
烤烟成熟期淀粉代谢关键酶活性与基因表达研究
2019-09-25 06:06潘飞龙田维强宋朝鹏郑明伟夏志林刘明宏芶剑渝李成刚
西北农林科技大学学报(自然科学版) 2019年9期
潘飞龙,田维强,宋朝鹏,郑明伟,夏志林,刘明宏,芶剑渝,李成刚
(1 河南农业大学 烟草学院,河南 郑州 450002;2 贵州省烟草公司遵义市公司,贵州 遵义 563000; 3 河南中烟工业有限责任公司,河南 郑州 450016)
烟草(NicotianatabacumL.)属茄科一年生或有限多年生草本植物,在我国南北各省广泛种植,是以收获叶片为目标的一种重要经济作物,其中烤烟种植范围最广,是卷烟工业的主要原料。烤烟是以积累淀粉为主的粉叶类植物[1],成熟鲜烟叶中淀粉含量可达40%[2],淀粉代谢作为烤烟最基本的初生代谢,直接影响着烟叶的基本生命活动[3]。烤烟成熟期烟叶淀粉的合成、积累、分解、转化状况,决定了烤后烟叶内部各化学成分之间的协调程度[4],进而影响烟叶的外观商品质量及内在品质[5]。因此,研究烤烟成熟期淀粉精细结构、淀粉代谢关键酶活性及基因表达模式,对探究烟叶品质形成作用机理及制定优质烟叶生产策略均有重要意义。
淀粉是烤烟的主要内含物质之一,成熟期烤烟淀粉代谢旺盛[18],其含量和比例最终影响烤后烟叶品质。虽然有关烤烟发育过程中碳水化合物积累和淀粉代谢调控方面的研究逐年增多,但迄今为止,对烤烟成熟期淀粉精细结构、淀粉代谢酶活性与基因表达模式系统性的研究却鲜有报道。本研究以不同淀粉积累型烤烟品种秦烟96和豫烟6号为材料,研究烤烟成熟期淀粉含量及比例、淀粉代谢关键酶活性与其基因表达的变化,探讨淀粉精细结构与淀粉代谢关键酶活性及其基因表达的关系,以阐明烤烟成熟期淀粉代谢的分子调控机制,为优质烟叶生产提供理论依据。
1 材料与方法
1.1 试验材料
试验于2017年在河南省洛阳市洛宁县小界卫洼烟叶标准化生产示范田进行。试验地位于东经111°38′,北纬34°26′,海拔1 114.6 m;该区属于暖温带大陆性季风气候,年平均气温13.7 ℃,日照2217.6 h,年降雨量600~800 mm。试验地土壤为黄棕壤,土壤基本理化性质:pH 7.26,有机质含量13.52 g/kg,碱解氮含量75.18 mg/kg,速效磷含量9.17 mg/kg,速效钾含量164.37 mg/kg。
选用当地主栽烤烟品种秦烟96和豫烟6号为试验材料。试验田内统一技术标准,全面落实漂浮育苗、平衡施肥、地膜覆盖、清棵培土、化学抑芽等技术措施,实施机械化作业、水肥一体化灌溉技术,其中水肥一体化采用烟叶水肥一体化自动注灌设备(贵州瑞欣现代农业有限公司)。烟苗于2017年5月8日移栽,收获时单株有效叶数18~20片,以培育“中棵烟”目标要求进行田间管理。烟田施氮量为72 kg/hm2,施用肥料为烟草专用复合肥(云南云叶化肥股份有限公司),m(N)∶m(P2O5)∶m(K2O)=1∶1∶2。烟叶于移栽后65 d打顶,打顶后30 d中部叶进入适熟阶段。
