
不同藻密度下诺氟沙星对萼花臂尾轮虫生命表统计学参数的影响
2019-09-17 11:07朱韩席贻龙徐秋磊黄克强李加男江珊
生态毒理学报 2019年3期
朱韩,席贻龙,徐秋磊,黄克强,李加男,江珊
安徽师范大学生命科学学院,安徽省重要生物资源的保护和利用研究省级重点实验室,皖江城市带退化生态系统的恢复与重建安徽省协同创新中心,芜湖 241000
中国是抗生素生产和使用的大国,年产抗生素原料21万t,人均年使用量高达138 g左右;抗生素还被大量地用于动物疾病的防治及动物饲料中[1]。喹诺酮类抗生素是近年来医用和兽用最普遍的抗生素。作为第三代喹诺酮的代表性药物,诺氟沙星(norfloxacin, NFLX)对革兰氏阴性菌和革兰氏阳性菌均具有很好的杀菌效果,因此被更广泛地使用。然而,包括诺氟沙星在内的绝大部分抗生素不能被动物和人体有效地吸收,约90%以原药或代谢产物的形式随病人和禽畜的排泄物进入环境[2]。通过淋洗作用进入水体中的诺氟沙星已被陆续检出。在黄海海域的水体中,诺氟沙星的含量可达108.8 ng·L-1[3];在枯水季节的珠江广州段,诺氟沙星含量在117~251 ng·L-1之间;在重庆某医院废水中,诺氟沙星含量高达1 620 ng·L-1[4]。水环境中的诺氟沙星等抗生素会通过饮水、农业灌溉和食品等对人类健康和生态系统安全造成威胁,所以有关抗生素对水生生物的生态毒性效应研究显得尤为重要。
有关诺氟沙星对水生动、植物的毒性作用已有一些报道,研究内容涉及诺氟沙星对浮萍(Lemnagibba)、鱼腥藻(AnabaenaCPB4337)、近头状伪蹄形藻(Pseudokirchneriellasubcapitata)和剑尾鱼(Xiphophorushellerii)等的急性毒性[5-7],诺氟沙星对蛋白核小球藻(Chlorellapyrenoidesa)、斜生栅藻(Scenedesmusobliquus)、大型溞(Daphniamagna)、中国对虾(Fenneropenaeuschinensis)、西伯利亚鲟鱼(Acipenserbaerii)和剑尾鱼(Xiphophorushellerii)等的慢性毒性[6,8-14]。但有关诺氟沙星对轮虫的毒性作用尚鲜有报道。轮虫是浮游动物中的一个重要类群。萼花臂尾轮虫(Brachionuscalyciflorus)等植食性轮虫以藻类、细菌和有机碎屑等为食物,同时又是晶囊轮虫、桡足类、水生昆虫和鱼类幼体等捕食者的食物,因此在水生态系统的物质循环和能量流动过程中具有重要的作用。轮虫具有分布广、繁殖速率快、世代时间短、个体小、易培养、对水环境变化敏感等特点,已被广泛地用作水生态毒理学研究的受试生物[15-16]。
已有研究表明,食物密度不仅影响轮虫的存活、生殖和种群增长,也影响污染物对轮虫的毒性[17-21]。自然水体中,藻类等的密度普遍具有时空变动的特点,因此研究不同食物密度下污染物对轮虫的毒性作用具有重要的意义。为此,本文采用生命表试验方法研究了3个藻密度下不同浓度的诺氟沙星对萼花臂尾轮虫生命表统计学参数的影响,旨在探究诺氟沙星毒性与藻密度之间的关系,以及各试验终点对诺氟沙星污染的相对敏感性。
1 材料与方法(Materials and methods)
1.1 轮虫的来源和培养
试验所用的萼花臂尾轮虫采于芜湖市镜湖,实验室内进行克隆培养。以EPA[22]作为培养液,以HB-4培养基[23]培养的、处于指数增长期的斜生栅藻为食物。实验前,在(25±1) ℃、自然光照的恒温培养箱内对萼花臂尾轮虫进行一周以上的预培养。预培养期间,每天定时投喂密度分别为1.0×106、2.0×106和4.0×106cells·mL-1的斜生栅藻并半数更换轮虫培养液一次,同时通过去除部分个体使轮虫种群始终处于指数增长状态。
1.2 测试液的配制
试验所用诺氟沙星(分子式C16H18FN3O3)由Solarbio公司生产,淡黄色粉末状,纯度大于98%。用电子天平称取200 mg诺氟沙星,用EPA培养液作为溶剂,配制浓度为1 000 mg·L-1的原液。诺氟沙星微溶于水,需用1 mol·L-1的NaOH进行助溶。配制时加入的NaOH体积以保证诺氟沙星刚好溶解为宜。待诺氟沙星溶解后,再用1 mol·L-1的HCl进行中和调节pH,以液体颜色由澄清变为浅黄为基准,再微调pH至7.5~8.5[12]。实验时再用EPA培养液将其稀释为所需浓度的测试液。每天所用诺氟沙星均新鲜配制以确保其质量。
