
不同贮藏温度下大菱鲆品质变化及货架期预测模型的建立
2019-09-18 02:06刘锋,梅俊,谢晶
渔业现代化 2019年4期
刘 锋,梅 俊,谢 晶
(上海水产品加工及贮藏工程技术研究中心,上海冷链装备性能与节能评价专业技术服务平台,食品科学与工程国家级实验教学示范中心(上海海洋大学),上海海洋大学食品学院,上海 201306)
大菱鲆(Scophthalmusmaximus)盛产于大西洋东侧沿岸,是名贵的低温经济鱼类[1]。大菱鲆因其蛋白含量高、营养丰富、味道鲜美等特点为消费者喜爱。自1992年从国外引进大菱鲆养殖以来,经过数十年的发展,大菱鲆已成为中国北方沿海养殖的主要品种之一[2]。随着大菱鲆产业逐渐复苏,如何有效保持其在贮藏过程中的品质特性,延长货架期,扩大销售范围、满足商超生鲜商品的供应需求,正日益引起人们的高度关注[3]。贮藏温度是决定水产品货架期的关键因素,不同温度下水产品的品质和货架期差异显著[4]。目前,关于水产品货架期预测模型的研究主要为零级和一级动力学模型。雷志方等[5]建立了不同温度条件下的金枪鱼货架期预测模型,动力学分析表明零级动力学模型比一级动力学模型更符合金枪鱼各项指标的变化。姜元欣等[6]通过对不同冻藏温度下罗非鱼品质变化的研究,确定挥发性盐基总氮(TVB-N)为敏感指标,并建立了恒定冷链温度下冻藏罗非鱼的货架期模型。Li等[7]基于菌落总数、TVB-N以及硫代巴比妥酸值(TBA),利用Arrhenius方程建立了不同贮藏温度下真空包装罗非鱼的动力学预测模型。崔正翠等[8]基于希瓦氏菌的生长情况利用修正的Gompertz方程建立了0 ℃~10 ℃下贮藏大菱鲆特定腐败菌希瓦氏菌的生长动力学和货架期预测模型。
本研究以感官、挥发性盐基总氮(TVB-N)、菌落总数、硫代巴比妥酸值、电导率、肌肉的微观结构变化、水分迁移状况为指标,研究大菱鲆贮藏在-3 ℃、0 ℃、4 ℃、10 ℃和15 ℃条件下的品质变化,应用动力学模型和Arrhenius方程建立挥发性盐基总氮(TVB-N)及菌落总数随贮藏温度和时间变化的货架期预测模型,确定货架期与不同温度之间的相关性,为大菱鲆保鲜技术的优化以及货架期的实时监测提供参考。
1 材料与方法
1.1 材料与试剂
新鲜养殖大菱鲆购于上海市浦东新区芦潮港水产批发市场,规格基本一致(600±50)g,活鱼装入加水的充氧袋中,立即运回实验室处理。平板计数琼脂(PCA)、三氯乙酸、MgO、甲基红、溴甲酚绿、硫代巴比妥酸、无水乙醇、二甲苯、福尔马林、苏木精、伊红,购于生工生物工程(上海)股份有限公司。
1.2 仪器与设备