1.2 试验设计
试验采用随机区组设计,重复3次,共36个小区,小区面积16.5 m2,行株距1.1 m×0.5 m,每个试验小区植烟30株。试验设置6个取样时期处理,烟叶打顶当天即移栽后65 d开始取样,随后每隔10 d取样1次,共取样6次。取样时选取小区内长势均匀一致的烟株,每个品种各6株,选取大小均匀一致的中部叶(10~12叶位),去除主脉,选取第6~7支脉叶肉组织,混合样品,每份样品约10 g,置于液氮中速冻,然后于-80 ℃冰箱中保存,用于淀粉含量、淀粉代谢关键酶活性和基因表达分析。每个时期进行3次重复。
1.3 测定项目及方法
1.3.1 淀粉含量测定 烟叶中直链淀粉和支链淀粉含量测定参照吴玉萍等[19]的方法,采用AA3型连续流动化学分析仪(德国BRAN+LUEBBE公司生产)测定。采用液氮研磨新鲜烟叶,用乙醇-饱和氯化钠溶液进行脱色处理,随后用40%高氯酸溶液提取,直链淀粉和支链淀粉分别在600 nm和570 nm波长下显色,其中直链淀粉采用在线氢氧化钠碱液分两次进样,支链淀粉直接进样显色,根据标准曲线计算直链淀粉和支链淀粉含量。
1.3.2 淀粉代谢关键酶活性测定 1,6-二磷酸腺苷葡萄糖焦磷酸化酶(ADP-glucose pyrophosphorylase,AGPase)、颗粒结合型淀粉合成酶(granule-bound starch synthase,GBSS)、可溶性淀粉合成酶(soluble starch synthase,SSS)、淀粉分支酶(starch branching enzyme,SBE)、淀粉去分支酶(debranching enzyme,DBE)、α-淀粉酶(α-amylase)和β-淀粉酶(β-amylase)活性分别按照AGPase、GBSS、SSS、SBE、DBE、α-淀粉酶和β-淀粉酶试剂盒说明书(苏州科铭生物技术有限公司)测定。
1.3.3 淀粉代谢关键酶基因表达分析 采用改良CTAB法[20]提取烟叶样品总RNA,通过随机引物法反转录合成cDNA[21]。从GenBank核酸数据库中检索烟草AGPase、GBSS、SSS、SBE、DBE、α-amylase和β-amylase序列,利用Roche LCPDS2设计引物,见表1,其中烟草核糖体蛋白基因L25作为内参基因。

表1 淀粉代谢相关基因qRT-PCR引物序列Table 1 Primers for qRT-PCR of starch metabolism related genes
使用HiScript® Ⅱ qRT SuperMix Ⅱ试剂配制反应体系于PCR仪(ABI9700)中合成第一链cDNA。按照Invitrogen公司的Real Master Mix (SYBR Green)试剂盒操作指导,采用实时荧光定量PCR(qRT-PCR)的方法检测基因的相对表达量。反应程序为:95 ℃预热5 min,95 ℃变性10 s,60 ℃退火30 s,循环40次。每个样品均设置3次重复。采用2-ΔΔCt算法[22]对试验结果进行分析。