1.3 急性毒性试验
为选择合适的慢性实验药物浓度,生命表实验之前,先进行24 h急性毒性实验。急性毒性实验前,在1.0×106、2.0×106和4.0×106cells·mL-1藻密度下进行勘探实验,然后根据勘探实验结果,将诺氟沙星浓度按等差间距设置为350、400、450、500、550和600 mg·L-16个浓度梯度,另设一个空白对照组;实验在8 mL玻璃杯中进行,每杯中放入10只龄长在4 h以内的轮虫幼体和5 mL测试液,每组设置3个重复。试验在(25±1) ℃、无光照的培养箱中进行,24 h后观察记录每个玻璃杯中轮虫的死亡数目,采用机率单位法获得24 h-LC50值。
1.4 慢性毒性试验
根据急性毒性试验所得到的各食物密度下的LC50值的1/8~1/100[24],将诺氟沙星浓度设置为5、20、35、50、65和80 mg·L-1。试验在1.0×106、2.0×106和4.0×106cells·mL-1等3个藻密度下进行,各藻密度下设置一个对照组,每组设置3个重复。试验时,以龄长在4 h以内的轮虫幼体作为受试母体。试验在8 mL玻璃杯中进行,每杯放置10只轮虫幼体和5 mL测试液(其中分别含有相应浓度的诺氟沙星及相应密度的斜生栅藻)。试验在(25±1) ℃、自然光照的恒温培养箱中进行。试验过程中,每隔12 h观察并记录母体的存活数和孵化出的幼体数,去除死亡母体,同时将孵化的幼体移至另一玻璃杯中继续培养,待其产卵后确定雌体类型,用于计算轮虫母体一生所产全部后代中的混交雌体比例;每隔24 h更换1次测试液(含有相应密度的斜生栅藻)。试验进行至轮虫母体全部死亡为止。
1.5 研究参数的定义与计算方法
1.6 数据统计和分析
采用SPSS 16.0分析软件对数据进行分析。采用Kaplan-Meier分析3个食物密度下诺氟沙星浓度对轮虫存活率的影响。对所得各数据作正态性检验后,对符合正态分布的各组数据通过单因素方差分析(one-way ANOVA)和多重比较(LSD检验)分析各浓度组与对照组间的差异显著性,通过双因素方差分析检测诺氟沙星浓度、斜生栅藻密度及其交互作用对轮虫各生命表统计学参数影响的显著性;对轮虫各生命表统计学参数与诺氟沙星浓度之间的关系进行回归分析。
2 结果(Results)
2.1 不同食物密度下诺氟沙星对轮虫的急性毒性
急性毒性试验结果表明,(25±1) ℃、无光照、斜生栅藻密度为1.0×106、2.0×106和4.0×106cells·mL-1时,诺氟沙星对萼花臂尾轮虫的24 h-LC50值分别是452.873、515.146和526.650 mg·L-1,其95%的置信区间分别为402.781~493.960、464.569~622.417和472.370~666.323 mg·L-1。
2.2 不同食物密度下诺氟沙星对轮虫存活率和繁殖率的影响
和对照组相比,各藻类食物密度下,暴露于各浓度诺氟沙星溶液中的萼花臂尾轮虫的存活率和和繁殖率均显著提高。当藻密度为1.0×106cells·mL-1时,暴露于35 mg·L-1诺氟沙星溶液中的轮虫繁殖率峰值最高,为5.53个后代·雌体-1·(12 h)-1;当藻密度为2.0×106cells·mL-1时,暴露于65 mg·L-1诺氟沙星溶液中的轮虫繁殖率峰值最高,为6.73个后代·雌体-1·(12 h)-1;当藻密度为4.0×106cells·mL-1时,暴露于20 mg·L-1诺氟沙星溶液中的轮虫繁殖率峰值最高,为7.13个后代·雌体-1·(12 h)-1(图1)。

图1 不同藻密度和诺氟沙星(NFLX)浓度下萼花臂尾轮虫的存活率和繁殖率注:图中数据表示为平均值±标准误。Fig. 1 Survivorship and fecundity of B. calyciflorus exposed to different concentrations of norfloxacin (NFLX) and cultured at different algal densities Note: data are expressed as Mean±SE.