BPS-100CB恒温恒湿箱(上海一恒科学仪器有限公司);低场核磁共振成像分析仪(上海纽迈电子科技有限公司);FOSS凯氏定氮仪(瑞典FOSS公司);WFZ UV-2100型紫外可见分光光度计(尤尼柯(上海)仪器有限公司);高温高压灭菌锅(HVE-50,日本Hi-rayama制造有限公司);Nikon-E200 光学显微镜(日本Nikon公司);RM2235 冷冻切片机(德国Leica公司)。
1.3 试验方法
1.3.1 样品处理
将新鲜的大菱鲆用蒸馏水冲洗,去腮和内脏,待水沥干后以整条鱼密封在聚乙烯保鲜袋中,分别于-3 ℃、0 ℃、4 ℃、10 ℃、15 ℃条件下贮藏,15 ℃每12 h取样,10 ℃每1 d取样,4 ℃每2 d取样,0 ℃每3 d取样,-3 ℃每4 d取样,感官评分达到腐败时停止取样,测定并记录不同指标数据的变化情况。
1.3.2 感官评定
参考GB 2733—2015 鲜、冻动物性水产品[9]及张彩丽等[10]对海水鱼感官评定方法,稍作修改。选取6个专业人员组成感官评定小组,从色泽、气味、组织形态和弹性4个方面对大菱鲆品质进行评分,总分值16~20为新鲜,9~15为品质良好,8分以下为腐败变质。具体评分标准见表1。
1.3.3 TVB-N、电导率和TBA的测定
TVB-N测定参照凡玉杰等[11]的方法;电导率测定参考胡玥等[12]的方法;TBA参考马超锋等[13]的方法测定。

表1 大菱鲆感官特性检验评分标准
1.3.4 菌落总数和持水力 (WHC)的测定
菌落总数按照GB 4789.2—2016[14]测定。持水力参照HAN[15]等的方法测定。
1.3.5 水分迁移状况的研究
参考王尊等[16]的方法,略作修改。将切好的鱼块(2 cm×2 cm×1 cm)用保鲜膜包裹,放入测定管中,采用核磁共振成像分析仪测定鱼样T2驰豫时间,设置T2测量参数:采样频率(SW)=100 kHz,模拟增益(RG1)=20 dB,硬脉冲90°脉宽(P1)=18 μs,数字增益(DRG1)=3 dB,采样点数(TD)=200 020,重复采样等待时间(TW)=5 000 ms,累加次数(NS)=8,硬脉冲180°脉宽(P2)=37 μs,回波时间(TE)=0.4 ms,回波个数(NECH)=5 000。根据CPMG指数衰减曲线图,用分析软件进行迭代反演得到横向弛豫时间T2图谱。将同样的鱼块再次放入测定管中,用核磁共振成像仪进行成像,得到质子密度图后对图像统一映射和伪彩。
1.3.6 背部肌肉微观组织观察
