洋调查规范第6部分:海洋生物调查[S]. 北京: 标准出版社, 2007.
[15] 成庆泰,郑葆珊. 中国鱼类系统检索[M]. 北京: 科学出版社, 1987: 84.
[16] FROESE R. Cube law, condition factor and weight-length relationships: history, meta-analysis and recommendations[J]. Journal of Applied Ichthyology, 2006, 22(4): 241-253.
[17] 詹秉义. 渔业资源评估[M]. 北京: 中国农业出版社, 1995: 18-221.
[18] PAULY D, DAVID N. ELEFAN I, a basic program for the objective extraction of growth parameters from length-frequency data[J]. Berichte der Deutschen Wissenschaftlichen Kommission fur Meeresforschung, 1981, 28(4): 205-211.
[19] 王雪辉,邱永松,杜飞雁. 南海北部深水金线鱼生物学及最适开捕体长[J]. 应用生态学报, 2005, 16(12): 2428-2434.
[20] 陈国宝,李永振,陈丕茂,等. 鱼类最佳体长频率分析组距研究[J]. 中国水产科学, 2008, 15(4): 659-666.
[21] BERNARD D R. Multivariate Analysis as a Means of Comparing Growth in Fish[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1981, 38(2): 233-236.
[22] MUNRO J L, PAULY D. A simple method for comparing the growth of fishes and invertebrates[J]. ICLARM Fishbyte, 1983, 1(1): 5-6.
[23] PAULY D, MANILA M. Some simple methods for the assessment of tropical fish stocks[R]. No. 234. Rome: FAO Fisheries Technical Paper, 1983: 20-32.
[24] PAULY D. On the interrelationships between natural mortality, growth parameters and mean environmental temperature in 175 fish stocks[J]. Journal du Conseil international pour l’Exploration de la Mer, 1980, 39(2): 175-192.
[25] 何宝全,李辉权. 珠江河口棘头梅童鱼的资源评估[J]. 水产学报, 1988, 12(2): 125-134.
[26] BEVERTON, RAYMOND J H, HOLT, et al. On the Dynamics of Exploited Fish Populations[M]. London: Chapman & Hall/Crc Press, 1957: 1-20.
[27] PAULY D, SORIANO M L. Some practical extensions to Beverton and Holt’s relative yield-per-recruit model[C]. The First Asian Fisheries Forum. Maila: Asian Fisheries Society, 1986: 491-496.
[28] GAYANILO JR F C, SPARRE P, PAULY D. FAO-ICLARM stock assessment toolsⅡ(FisatⅡ): user’s guide (revised version) [R]. Rome: Food and Agriculture Organization of the United Nations, 2005: 1-154.
[29] 田辉伍,岳兴建,陈大庆,等. 怒江东方墨头鱼资源量和死亡参数的估算[J]. 生态学杂志, 2012, 31(1): 235-240.
[30] 何雄波,李军,沈忱,等. 闽江口主要渔获鱼类的生态位宽度与重叠[J]. 应用生态学报, 2018, 29(9): 3085-3092.
[31] 孙瑞林,陈志海. 浙江的龙头鱼定置刺网渔业[J]. 海洋渔业, 1986, 8(5): 215-216.
[32] 黄良敏. 闽江口和九龙江口及其邻近海域渔业资源现状与鱼类多样性[D]. 青岛: 中国海洋大学, 2011.
[33] 王家樵,黄良敏,李军,等. 闽江口及附近海域主要拖网鱼类的保护等级评价[J]. 海洋渔业, 2017, 39(5): 481-489.
[34] 陈潇,潘文斌,王牧. 福建闽江水口水库凤眼莲空间分布特征及其动态[J]. 湖泊科学, 2012, 24(3): 391-399.
[35] 叶章蕊,卢毅敏. 组合权重模糊联系度模型在水质评价中的应用[J]. 长江科学院院报, 2016, 33(9): 33-39.
[36] 周琦,池飞,逄勇,等. 可利用水资源量正逆向联合计算方法[J]. 水资源保护, 2016, 32(5): 42-46.