2.3 不同食物密度下诺氟沙星对轮虫生命表统计参数的影响
统计分析结果表明,3个藻密度下,诺氟沙星浓度对轮虫的生命期望和世代时间均有显著性影响(P<0.01);与对照组相比,暴露于5~80 mg·L-1诺氟沙星溶液中轮虫的生命期望和世代时间均显著延长(P<0.01)。当藻密度为1.0×106cells·mL-1时,暴露于5 mg·L-1诺氟沙星溶液中轮虫的生命期望和世代时间均延长到对照组的1.4倍;暴露于35 mg·L-1诺氟沙星溶液中轮虫的生命期望和世代时间延长的幅度最大,均为对照组的2.1倍。当藻密度为2.0×106cells·mL-1时,暴露于5 mg·L-1诺氟沙星溶液中轮虫的生命期望和世代时间均延长到对照组的1.6倍;暴露于50 mg·L-1诺氟沙星溶液中轮虫的生命期望和世代时间延长的幅度最大,均为对照组的2.0倍。当藻密度为4.0×106cells·mL-1时,暴露于5 mg·L-1诺氟沙星溶液中轮虫的生命期望和世代时间均延长到对照组的1.6倍,而暴露于其他各浓度诺氟沙星溶液中轮虫的生命期望和世代时间延长的幅度低于藻密度为1.0×106和2.0×106cells·mL-1时。总体上,5 mg·L-1的诺氟沙星使生命期望和世代时间的延长幅度随着藻密度的升高而增大,但6个处理组的平均提高幅度却随着藻密度的升高而减小(1.0×106、2.0×106和4.0×106cells·mL-1藻密度下轮虫生命期望延长幅度的平均值依次为对照组的1.9倍、1.8倍和1.6倍,世代时间延长幅度的平均值依次为对照组的1.8倍、1.8倍和1.6倍)(表1)。
3个藻密度下,诺氟沙星浓度对轮虫的净生殖率和种群内禀增长率均具有显著性影响(P<0.01);与空白对照组相比,暴露于5~80 mg·L-1诺氟沙星溶液中轮虫的净生殖率和种群内禀增长率均显著提高(P<0.01)。当藻密度为1.0×106cells·mL-1时,暴露于5 mg·L-1诺氟沙星溶液中轮虫的净生殖率和种群内禀增长率分别为对照组的2.7倍和1.59倍;暴露于20~80 mg·L-1诺氟沙星溶液中轮虫的净生殖率和种群内禀增长率分别为对照组的5.2倍~5.9倍和1.83倍~2.12倍。当藻密度为2.0×106cells·mL-1时,暴露于5 mg·L-1诺氟沙星溶液中轮虫的净生殖率和种群内禀增长率分别为对照组的4.0倍和1.69倍,而暴露于20~80 mg·L-1诺氟沙星溶液中轮虫的净生殖率和种群内禀增长率提高的幅度(4.7倍~5.4倍和1.74倍~1.79倍)均低于藻密度为1.0×106cells·mL-1时。