参照李婷婷等[17]的方法,将大菱鲆的背部肌肉切成5 mm×5 mm×5 mm的小块,置于5%的福尔马林溶液中固定24 h,乙醇梯度脱水后,经二甲苯透明处理、石蜡包埋,制成1 cm3的石蜡块,然后用Leica切片机切成10 μm厚的薄片,HE常规染色后在光学显微镜下(10×10倍)观察切片组织。大菱鲆屠宰后取新鲜鱼样,不同贮藏温度的样品分别在贮藏末期取样(15 ℃ 3 d,10 ℃ 5 d,4 ℃ 11 d,0 ℃ 15 d,-3 ℃ 20 d)。
1.3.7 大菱鲆货架期预测模型的建立
(1)动力学模型。在评价水产品的品质变化时,通常采用一级反应动力学方程进行模拟预测。
B=B0ekt
(1)
式中:B0为测定品质指标的初始值;k为反应速率常数;t为贮藏时间,d;B为贮藏第t天时品质指标的测定值。
(2) Arrhenius方程。Arrhenius方程反应温度T与反应速率常数k之间的关系,方程如下:
(2)
式中:k为反应速率常数;A0为指前因子;Ea为活化能,J/mol;R为气体常数,8.314 510 J/(mol·K);T为绝对温度,K。 将270 K、273 K、277 K、283 K和288 K温度下的反应速率常数k结合Arrhenius方程进行拟合得到活化能Ea和指前因子A0。
(3)货架期模型的建立。结合动力学模型和Arrhenius方程得到大菱鲆的货架期模型:
(3)
式中:tSL为货架期,d。
1.4 数据分析
以上每个指标做3次平行实验,利用Excel绘制图表,采用Origin8.5软件绘制折线图,并使用SPSS22.0对试验数据进行相关性分析和显著性分析,P<0.01表示显著差异。
2 结果与分析
2.1 感官评价
由图1可知,随着贮藏时间的延长,各组大菱鲆的感官品质均下降。这是由于大菱鲆在贮藏过程中酶活性的化学反应以及微生物大量繁殖的共同作用所致,15 ℃和10 ℃的样品感官下降迅速,分别在第3天和第6天达到感官不可接受程度;4 ℃和0 ℃在贮藏前期差异不显著(P>0.01),贮藏10 d后感官下降速率增大,4 ℃和0 ℃贮藏分别于第13天和第16天超出可接受范围;-3 ℃贮藏的大菱鲆时间最长,可达24 d,这是由于微冻环境抑制了微生物的繁殖以及酶活性的化学反应,大幅延长大菱鲆的货架期,但是在到达货架期终点时,0 ℃样品的感官品质要略好于-3 ℃。

图1 不同贮藏温度大菱鲆感官评分的变化
2.2 TVB-N的变化
