
湖南省吉首市发现利安德水鼩
2019-09-23 07:57张佩玲黄太福吴涛瞿勇张佑祥刘志霄
四川动物 2019年5期
张佩玲, 黄太福, 吴涛, 瞿勇, 张佑祥, 刘志霄
(吉首大学生物资源与环境科学学院,湖南吉首416000)
2017年5月25日,在湖南省吉首市马颈坳镇雾儿村进行小型哺乳动物调查时,于林间地面(109.815 239 0°E,28.383 342 7°N,海拔341 m)捡到1号食虫类标本(编号:201752501JS)。基于形态特征和分子数据,将其鉴定为劳亚食虫目Eulipotyphla鼩鼱科Soricidae东方水鼩属Chimarrogale的利安德水鼩Chimarrogaleleander(Thomas, 1902)。该种在我国分布于浙江、福建、广东、广西、江苏、江西、贵州、台湾、陕西、湖北(罗蓉,1993;Smith,解焱,2009;Yuanetal.,2013;蒋志刚等,2015;Abramovetal.,2017;刘少英等,2018),而湖南省未见记录,故本次发现应属湖南省哺乳类1新记录种。标本浸泡在95%酒精中,保存于吉首大学生物资源与环境科学学院动物标本室。
1 外形及头骨特征
该标本体型中等,体质量17.4 g,头体长87.96 mm,尾长(65.2 mm)小于头体长,后足(23.54 mm)较大。全身毛发短密细腻,在身体两侧和臀部有长的白色芒毛,且后者分布更为集中,比喜马拉雅水鼩Chimarrogalehimalayica的臀部芒毛更浓密。身体背腹面异色,分界不明显,背部毛基灰色,毛尖黑色;腹部毛基灰色,毛尖黑灰色。鼻部和吻部毛色深黑。前后足腹面黑褐色,深于背面。趾的两侧均有白色梳状栉毛,栉毛毛尖白色,毛基黑色且形成黑色线条。尾背面黑色,腹面前1/3白色,后2/3黑色。尾基部的毛发明显短于尾中后部,尾尖端有长的栉毛(图1)。外耳耳廓短小,隐于毛下,耳廓上长有毛发。
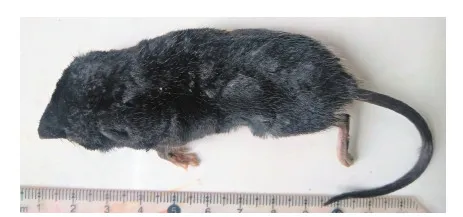
图1 利安德水鼩的外形特征 (黄太福 摄)
头骨外观狭长,颅全长25.29 mm,颅基长23.78 mm,眶间距5.47 mm,脑颅宽13.02 mm。颅骨骨壁薄,脑颅扁平,矢状嵴和人字嵴明显,从吻端到人字嵴的连线与齿槽线近乎平行。第1上门齿前尖向下垂直,有一小后尖,第2、3上门齿和上颌第一枚单尖齿基部约等大、尖等高;上前臼齿前尖低矮、后尖宽大尖锐且后缘呈圆弧状;第1上臼齿比第2上臼齿大,呈“W”状,第3上臼齿退化变小(图2)。上臼齿列长7.32 mm,下臼齿列长8.33 mm。齿式3.1.1.3/1.1.1.3=28。

图2 利安德水鼩的头骨特征 (张佑祥 摄)
A.头骨背面观dorsal view of skull; B. 头骨腹面观ventral view of skull; C. 头骨侧面观lateral view of skull; D. 下颌骨侧面观lateral view of mandible; E. 下颌骨正面观front view of mandible
2 分子分析
异丙醇法提取DNA,使用通用引物L14724_hk3和H15915_hk3(Heetal.,2010)进行PCR扩增,委托生工生物工程(上海)股份有限公司完成产物测序,获得的线粒体cytb基因序列长度为1 140 bp。从GenBank下载利安德水鼩在中国不同分布省区的序列(包括模式产地福建)和其亲缘关系较近种的序列用于数据分析(Yuanetal.,2013;Abramovetal.,2017)。使用MEGA 7.0(Kumaretal.,2016)以邻接法(NJ)构建系统发育树,显示利安德水鼩为单系,内部分为3个支系(分别为“台湾”“福建”和“陕西+湖北+湖南”)(图3)。基于Kimura 2-parameter模型计算序列间的遗传距离(Kimura,1980),显示“陕西+湖北+湖南”和“福建”支系之间的遗传距离为5.7%,“陕西+湖北+湖南”和“台湾”的为8.2%,“福建”和“台湾”的为7.0%。