当藻密度为4.0×106cells·mL-1时,暴露于各浓度诺氟沙星溶液中轮虫的净生殖率和种群内禀增长率(4.5倍~6.6倍和1.94倍~2.56倍)提高的幅度均高于藻密度为1.0×106和2.0×106cells·mL-1时。总体上,5 mg·L-1的诺氟沙星对净生殖率和种群内禀增长率的提高幅度随着藻密度的升高而增大,但6个处理组的平均提高幅度却随着藻密度由1.0×106升高至2.0×106和4.0×106cells·mL-1时呈现出先降低后升高的趋势(藻密度由低到高时,轮虫净生殖率提高幅度的平均值依次为对照组的5.0倍、4.8倍和6.0倍,种群内禀增长率提高幅度的平均值依次为对照组的1.85倍、1.76倍和2.20倍)(表1)。
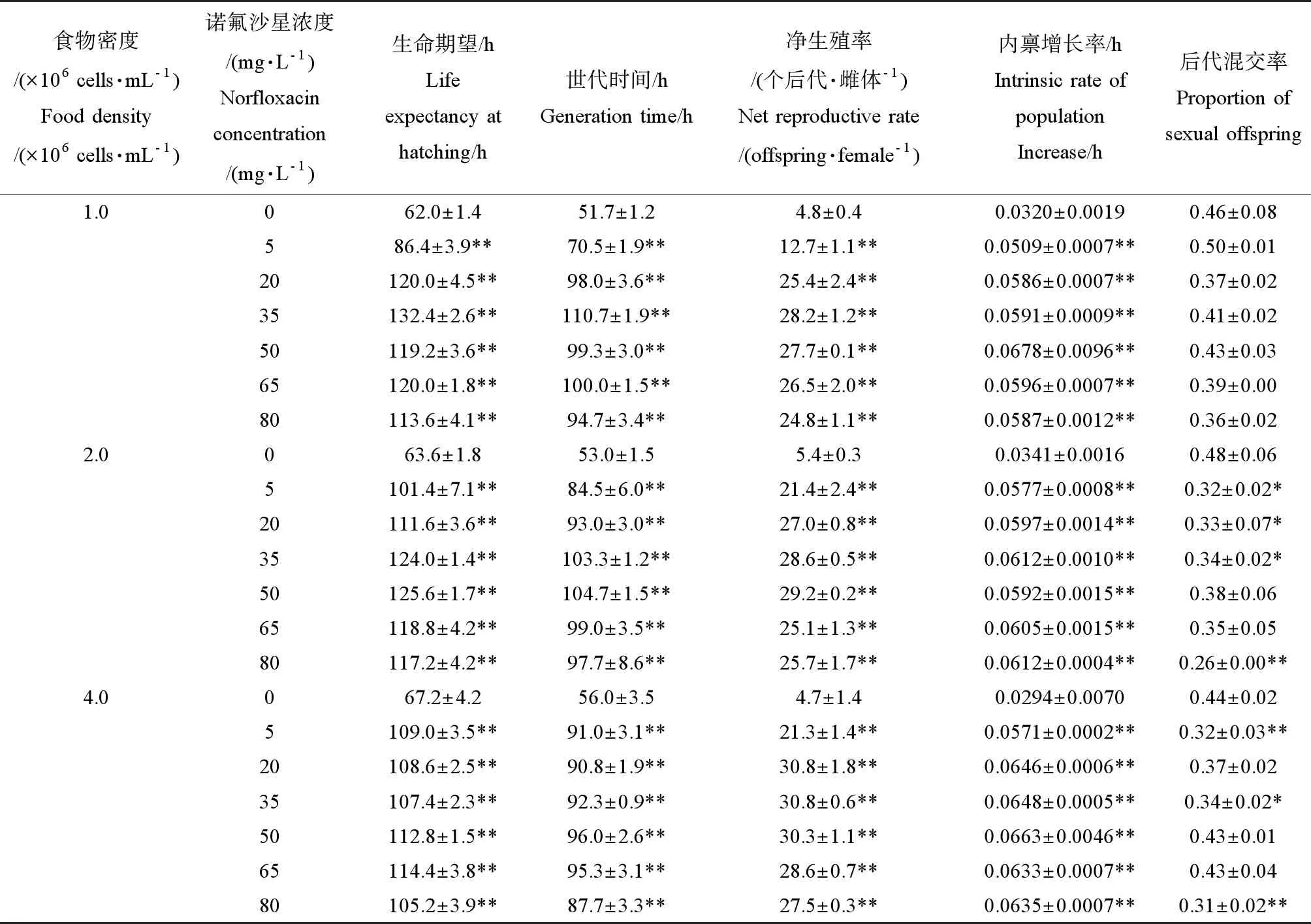
表1 不同藻密度和诺氟沙星浓度下萼花臂尾轮虫的生命表统计学参数Table 1 Life-table demographic parameters of B. calyciflorus exposed to different concentrations of norfloxacin and cultured at different algal densities
注:表中数据表示为平均值±标准误。和同一藻密度下的对照组相比有显著性差异(*P<0.05、**P<0.01)。
Note: data are expressed as Mean±SE. Compared with the control, *, ** means significantly (P<0.05 orP<0.01) different from the control at each algal density.
当藻密度为1.0×106cells·mL-1时,诺氟沙星浓度对轮虫的后代混交率无显著性影响(P>0.05);而当藻密度为2.0×106和4.0×106cells·mL-1时,诺氟沙星浓度对轮虫的后代混交率有显著性影响(P<0.05或P<0.01)。与空白对照组相比,当藻密度为2.0×106cells·mL-1时,暴露于5~35和80 mg·L-1诺氟沙星溶液中的轮虫后代混交率显著降低;当藻密度为4.0×106cells·mL-1时,暴露于5、35和80 mg·L-1诺氟沙星溶液中的轮虫混交率显著降低(表1)。