挥发性盐基总氮是指水产品在贮藏期间受到微生物和酶的作用,蛋白质降解产生氮以及胺类等碱性挥发性物质,是国际上衡量水产品腐败变质的重要指标[18]。根据GB/T18108—2008《鲜海水鱼》[19]规定:海水鱼的TVB-N≤15 mg/100g,属于一级品;TVB-N≤20 mg/100g,属于二级品;TVB-N≤30 mg/100g为三级品。分析TVB-N值可以得出15 ℃、10 ℃、4 ℃、0 ℃和-3 ℃贮藏大菱鲆的货架期分别为2.5 d、5 d、12 d、15 d和24 d,可见低温可以抑制酶活性和微生物的繁殖,降低蛋白质的降解速度(图2)。
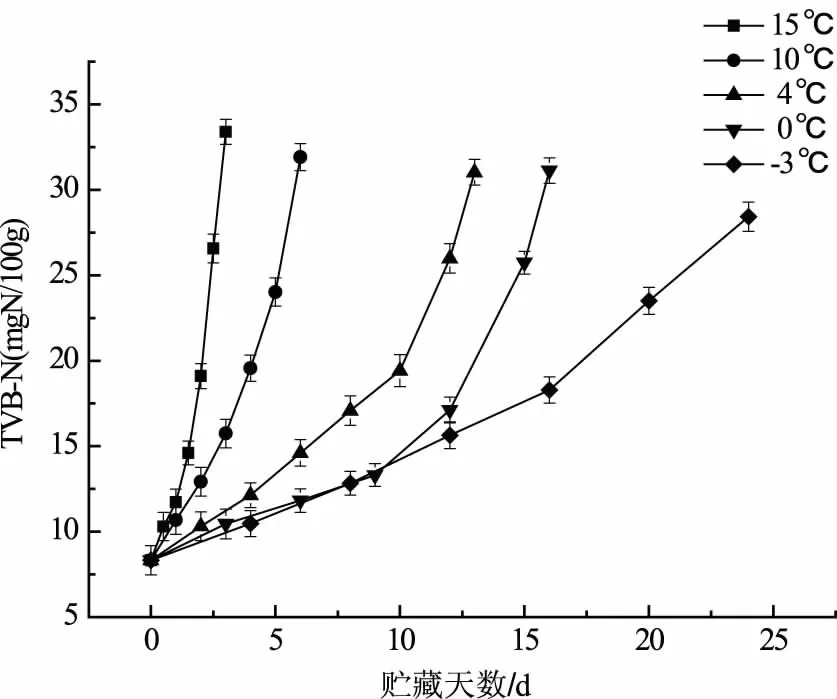
图2 不同贮藏温度下大菱鲆TVB-N的变化
2.3 电导率的变化
电导率能够反映物体的导电能力,是一种快速检测水产品新鲜度的方法[20]。由图3可知,新鲜大菱鲆的电导率为1 546 us/cm,贮藏在不同温度下的大菱鲆电导率均不断上升,这是因为在贮藏过程中,鱼体内的脂肪、蛋白质等大分子物质在外源性微生物蛋白酶的作用下分解成小分子物质,并以大量离子的形式存在,从而使溶液的导电能力增强。15 ℃和10 ℃贮藏的样品相比于4 ℃、0 ℃以及-3 ℃,电导率上升的速度要快得多。主要是因为温度升高,引起电解质的电离度增大,离子迁移速度增大,电导率也随之增加。随着贮藏时间的延长,大菱鲆鱼肉分解出来的产物越来越多,溶液导电能力越强,电导率不断增加,新鲜度也越差。实验表明,大菱鲆的电导率与贮藏温度紧密相关,降低温度可减缓其品质裂变进程,保证新鲜度。大菱鲆在15 ℃、10 ℃、4 ℃、0 ℃和-3 ℃贮藏温度下TVB-N与电导率的相关系数分别为0.968、0.978、0.957、0.912、0.951,呈显著相关(P<0.01),这与宋丽荣[21]等人对罗非鱼贮藏期间TVB-N与电导率相关性研究得出的结果相一致。

图3 不同贮藏温度下大菱鲆电导率的变化
2.4 菌落总数的测定
微生物是引起水产品腐败变质的重要因素之一,微生物的生长繁殖使鱼体内的蛋白质、氨基酸等物质降解,产生胺类、硫化氢等物质,发出具有腐败特征的臭味[22],因此,检测大菱鲆贮藏过程中的微生物动态变化能够很好地反映其品质特性。
大菱鲆在15 ℃、10 ℃、4 ℃、0 ℃和-3 ℃的微生物变化如图4所示,该图表明不同贮藏温度下大菱鲆菌落总数的变化差异显著(P<0.01)。