由评审小组主席主持,先由每一个课题的评审人报分数,如果大家的评分一致都很差,如几个人都给了5分以上,就直接不讨论(ND)而结束该课题评审;对于分数较好或意见不一致的近半数申请书会做进一步的充分讨论,并给予定量评分。讨论分两个层面,先是评审人讨论,由第一评审人简要介绍课题并陈述给分理由,另外几个评委补充陈述;第二个层面由所有参会人员讨论,每人都有资格发表意见,完毕后主持人请该课题评审人重新打分。
1.4 数据处理
试验数据采用Microsoft Excel 2010进行整理,采用Origin 9.1进行绘图,采用SPSS 22.0统计软件进行Pearson相关性分析及单因素方差分析,单因素方差分析采用新复极差法,显著性水平P<0.05,极显著性水平P<0.01。
2 结果与分析
2.1 烤烟成熟期烟叶淀粉含量的变化
烤烟成熟期烟叶淀粉组分含量及比例的变化见图1。
如图1-a所示,秦烟96和豫烟6号在整个烤烟成熟期烟叶淀粉含量呈先升高后降低的变化趋势,烟叶打顶(移栽后65 d)后,叶片内淀粉快速积累,两个品种烟叶淀粉含量均于移栽后85 d达到高峰,其淀粉含量分别为317.4和285.6 mg/g;从移栽后75 d开始,秦烟96烟叶淀粉含量显著高于豫烟6号。与烟叶淀粉积累规律相似,烟叶内直链淀粉和支链淀粉含量也呈先升高后降低的单峰波动变化(图1-b、c),移栽后85 d之前,秦烟96叶片内直链淀粉含量显著或极显著高于豫烟6号,此后两个品种烟叶直链淀粉含量差异不显著;秦烟96烟叶支链淀粉开始积累至达到峰值的时间长于豫烟6号,其支链淀粉含量于移栽后95 d达到高峰。移栽后85 d之前,豫烟6号烟叶支链淀粉与直链淀粉的比值高于秦烟96(图1-d);移栽后95 d,秦烟96 烟叶支链淀粉与直链淀粉的比值快速升高,显著高于豫烟6号烟叶支链淀粉与直链淀粉的比值,此后其比值波动下降。
2.2 烤烟成熟期淀粉代谢关键酶活性的变化
秦烟96烟叶中AGPase活性从移栽后65 d开始快速升高,至移栽后85 d达到高峰,之后快速下降;豫烟6号烟叶中AGPase活性变化趋势与秦烟96相似,但其活性低于秦烟96(图2-a)。两个品种烤烟GBSS Ⅰ活性在整个烤烟成熟期均较高,随着烤烟的成熟GBSS Ⅰ活性呈先升高后降低的变化趋势,且在成熟期差异不显著(图2-b)。
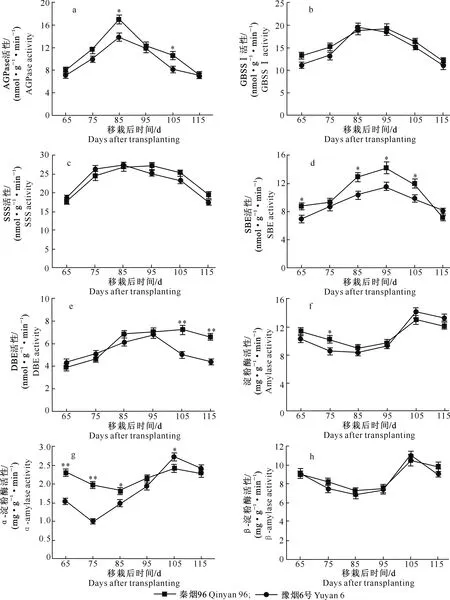
图2 烤烟成熟期淀粉代谢关键酶活性的变化Fig.2 Changes of key enzyme activities in starch metabolism of flue-cured tobacco at mature stage