双因素方差分析结果表明,藻密度对轮虫的生命期望有显著性影响(P<0.05),对轮虫的净生殖率和后代混交率有显著性影响(P<0.01);诺氟沙星浓度对轮虫所有生命表统计参数都有显著性影响(P<0.01);藻密度和诺氟沙星浓度的交互作用对轮虫的生命期望和世代时间有显著性影响(P<0.01),对轮虫的净生殖率有显著性影响(P<0.05)(表2)。

表2 藻密度和诺氟沙星浓度对萼花臂尾轮虫生命表统计参数的影响(双因素方差分析)Table 2 Effects of algal density and norfloxacin (NFLX) concentration on life table demographic parameters of B. calyciflorus (Two-way ANOVA)
回归分析结果表明,当藻密度为1.0×106和2.0×106cells·mL-1时,诺氟沙星浓度与轮虫的生命期望、世代时间、净生殖率和内禀增长率之间具有显著的剂量-效应关系;当藻密度为4.0×106cells·mL-1时,诺氟沙星浓度与轮虫的净生殖率和内禀增长率之间具有显著的剂量-效应关系(表3)。
3 讨论(Discussion)
3.1 抗生素对萼花臂尾轮虫的毒性
在25 ℃、藻密度分别为1.0×106、2.0×106和4.0×106cells·mL-1的条件下,利福平对萼花臂尾轮虫的 24 h-LC50值分别是 18.0、25.0和19.7 mg·L-1[25],盐酸四环素对萼花臂尾轮虫的24 h-LC50值分别是107.95、116.92和111.23 mg·L-1[26]。本研究中,诺氟沙星对萼花臂尾轮虫的24 h-LC50值分别是452.873、515.146和526.650 mg·L-1。因此,与利福平和盐酸四环素相比,诺氟沙星对萼花臂尾轮虫的毒性最低。
3.2 抗生素浓度和藻密度对轮虫存活的影响
已有研究表明,暴露于5.6~2 000 mg·L-1链霉素、90~2 000 mg·L-1四环素和泰乐霉素以及50~200 μg·L-1阿莫西林溶液中的萼花臂尾轮虫平均寿命均显著缩短,暴露于90~ 2 000 mg·L-1链霉素、四环素和泰乐霉素溶液中的褶皱臂尾轮虫(B.plicatilis)平均寿命均显著缩短,暴露于100~200 μg·L-1阿莫西林溶液中的哈瓦那臂尾轮虫(B.havanaensis)平均寿命也显著缩短[27-28]。但也有研究结果显示,抗生素对萼花臂尾轮虫存活的影响取决于抗生素的种类和藻类食物密度[25-26]。本研究中,3个斜生栅藻密度下,暴露于5~80 mg·L-1诺氟沙星溶液中的萼花臂尾轮虫生命期望均显著延长(表4)。