大菱鲆菌落总数的初始值为3.85 log(CFU/g),随着贮藏时间的延长,各组菌落总数值不断上升,15 ℃和10 ℃微生物生长直接进入对数期,分别在第3 天和第6 天达到7.12 log(CFU/g)和7.06 log(CFU/g),超过了水产品微生物数量的最高可接受限度(7 log CFU/g)[23],4 ℃和0 ℃贮藏的微生物贮藏前期生长缓慢,这是因为微生物的新陈代谢需要酶的催化,低温抑制了酶的催化速率,分别在第12 天和第15 天接近可接受阈值,而-3 ℃的菌落数在初始阶段有所下降,可能是因为鱼体自身携带的一些细菌因不适应微冻环境受到抑制,直到贮藏末期仍未超过可接受阈值,结果表明-3 ℃可大大延长大菱鲆的货架期。
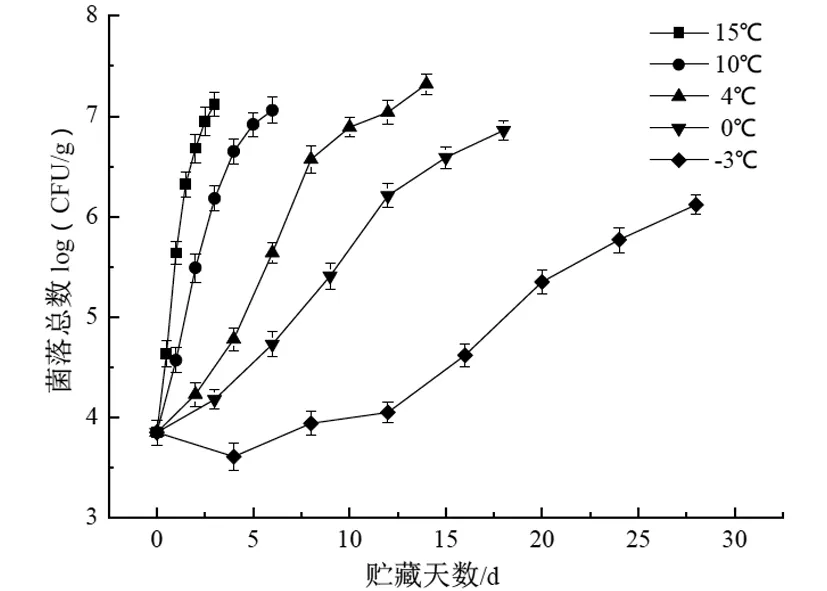
图4 不同贮藏温度下菌落总数的变化
2.5 持水力(WHC)的测定
由图5可知,5组大菱鲆的持水性不断下降,15 ℃和10 ℃环境下贮藏的持水力下降速度要远大于4 ℃、0 ℃、-3 ℃组的样品,可能是因为温度的升高导致蛋白质降解越来越严重,肌原纤维蛋白变性程度增大,结合水的能力下降,并且在贮藏前期,5组样品的持水性差异不明显,后期越来越显著(P<0.01),原因可能是贮藏时间的增加会加剧蛋白质的变性[24]。观察图4和图5发现,二者在表征大菱鲆鲜度上呈现负相关(P<0.01),15 ℃、10 ℃、4 ℃、0 ℃和-3 ℃贮藏温度下的菌落总数与持水力的相关系数分别为-0.906、-0.955、-0.983、-0.992和-0.965。

图5 不同贮藏温度下大菱鲆持水力的变化
2.6 TBA的变化
大菱鲆在贮藏过程中脂肪氧化产生丙二醛,丙二醛与硫代巴比妥酸反应生成稳定的红色络合物,通过测定该物质在532 nm处的吸光度判断大菱鲆的氧化程度[25]。脂肪氧化通常发生在贮藏的初始阶段。由图6可以看出,大菱鲆的TBA值在贮藏期间总体呈先上升后下降的趋势,主要是因为贮藏后期丙二醛与蛋白质、核酸等物质发生反应被分解,引起质量浓度降低。TBA的初始值为0.045 mg/kg,不同温度下贮藏末期TBA值均出现上升,表明大菱鲆在贮藏期间发生了脂质氧化,品质出现劣变。
2.7 鱼肉水分变化的 LF-NMR研究
鱼肉中的水分主要以不易流动水的形式存在[26],其在贮藏加工与销售过程中起重要作用,因此,本试验着重考察鱼肉在贮藏期间不易流动水横向弛豫时间T22和驰豫面积P22的变化(表2)。随着贮藏时间的延长,5种不同贮藏温度下鱼肉中不易流动水均呈下降趋势,总体来看,-3 ℃鱼片的T22下降速率相比于其他温度较为缓慢。T22值不断下降表明鱼肉在贮藏过程中水分子受到的约束力增大,水分活度减小,疏水性降低,可能是因为贮藏后期蛋白质变性程度变大,致使空间结构发生改变,部分不易流动水转化为自由水。在贮藏末期样品已经变质,蛋白质空间结构被严重破坏,水分迁移速率显著降低(P<0.01)。