3 讨论
在前人的研究中,利安德水鼩的分类归属经历了多次变化:最初,Thomas(1902)依据采集于我国福建挂墩的标本将其命名为Chimarrogaleleander;Allen(1938)在研究中国和蒙古国的哺乳动物时将其作为喜马拉雅水鼩福建亚种C.himalayicaleander记载;而Ellerman和Morrison-Scott(1951)将其归并于C.platycephalaleander;Harrison(1958)重新恢复了C.himalayica种的地位;但是之后又被修正为C.h.leander,并长期以C.h.leander被记载(Hoffmann,1987;罗蓉,1993;陈卫,2002;Smith,解焱,2009;郑生武,2010)。Yuan等(2013)基于系统发育和生物地理分析恢复了利安德水鼩种的地位,且由于遗传距离较大而将“台湾”标本和“陕西+湖北”标本分别作为假定种1和假定种2;蒋志刚等(2015,2016)在整理中国哺乳动物名录及脊椎动物红色名录时将其作为独立种记载;Abramov(2017)在修订东方水鼩属Chimarrogale的分类系统时,认为Yuan等(2013)划分的类群过细,而将“台湾”标本和“陕西+湖北”标本重新归于利安德水鼩;刘少英等(2018)认为我国中部和东部以及台湾分布的喜马拉雅水鼩亚种C.himalayicaleander属于独立种,即利安德水鼩。

图3 基于线粒体cyt b基因构建的邻接树
当前,我国分布的东方水鼩属物种为3种:利安德水鼩、喜马拉雅水鼩C.himalayica和灰腹水鼩C.styani(蒋志刚等,2015,2016;Abramovetal.,2017),三者之间的差异主要在于体型和毛色(罗蓉1993;Motokawaetal.,2006;Abramovetal.,2017)。罗蓉(1993)描述的喜马拉雅水鼩福建亚种C.himalayicaleander尾基部1/2的腹面白色,尾背色和尾尖毛色一致,为黑色;背部长有白色针毛,臀部更为浓密,趾两侧有白色栉毛等特征,与我们获得的标本特征基本一致,同时分子数据也支持湖南标本属于利安德水鼩。在最新出版的《中国兽类图鉴》(刘少英等,2018)中,提到喜马拉雅水鼩可能只分布在喜马拉雅山脉南麓,即西藏南部、云南和广西等地,而此前被认为是喜马拉雅水鼩的分布范围(中国中部和东部),则很可能分布的是利安德水鼩,本研究初步支持了该观点。同时,系统发育树和遗传距离的结果与Yuan等(2013)和Abramov等(2017)的研究一致,显示利安德水鼩分为3个支系且支系间的遗传距离较大,并且支持3个支系标本间的地理跨度大,隔离明显,然而是否已形成了种或亚种的分化还有待采集标本从多方面(如多基因甚至基因组、染色体核型和电镜扫描等)深入研究予以厘清。
猜你喜欢
环球时报(2022-11-10)2022-11-10
科普童话·百科探秘(2020年9期)2020-09-06
奥秘(2020年7期)2020-08-02
好日子(2019年4期)2019-05-11
神州民俗(2018年9期)2018-11-21
流行色(2017年1期)2017-05-31
新西部·中旬刊(2016年5期)2016-06-08
名作欣赏(2014年29期)2014-02-28
艺术评论(2013年12期)2013-11-10
意林(2010年19期)2010-10-22