如图2-c所示,烤烟成熟期烟叶SSS活性呈先升高后降低的变化趋势,但两个品种烤烟SSS活性达到峰值的时间略有不同,豫烟6号SSS活性峰值时间早于秦烟96。在整个烤烟成熟期,秦烟96和豫烟6号烟叶SBE活性变化趋势虽然一致,呈单峰波动变化,但秦烟96烟叶SBE活性显著高于豫烟6号;移栽后95 d,秦烟96和豫烟6号SBE活性达到最高,分别为14.23和11.57 nmol/(g·min)(图2-d)。秦烟96和豫烟6号烟叶DBE活性在移栽后95 d之前变化趋势相似,DBE活性随着烤烟的成熟逐渐升高;但移栽后95 d之后,两者DBE活性差异较大,秦烟96仍保持较高的DBE活性,呈波动缓慢降低,而豫烟6号烟叶DBE活性则快速降低(图2-e)。
在整个烤烟成熟期,秦烟96和豫烟6号烟叶淀粉酶活性呈先降低后升高再降低的波动变化趋势,移栽后95 d之后,其淀粉酶活性快速升高,至移栽后105 d达到峰值,随后淀粉酶活性略有降低(图2-f)。虽然在烤烟成熟期两个品种烤烟α-淀粉酶活性变化趋势相似,但与秦烟96相比,豫烟6号烟叶α-淀粉酶活性波动较大;移栽后105 d,秦烟96和豫烟6号烟叶α-淀粉酶活性达到最高,分别为2.42和2.73 mg/(g·min)(图2-g)。秦烟96和豫烟6号烟叶β-淀粉酶活性变化趋势与淀粉酶活性一致,移栽后105 d,两者β-淀粉酶活性达到峰值,分别为12.98和14.13 mg/(g·min)(图2-h)。
2.3 烤烟成熟期淀粉代谢关键酶基因表达量的变化
如图3所示,通过对秦烟96和豫烟6号成熟期烟叶淀粉代谢相关基因(NtAGPase、NtGBSSⅠ、NtSSS、NtSBE、NtDBE、Ntα-amylase、Ntβ-amylase)表达量的分析发现,在烟叶整个成熟阶段,NtAGPase基因表达量呈先升高后降低的变化趋势,秦烟96和豫烟6号在移栽后85~95 d具有较高的NtAGPase基因表达量,之后其表达量快速下调。从移栽后65 d开始,NtGBSSⅠ基因表达量逐渐上调,至移栽后95 d,其表达量达到最高,豫烟6号NtGBSSⅠ基因表达量显著高于秦烟96。
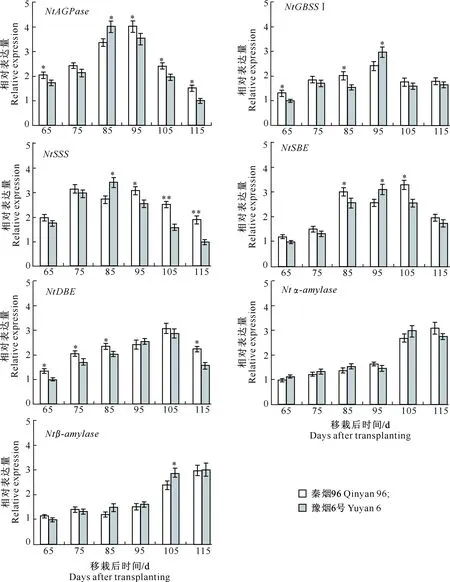
图3 烤烟成熟期淀粉代谢关键酶基因相对表达量的变化Fig.3 Changes of expression levels of key enzyme genes in starch metabolism in flue-cured tobacco at mature stage
秦烟96烟叶NtSSS基因表达量在成熟期呈双峰波动变化,移栽后75 d其表达量达到第一个峰值;而豫烟6号NtSSS基因表达量变化趋势与之不同,NtSSS基因表达量呈先升高后降低的单峰变化趋势,移栽后85 d其表达量达到最高(图3)。烤烟成熟期NtSBE基因表达量变化趋势与NtSSS基因表达量一致,但其达到峰值的时间略有不同。在整个烤烟成熟期,秦烟96和豫烟6号NtDBE基因表达量先升高后降低,移栽后105 d,NtDBE基因表达量达到最高。
移栽后65~95 d,秦烟96和豫烟6号烟叶Ntα-amylase基因表达量较低,移栽后95 d之后,其表达量显著上调,并保持较高的表达水平(图3)。烟叶中Ntβ-amylase基因表达量变化规律与Ntα-amylase基因表达量相似,其基因表达量也于移栽后95 d之后快速上调。