3.3 抗生素浓度和藻密度对轮虫繁殖的影响
暴露于5.6 mg·L-1或更高浓度的链霉素和四环素、90 mg·L-1泰乐霉素以及50~200 μg·L-1阿莫西林溶液中的萼花臂尾轮虫净生殖率显著降低,暴露于50~200 μg·L-1阿莫西林溶液中的哈瓦那臂尾轮虫净生殖率显著降低,暴露于90 mg·L-1或更高浓度链霉素、四环素和泰乐霉素溶液中的褶皱臂尾轮虫净生殖率显著降低[27-28]。然而,近年来的研究结果显示,抗生素对轮虫净生殖率的影响也取决于抗生素的种类和藻类食物密度[25-26]。本研究中,3个斜生栅藻密度下,暴露于5~80 mg·L-1诺氟沙星溶液中的萼花臂尾轮虫净生殖率均显著升高(表4)。
通常情况下,单巢目轮虫主要通过孤雌生殖繁殖后代,此时非混交雌体产出非混交卵,该卵孵化出非混交雌体;但在一定的条件下非混交雌体产出混交卵,该卵孵化出混交雌体从而进行有性生殖[29]。有关污染物对轮虫后代混交率的影响已有一些报道,结果显示一定浓度的、具有雌激素活性的水体污染物,如雌二醇、保幼激素、睾丸素、邻苯二甲酸酯类物质和Zn2+等,均显著提高了轮虫的后代混交率[30-33]。就抗生素的影响而言,已有研究结果表明,在1.0×106、2.0×106和4.0×106cells·mL-1的斜生栅藻密度下,暴露于2.0~10.0 mg·L-1具雌激素效应的利福平溶液中的萼花臂尾轮虫后代混交率均显著升高[25],盐酸四环素浓度也对萼花臂尾轮虫的后代混交率具有显著的影响[26]。与上述研究结果不同的是,本研究发现,在1.0×106cells·mL-1的斜生栅藻密度下,诺氟沙星浓度对萼花臂尾轮虫的后代混交率无显著性影响;而在2.0×106和4.0×106cells·mL-1的斜生栅藻密度下,诺氟沙星浓度却对萼花臂尾轮虫的后代混交率具有显著的影响。看来,欲揭示抗生素对轮虫混交率影响的总体规律,需要对更大浓度范围的、更多种类的抗生素进行测试。
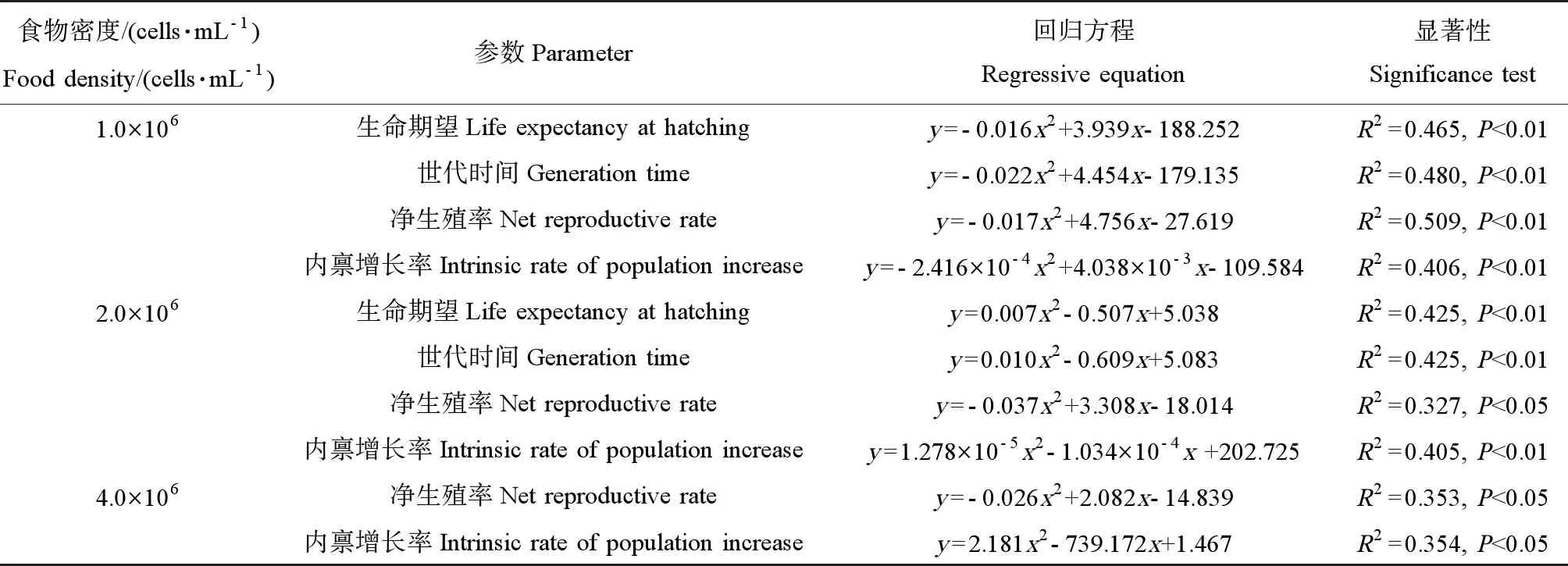
表3 各藻密度下萼花臂尾轮虫部分生命表统计学参数与诺氟沙星浓度间的关系Table 3 The relationships between some life-table demographic parameters of B. calyciflorus cultured at different algal densities and norfloxacin concentration