图6 不同贮藏温度下大菱鲆TBA的变化
而所有组的驰豫面积P22在贮藏前期均呈上升趋势,可能是因为鱼体死后机体缺氧、细胞膜通透性增大、肌丝胀大、吸水能力增强所致[27]。P22值在贮藏后期下降,是因为长时间的贮藏使鱼体表面出现干耗,肌原纤维收缩将细胞内的水分挤压出来,不易流动水转化为自由水。各温度在贮藏末期P22含量存在差异,可见温度和时间都会影响鱼肉中不易流动水P22的变化。

表2 大菱鲆在不同贮藏温度下水分横向弛豫时间T22和驰豫面积P22的变化
图7表示大菱鲆在不同贮藏温度期间的核磁共振(MRI)成像图。一般而言,MRI图像中亮度越强,表明该部分的水分含量越高[28]。由图7可得,在贮藏过程中,不同贮藏温度下大菱鲆MRI图像的亮度逐渐降低,贮藏末期亮度基本消失,表明鱼样已发生腐败。15 ℃和10 ℃成像亮度的减弱程度大于4 ℃、0 ℃和-3 ℃,表明低温贮藏可以延缓鱼体的腐败,适宜用于保持鱼体原本的鲜度和鱼肉的品质,抑制鱼体死后出现的生物化学变化[29]。
2.8 肌肉组织微观结构的变化
图8显示大菱鲆鱼肉在不同贮藏温度下的横断面微观组织结构。图8a表明,新鲜大菱鲆鱼肉肌纤维结构紧密,轮廓清晰,肌纤维之间有一些微小的间隙;当15 ℃和10 ℃鱼样贮藏至第3 天和第5 天时(图8b和图8c),肌纤维受到挤压发生严重位移,且肌纤维断面边缘出现大量絮状物,轮廓模糊,肌纤维扭曲变形、断裂严重;当4 ℃和0 ℃贮藏至11 d和15 d后(图8d和图8e),两组的肌纤维均逐渐变形,不断发生断裂和小片化,肌节间隙增大,肌纤维结构由紧密变得疏松,这两组温度下的鱼样相比于15 ℃和10 ℃,细胞完整性保持较好;当-3 ℃贮藏至第20天时(图8f),与新鲜鱼样相比,肌纤维结构依然较完整,轮廓清晰,但肌纤维内部出现裂口损伤,肌节间隙增大,可能是因为微冻条件下细胞内部产生许多小冰晶,导致细胞受到机械损伤,肌纤维断裂。

图7 不同贮藏温度下大菱鲆核磁共振成像图的变化
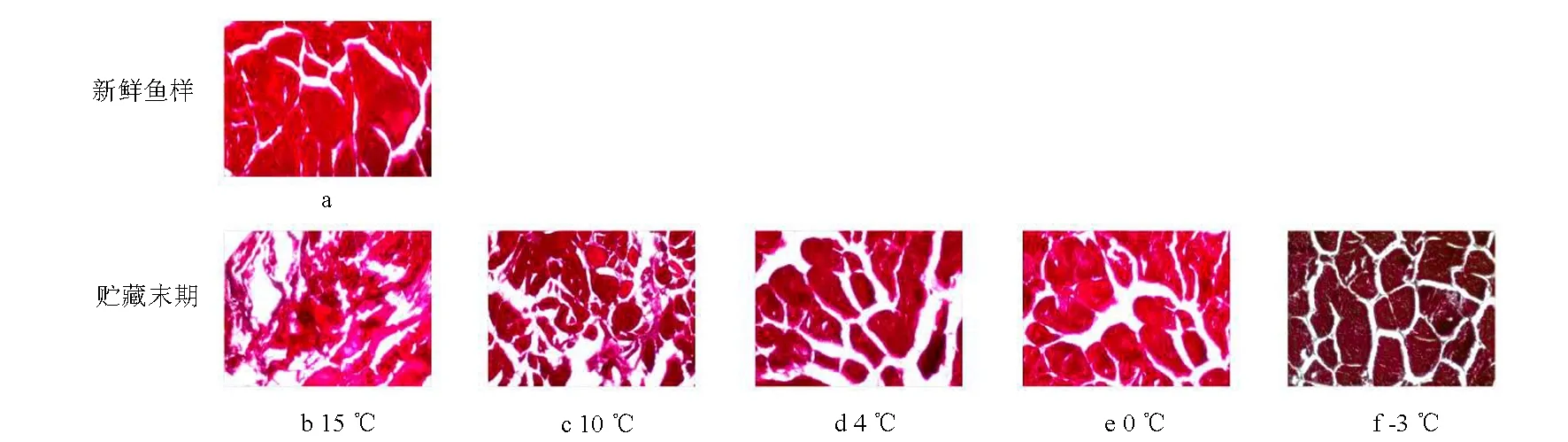
图8 不同温度下大菱鲆肌肉组织的微观结构变化
2.9 货架期预测模型的建立
2.9.1 大菱鲆品质动力学分析
根据公式(1),利用Origin 8.5分别对大菱鲆的TVB-N值和菌落总数值进行非线拟合,得到一级动力学模型的反应速率常数k及回归系数R2(表3),R2越大,表明拟合度越高,预测效果越好。

表3 不同贮藏温度下大菱鲆品质变化的动力学模型参数
2.9.2 Arrhenius方程中指前因子A0和活化能Ea的计算