2.4 烤烟成熟期淀粉代谢关键酶活性及其基因表达量与淀粉含量的相关性
由表2可知,烤烟打顶至适熟阶段,烟叶中AGPase、GBSS Ⅰ、SSS、SBE和DBE活性与烟叶淀粉含量之间达显著或极显著正相关水平;AGPase、GBSS Ⅰ与烟叶直链淀粉含量之间达极显著正相关水平;SSS、SBE、DBE与烟叶支链淀粉含量之间达显著或极显著正相关水平;SBE、DBE与烟叶支链淀粉与直链淀粉的比值达显著正相关水平;而烟叶中淀粉酶、α-淀粉酶和β-淀粉酶活性则与烟叶中淀粉含量及比例相关性较弱。烟叶适熟至过熟阶段,烟叶中AGPase、GBSS Ⅰ、SSS、SBE和DBE活性与烟叶淀粉含量相关性减弱,而烟叶中淀粉酶(α-淀粉酶、β-淀粉酶)活性与烟叶淀粉含量及比例相关性显著增强,达显著或极显著负相关水平。
通过对烤烟成熟期淀粉代谢关键酶基因表达量与淀粉含量及比例之间相关系数的关联度分析,结果(表3)显示,烟叶打顶至适熟阶段,所有淀粉代谢关键酶基因表达量与烟叶淀粉含量及比例呈正相关,但其相关性与淀粉代谢关键酶活性相比波动较大,且大部分相关性减弱。烟叶适熟至过熟阶段,Ntα-amylase和Ntβ-amylase基因表达量与烟叶淀粉含量及比例相关性显著增强,达显著或极显著负相关水平;NtAGPase、NtGBSSⅠ和NtSSS基因表达量与烟叶支链淀粉与直链淀粉的比值达显著或极显著正相关水平。
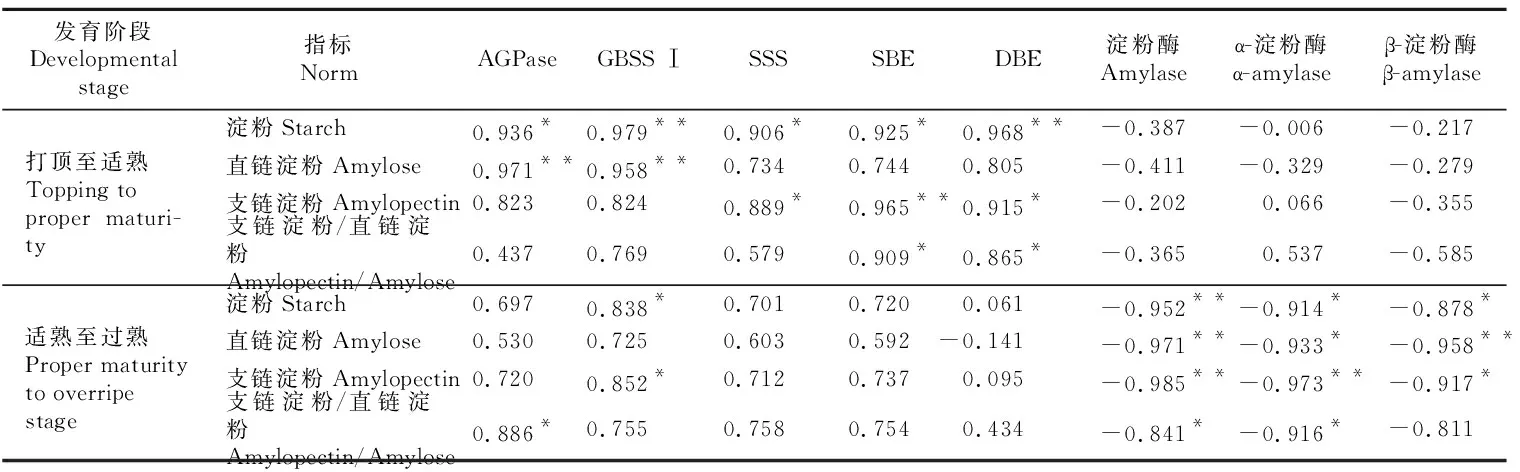
表2 烤烟成熟期淀粉代谢关键酶活性与淀粉含量的相关性Table 2 Correlation between key enzyme activities of starch metabolism and starch content in flue-cured tobacco at mature stage
注:打顶至适熟.移栽后65~95 d;适熟至过熟.移栽后95~115 d。*和**分别表示在P<0.05和P<0.01水平上显著。下同。
Note:Topping to proper maturity.65-95 days after transplanting;Proper maturity to overripe stage.95-115 days after transplanting.* and ** indicate significant difference atP<0.05 andP<0.01.The same below.
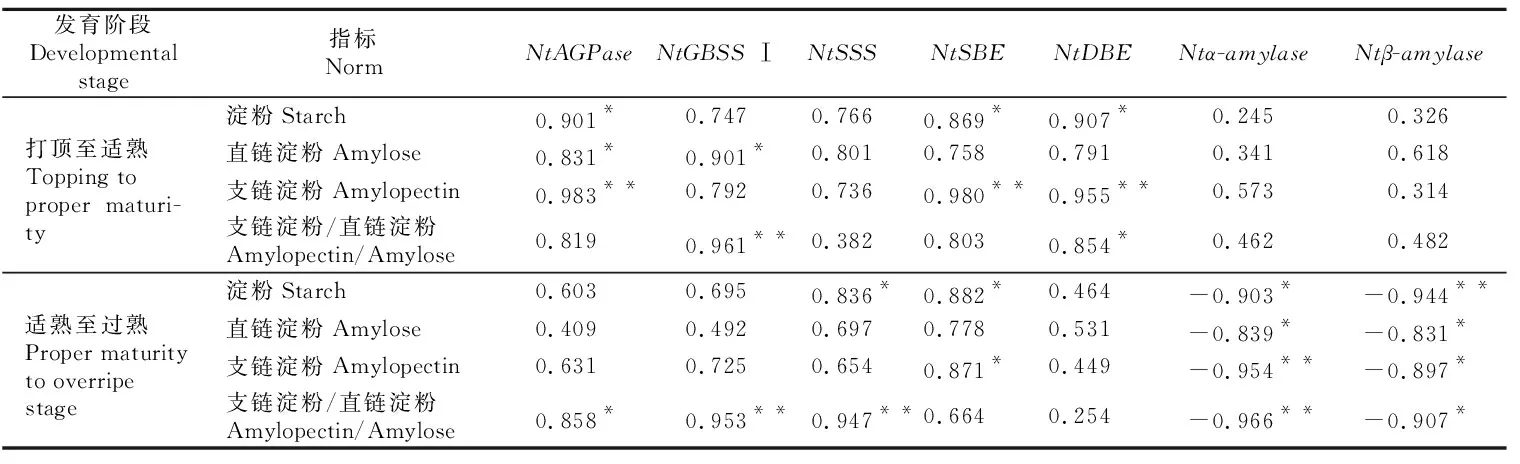
表3 烤烟成熟期淀粉代谢关键酶基因相对表达量与淀粉含量的相关性Table 3 Correlation between starch metabolism key enzyme activities and gene expression in flue-cured tobacco at maturity