注:x为诺氟沙星浓度,单位为mg·L-1。
Note:xis norfloxacin concentration, and the unit is mg·L-1.
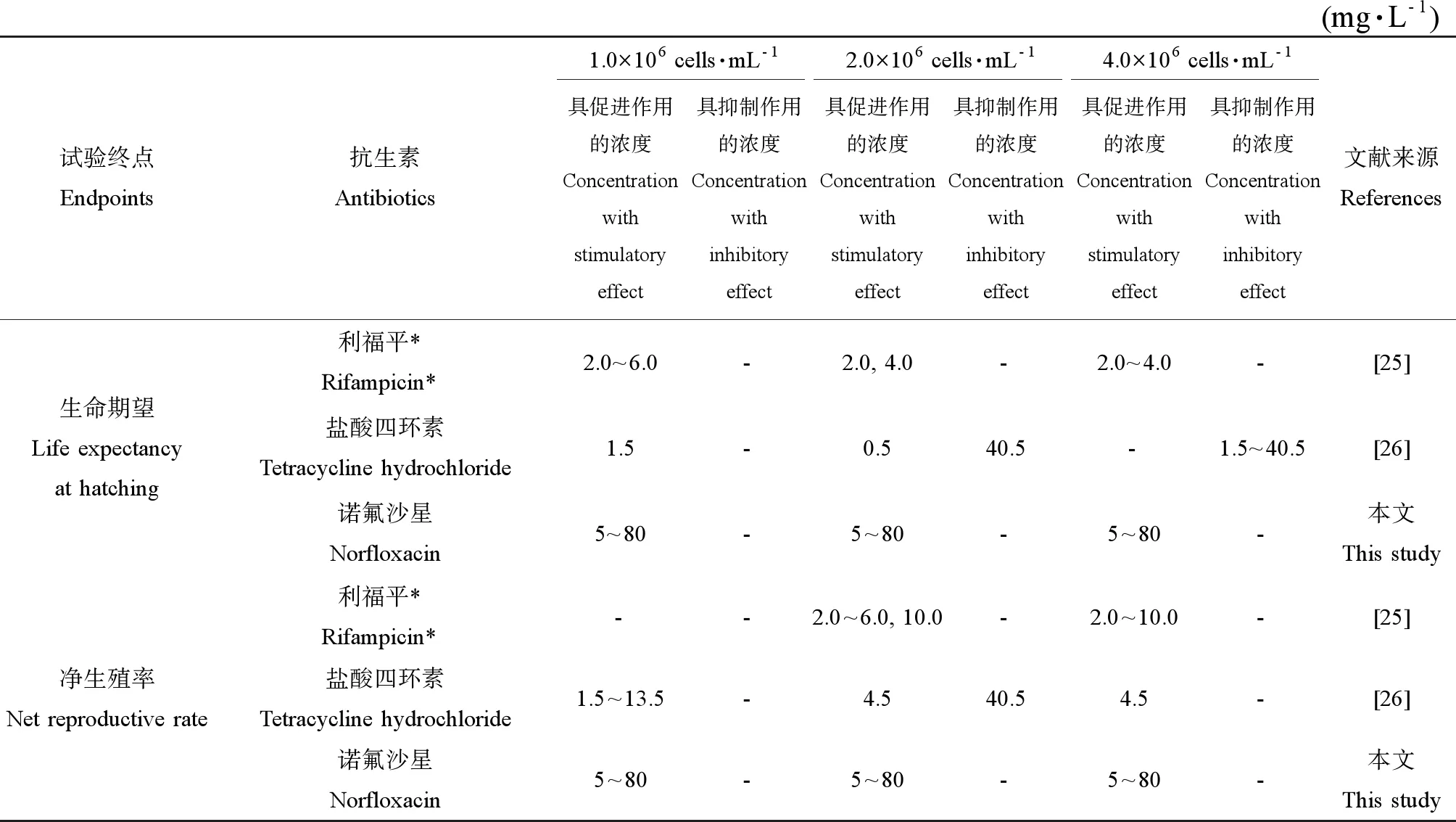
表4 3个斜生栅藻密度下对萼花臂尾轮虫存活和生殖具有促进或抑制作用的抗生素浓度Table 4 The concentrations at which antibiotics had stimulatory or inhibitory effects on the survival and reproduction of B. calyciflorus cultured at three densities of S. obliquus
注: 利福平、盐酸四环素和诺氟沙星的测试浓度范围为2.0~10.0 mg·L-1、0.5~40.5 mg·L-1和5~80 mg·L-1。
Note: * The range of tested concentrations of rifampicin, tetracycline hydrochloride and norfloxacin are 2.0-10.0 mg·L-1, 0.5-40.5 mg·L-1and 5-80 mg·L-1.
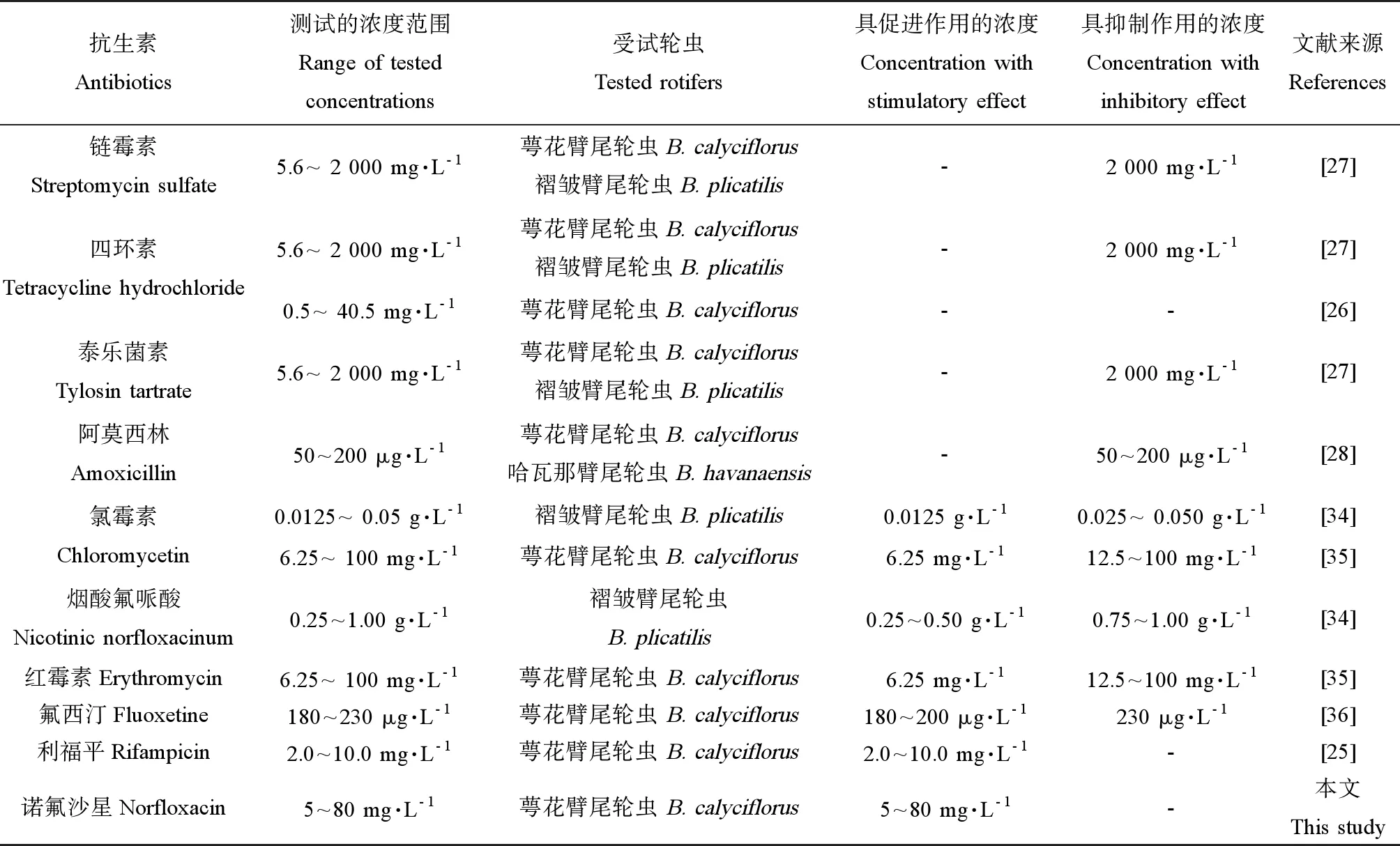
表5 抗生素对轮虫种群增长的影响Table 5 Effects of antibiotics on population growth of rotifers
3.4 抗生素浓度和藻密度对轮虫种群增长的影响
抗生素对轮虫种群增长的影响取决于抗生素种类和测试的浓度范围以及受试生物的种类。总体上,较低浓度的抗生素促进轮虫的种群增长,而较高浓度的抗生素则抑制轮虫的种群增长(表5)。本研究中,3个斜生栅藻密度下,5~ 80 mg·L-1的诺氟沙星均显著提高了萼花臂尾轮虫的种群内禀增长率。更大浓度范围的诺氟沙星对轮虫种群增长的影响仍有待于进一步的研究。
Pourriot[37]认为,在食物充足的情况下,浮游性轮虫在孤雌生殖阶段的增殖速率更多地取决于其胚胎发育速率和生命的早期阶段历时(世代时间),而不是净生殖率。本研究中,当斜生栅藻密度由1.0×106升高至2.0×106cells·mL-1时,6个处理组轮虫净生殖率平均提高幅度减小(19.9%),这可能是轮虫种群内禀增长率平均提高幅度减小(9%)的主要原因,因为世代时间的平均延长幅度仅减小了1.7%。当斜生栅藻密度由2.0×106升高至4.0×106cells·mL-1时,6个处理组轮虫净生殖率平均提高幅度的增大(115.8%)和世代时间平均延长幅度的减小(18.5%)导致了轮虫种群内禀增长率平均提高幅度的增大。