[37] 黄莹. 长门码头改建工程海域生态环境影响分析[J]. 福建质量管理, 2016(2): 65.
[38] 王恩辉. 我国典型潮间带沉积物中氮污染状况及浮游细菌群落对氮污染的响应[D]. 烟台: 中国科学院烟台海岸带研究所, 2017.
[39] 刘其根,沈建忠,陈马康,等. 天然经济鱼类小型化问题的研究进展[J]. 上海水产大学学报, 2005, 14(1): 79-83.
[40] 杜晓雪,高春霞,田思泉,等. 温台渔场龙头鱼生长、死亡及最适开捕规格研究[J]. 水产学报, 2018, 42(10): 1-15.
[41] GULLAND J A. Fish stock assessment: a manual of basic methods[M]. Volume 1. Chichester, UK: FAO/Wiley series on food and agriculture, 1983: 1-223.
[42] MEHANNA S F. Stock assessment and management of egyptian soleChabanaud, 1927 (Osteichthyes: Soleidae) in the southeastern Mediterranean, Egypt[J]. Turkish Journal of Zoology, 2007, 31(4): 379-388.
Estimation of Biological Parameters and Stock ofin the Min River Estuary, East China Sea
GUO Jun-hong1, LI Jun1, SHEN Chen1, SHI Yan1, FENG Chen1, HE Xiong-bo1, KANG Bin1,2
(1.,,361021,; 2.,266003,)
【】To investigate the resource ofin the Min River Estuary and estimate the biological parameters. 【】A total of 3 770 individuals ofwere sampled from 11 sites in the Min River Estuary in January, May, August and November 2015, respectively. The body length and weight of each individual sample were measured. The ELEFAN I method in FiSAT Ⅱ software was used to estimate the growth, mortality, optimum catchable size and yield-per-recruit of. 【】The Length()-Weight() relationship ofin the Min River Estuary was=0.00073.804 2(2=0.9611) and the parameter,L,mand0of Von Bertalanffy growth function was 0.52, 28.7 cm, 246.1 g and -0.317 years, respectively. Following Von Bertalanffy growth function, growth inflexion ofwas estimated, which showed the age of 2.252 years, body length 21.2 cm and weight of 77.1 g. The total mortality coefficient estimated with linear catch curve based on body length was 1.67; the natural mortality coefficient calculated by Pauly’s empirical equation was 1.04; the fishing mortality coefficient was 0.63 and the exploitation rate was equal to 0.377. The dynamic pool model suggested the age of capture was 0.712 years, suggesting the first catchable size was 11.9 cm. 【】The stock ofin the Min River Estuary has not been overexploited.
; Min River Estuary; biological parameters; exploitation rate; yield-per-recruit
S931.1
A
1673-9159(2019)05-0056-09
10.3969/j.issn.1673-9159.2019.05.009
2019-02-11
国家自然科学基金项目(41476149)
郭峻宏(1994―),男,硕士研究生,主要从事渔业资源与生态研究。E-mail:897959794@qq.com
康斌,男,教授,博士。E-mail:bkangfish@163.com
郭峻宏,李军,沈忱,等. 闽江口龙头鱼()的生物学参数及资源评估[J]. 广东海洋大学学报,2019,39(5):56-64.
(责任编辑:刘庆颖)
猜你喜欢
天天爱科学(2022年9期)2022-09-15 01:12:28
浙江海洋大学学报(自然科学版)(2022年4期)2022-08-18 02:40:50
科普童话·百科探秘(2022年8期)2022-05-30 10:48:04
昆明医科大学学报(2022年1期)2022-02-28 07:46:32
心声歌刊(2021年4期)2021-12-02 01:14:20
渔业研究(2021年3期)2021-06-29 02:07:24
闽都文化(2020年5期)2020-08-25 10:10:44
福建基础教育研究(2020年2期)2020-05-28 08:39:55
江苏农业科学(2017年10期)2017-07-21 17:12:51
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28 12:21:38