如图4所示,分析了烤烟打顶至适熟、适熟至过熟发育阶段淀粉代谢关键酶活性与各相关基因表达量之间的相关性,结果表明,烟叶打顶至适熟阶段,大部分烟叶淀粉代谢关键酶基因表达量与各相关酶活性呈正相关水平,淀粉酶和β-淀粉酶活性与各相关基因表达量呈负相关。烟叶适熟至过熟阶段,烟叶中AGPase、GBSS Ⅰ、SSS、SBE和DBE活性与各相关基因表达量之间相关性减弱,而淀粉酶、α-淀粉酶和β-淀粉酶活性与各相关基因表达量之间相关性显著增强,其中Ntα-amylase和Ntβ-amylase基因表达量与淀粉酶(α-淀粉酶、β-淀粉酶)活性达显著或极显著正相关。
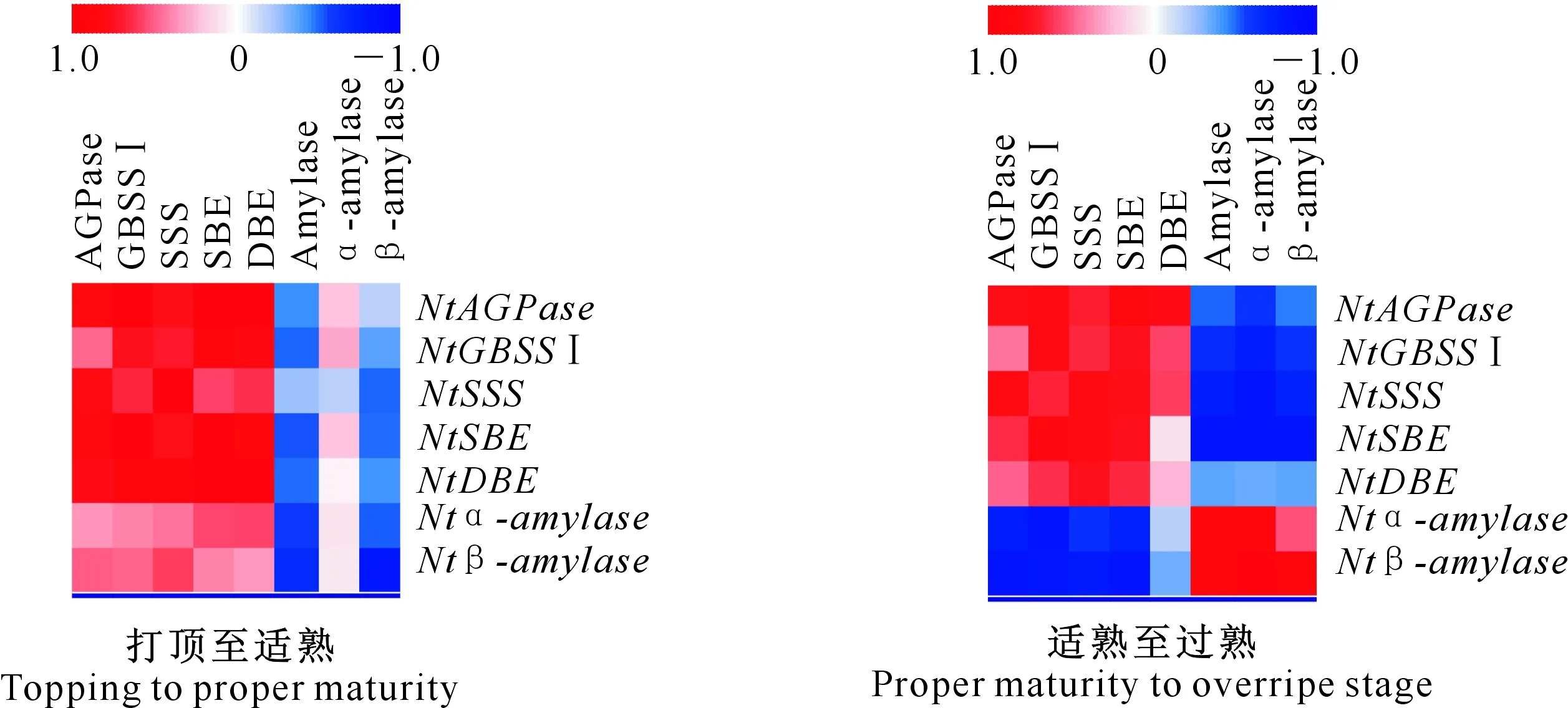
图4 烤烟成熟期淀粉代谢关键酶活性与基因相对表达量的相关性Fig.4 Correlation between key enzyme activities of starch metabolism and gene expression in flue-cured tobacco at mature stage
3 讨 论
烤烟成熟期烟叶淀粉快速合成、积累,代谢旺盛。牛德新等[23]研究表明,随着烟叶的成熟,烟叶淀粉含量呈上升趋势。杨胜男等[17]研究表明,烟叶成熟期淀粉含量逐渐上升,至烟叶生理成熟期达到最高,随后淀粉含量下降;直链淀粉和支链淀粉含量变化趋势与总淀粉含量变化一致,但支链淀粉含量达到峰值的时间则提前。本研究结果表现出相似的变化规律,秦烟96和豫烟6号从移栽后65 d淀粉快速积累,标志着烟叶生长进入成熟阶段;移栽后85 d淀粉含量达到峰值,说明此时烟叶为生理成熟期,淀粉含量最高,当烟叶适熟(移栽后95 d)时,两个品种烟叶淀粉含量稍有降低,但秦烟96淀粉含量显著高于豫烟6号,说明不同基因型烤烟淀粉代谢存在一定差异[24],秦烟96烟叶淀粉代谢活动高于豫烟6号。通过对成熟期烟叶淀粉组成比例研究发现,烟叶淀粉中支链淀粉含量显著高于直链淀粉,直链淀粉和支链淀粉含量变化趋势与总淀粉含量变化相同,呈先升高后降低的单峰波动变化,烟叶生理成熟期直链淀粉和支链淀粉含量最高,烟叶适熟时支链淀粉所占比例最高,这与前人的研究不同[13],这可能与烤烟品种及田间管理措施有关,有待进一步研究。成熟期烤烟积累大量淀粉,其中大部分淀粉在烟叶烘烤变黄期降解,而直链淀粉含量所占比例越高,烟叶淀粉易形成结晶区,烟叶淀粉不易降解,烤后烟叶淀粉含量较高,烟叶品质降低。