根据式(2),对k和T作非线性拟合,计算可得大菱鲆TVB-N及菌落总数变化预测模型中的指前因子A0和活化能Ea(表4)。

表4 TVB-N和TVC变化预测模型中的指前因子A0和活化能Ea
将TVB-N及菌落总数的A0和Ea分别代入到式(3)中可得到对应的货架期预测模型。
大菱鲆基于TVB-N的货架期预测模型:
(4)
大菱鲆基于菌落总数的货架期预测模型:
(5)
式中:BTVB-N、BTVC为大菱鲆贮藏第t天时的TVB-N和菌落总数的测定值;BTVB-N0、BTVC0为TVB-N和菌落总数的初始值;tsL为货架期,d。
2.10 货架期模型的验证与评价
为了验证该动力学预测模型的准确性,比较283 K温度下大菱鲆TVB-N及菌落总数值的实际测量值与模拟预测值,结果见表5。
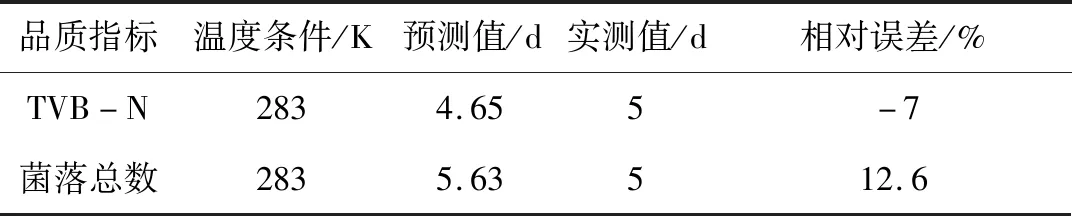
表5 283 K温度下大菱鲆的货架期预测误差
由表5可知,以TVB-N值建立的货架期预测模型,大菱鲆的实测值与货架期方程预测值相比,相对误差在±10%以内,而以菌落总数建立的货架期预测模型相对误差达到12.6%,因此从整体的预测效果来看,以TVB-N值建立的货架期预测模型能够较为准确地预测大菱鲆的货架期。
3 结论
研究了大菱鲆贮藏在不同温度下的品质变化,选取TVB-N值和菌落总数构建货架期预测模型。结果表明,降低温度可显著延缓各指标的变化速率,抑制产品劣变,从而延长大菱鲆的货架期。大菱鲆在贮藏过程中的TVB-N值和菌落总数符合一级动力学模型,通过一级动力学模型结合Arrhenius方程构建的货架期预测模型具有较高的拟合度,10 ℃指标实测值与模型预测值相对误差在10%以内,表明该模型可信度高。该模型只需测定产品的TVB-N值就可以得到产品的剩余货架期,操作快速准确。该模型可预测-3 ℃~15 ℃下大菱鲆的货架期,应用范围更广。因此,该货架期预测模型具有较高的实用性,可成为预测大菱鲆货架期的一种有效途径。
猜你喜欢
干旱地区农业研究(2022年3期)2022-05-24
河北渔业(2022年3期)2022-03-18
矿冶工程(2021年4期)2021-09-15
中国科技纵横(2020年20期)2020-11-28
世界有色金属(2020年15期)2020-10-10
销售与市场·管理版(2018年6期)2018-10-08
科学中国人(2018年1期)2018-06-08
农村百事通(2016年18期)2016-11-05
河北渔业(2016年5期)2016-09-08
河北渔业(2016年4期)2016-07-30