翟盼等[25]认为,一定浓度的利福平能够有效杀灭轮虫及喂食的藻类中可能存在的各种有害细菌,最终对轮虫的存活、孤雌生殖和种群增长产生了积极的影响。项贤领等[26]认为,低浓度的盐酸四环素促进轮虫的存活、孤雌生殖和种群增长可能归因于其“毒物兴奋效应(hormesis)”。作为抗菌药物的代表,适宜浓度的诺氟沙星能够有效杀死革兰氏阴性和阳性菌[38],但其对萼花臂尾轮虫存活、孤雌生殖和种群增长产生积极作用的原因究竟是其杀菌作用,或毒性兴奋效应,或两者兼而有之,仍有待进一步研究。
3.5 轮虫生命表统计学参数对抗生素污染的敏感性
众多的研究表明,轮虫各生命表统计学参数对污染物的敏感性常因污染物的种类等不同而有差异。就抗生素而言,已有的研究发现,萼花臂尾轮虫和褶皱臂尾轮虫的生殖(以净生殖率为指标)比存活对链霉素、四环素和泰乐菌素的敏感性强[27],萼花臂尾轮虫的有性生殖(以后代混交率为指标)比无性生殖(以净生殖率和种群内禀增长率为指标)对利福平的敏感性更强[25]。本研究结果表明,除1.0×106cells·mL-1藻密度下轮虫的后代混交率外,3个藻密度下萼花臂尾轮虫的各生命表统计学参数均对诺氟沙星具有相同的敏感性。从剂量-效应关系角度来看,生命期望、世代时间、净生殖率和内禀增长率适合于评价诺氟沙星对萼花臂尾轮虫的慢性毒性。
总之,亚致死浓度的诺氟沙星促进轮虫的存活、发育、孤雌生殖和种群增长,促进作用的幅度受藻密度的影响显著。亚致死浓度的诺氟沙星对轮虫后代混交率具有抑制作用,抑制作用的显著性与否与诺氟沙星浓度和藻密度有关。
猜你喜欢
口腔护理用品工业(2021年4期)2021-11-02
昆明医科大学学报(2021年8期)2021-08-13
中国生殖健康(2020年8期)2021-01-18
中国生殖健康(2020年2期)2021-01-18
科学养鱼(2020年10期)2020-11-23
课外语文·中(2020年2期)2020-03-24
课外语文(2020年5期)2020-03-14
知识窗(2019年4期)2019-04-26
保健与生活(2019年1期)2019-01-13
中国生殖健康(2018年1期)2018-11-06