植物果实发育期间淀粉积累强度与淀粉代谢相关酶活性紧密相连[25],AGPase是高等植物淀粉生物合成第一个关键调节酶[26],催化G-1-P和ATP生成淀粉前体物质,其活性大小决定淀粉合成速率及淀粉合成量的多少[27]。大量研究表明,抑制AGPase活性可导致淀粉生物合成部分或完全停止,而AGPase基因过表达可提高转基因作物种子质量及淀粉含量[28]。本研究发现,从移栽后65 d开始,烟叶AGPase活性逐渐增强,烟叶生理成熟期(移栽后85 d)时AGPase活性最高,此时烟叶淀粉含量最高;烟叶工艺成熟期(移栽后95 d)AGPase活性显著降低,相应的烟叶淀粉含量呈不同程度降低,但支链淀粉与直链淀粉的比值显著升高,表明烟叶直链淀粉降解速率高于支链淀粉,这可能与SBE、DBE等酶活性有关;与豫烟6号相比,秦烟96具有更高的AGPase活性及淀粉含量,这与Petreikov等[29]在番茄上的研究结果一致。GBSS是直链淀粉合成的关键酶[30],它通过α-1,4-D-糖苷键将ADPG中葡萄糖残基与葡聚糖非还原端相连接,形成线性大分子。本研究发现,烤烟成熟期GBSS Ⅰ活性与直链淀粉含量变化趋势一致,进一步证实了GBSS Ⅰ对直链淀粉合成的重要性。
SSS与GBSS功能相同,通过α-1,4-D-糖苷键将ADPG中葡萄糖残基与葡聚糖非还原端相连接,但其主要存在于质体基质中,与SBE共同参与直链淀粉的合成[31]。SBE又称Q酶,它不仅能水解α-1,4-糖苷键连接的葡聚糖链,还能将切下的短链通过α-1,6-糖苷键与受体链连接,形成支链淀粉的分支结构[32]。有研究认为DBE对支链淀粉分支进行修饰,最终合成具有一定晶体结构的淀粉结晶体[33]。Nakamura等[34]认为,SBE与DBE活性之间的动态平衡对支链淀粉α-1,4-侧链的长度分配起着决定性作用。本研究结果发现,在整个烤烟成熟发育期,SSS、SBE和DBE活性均呈先升高后降低的单峰变化趋势,自移栽后65 d,三者酶活性快速增强,烟叶支链淀粉含量升高;烟叶工艺成熟期(移栽后95 d),秦烟96具有更高的SSS和SBE活性,相应的秦烟96具有更高的支链淀粉含量及比例,这与前人研究结果一致。淀粉降解主要通过淀粉酶(α-淀粉酶、β-淀粉酶)进行。本研究发现,自烟叶移栽后95 d达到工艺成熟,烟叶淀粉酶活性逐渐升高,淀粉降解,说明烟叶逐渐进入过熟阶段,淀粉代谢逐渐减弱。
淀粉代谢相关基因参与调控烟叶的淀粉代谢[35]。本研究发现,秦烟96和豫烟6号NtAGPase基因表达量在成熟期呈先升高后降低的变化趋势,移栽后85~95 dNtAGPase基因表达量较高,之后快速下调,这与贾宏昉等[16]的研究结果一致。烟叶中NtGBSSⅠ基因表达量与GBSS Ⅰ活性相似,移栽后95 d豫烟6号具有较高的NtGBSSⅠ基因表达量,但其直链淀粉含量少于秦烟96,这可能是因为秦烟96成熟前期直链淀粉积累较多。成熟期秦烟96烟叶NtSSS基因表达量呈双峰波动变化,而豫烟6号NtSSS基因表达量则呈单峰变化,NtSBE基因表达量变化趋势与NtSSS基因表达量一致,而两个品种烤烟NtDBE基因表达量则呈先升高后降低的变化趋势;秦烟96在烟叶成熟期NtSSS、NtSBE和NtDBE基因表达量高于豫烟6号,这可能是导致秦烟96烟叶支链淀粉含量高于豫烟6号的主要原因,这与杨胜男等[17]的研究结果一致。移栽后105 d,秦烟96和豫烟6号烟叶Ntα-amylase和Ntβ-amylase基因表达量显著上调,说明烟叶生长进入衰老阶段,淀粉代谢减弱。
综合分析烤烟成熟期打顶至适熟、适熟至过熟发育阶段淀粉含量及比例、淀粉代谢关键酶活性与基因表达量之间的关联性可知,烟叶打顶至适熟阶段,AGPase、GBSS Ⅰ、SSS、SBE和DBE活性与烟叶淀粉含量之间达显著或极显著正相关水平,其中AGPase、GBSS Ⅰ与烟叶直链淀粉含量达极显著正相关水平,SSS、SBE、DBE与烟叶支链淀粉含量之间达显著或极显著正相关水平,这进一步证实了GBSS Ⅰ是直链淀粉合成的关键酶,SSS、SBE和DBE是支链淀粉合成关键酶;SBE、DBE与烟叶中支链淀粉与直链淀粉的比值达显著正相关水平,说明SBE和DBE是调控烟叶支链淀粉与直链淀粉比例的关键酶。烟叶适熟至过熟阶段,AGPase、GBSS Ⅰ、SSS、SBE和DBE活性与烟叶淀粉含量相关性减弱,而淀粉酶(α-淀粉酶、β-淀粉酶)活性则与烟叶淀粉含量及比例相关性显著增强,说明当烟叶进入过熟阶段后,对烟叶淀粉代谢起主要调控作用的是淀粉酶(α-淀粉酶、β-淀粉酶)。烤烟成熟期淀粉代谢相关基因与淀粉含量及比例相关性略低于淀粉代谢关键酶活性与淀粉含量及比例相关性,说明淀粉代谢关键酶直接参与烟叶淀粉代谢的调控,而淀粉代谢相关基因则是在分子层面对烟叶淀粉代谢进行调控。
综上所述,烤烟成熟期经历了复杂的生理生化变化过程,淀粉代谢是一个受多种酶、多种基因调控的代谢活动。此外,淀粉代谢产生的糖类往往作为信号分子,与激素、氮等信号协同调控淀粉代谢和基因表达,由此进一步表明烟叶淀粉代谢的复杂性。
4 结 论
烤烟碳水化合物积累以淀粉为主,在烤烟发育成熟期,烟叶淀粉、直链淀粉和支链淀粉含量呈先升高后降低的变化趋势,烟叶生理成熟期其含量最高。烟叶打顶至适熟阶段,AGPase和GBSS Ⅰ对烟叶直链淀粉的积累贡献最大,NtGBSS Ⅰ对直链淀粉代谢起主要调控作用;SSS、SBE和DBE活性对烟叶支链淀粉积累起重要作用,NtSBE和NtDBE对烟叶支链淀粉代谢起主要调控作用。当烟叶过熟时,淀粉酶(α-淀粉酶、β-淀粉酶)主要参与烟叶的淀粉代谢活动。同一生态环境和栽培条件下,秦烟96淀粉积累较多,具有较高的支链淀粉比例,SBE和DBE活性的差异可能是造成烟叶直链淀粉与支链淀粉比例不同的重要原因。
猜你喜欢
食品与生物技术学报(2021年5期)2021-01-16
房地产导刊(2020年9期)2020-10-28
中国粮油学报(2019年4期)2019-07-12
天然产物研究与开发(2018年6期)2018-07-09
浙江柑橘(2016年4期)2016-03-11
中国果业信息(2015年9期)2015-01-22
北京航空航天大学学报(2014年1期)2014-12-19
植物营养与肥料学报(2014年1期)2014-03-11
中国机械工程(2012年15期)2012-07-25
中国科技信息(2010年20期)2010-11-07
