
鼎湖山南亚热带常绿阔叶林荷木种群动态变化
2019-09-10 17:29:14陈青李萌姣李瑶
广西植物 2019年3期
陈青 李萌姣 李瑶
摘要:荷木(Schimasuperba)是鼎湖山南亚热带常绿阔叶林群落中的优势种,为了解南亚热带常绿阔叶林的群落构建机制,该研究分析了鼎湖山南亚热带常绿阔叶林20hm2样地中荷木种群在2005—2017年间的动态变化,并从径级和生境两个方面分析了其死亡率的分布特征。结果表明:(1)荷木种群个体数量从2005—2017年呈现出减少的态势。(2)在径级方面,荷木的死亡率和相对生长率的变化在不同径级区间呈现出不同的变化规律。径级在1~10cm范围的荷木相对生长率大并迅速下降,死亡率上升且维持在较高水平;径级在10~50cm范围的荷木随着径级增大,相对生长率减缓,然后下降,而死亡率不断下降;在50cm以上的径级范围内,荷木相对生长率小、死亡率大。(3)不同生境中,2010年各个生境死亡率相差不大,2015年高海拔生境(高坡、山脊、高谷)死亡率大于低海拔生境(低坡、低谷),2017年则为低海拔生境大于高海拔生境。(4)荷木的死亡率与土壤中酸碱度呈正相关关系,而与土壤有机质、总氮、总磷、有效钾、有效氮含量呈负相关关系。
关键词:径级,死亡率,相对生长率,种群生态,生境
中图分类号:Q948.15
文献标识码:A
文章编号:1000-3142(2019)03-0403-08
种群动态变化是指种群的大小在一定的时间和空间上的分布格局和变化规律,它一直是种群生态学研究非常重要的一个部分(Andrzejczyk&Brzeziecki,1995)。其研究主要通过编制种群生命表(李清河等,2009)、存活曲线、谱分析法(伍业钢和韩进轩,1988)、动态模型法(Tilmanetal.,1994;Rhodesetal.,1996)等方法。生命表可以表示死亡率、存活率等重要信息,存活曲线直观且常见。谱分析法一直作为昆虫种群动态变化的研究方法。伍业钢和韩进轩(1988)首次将其用于森林种群动态变化研究中。针对种群的空间分布格局,Tilmanetal.(1997)在Levins模型(Levins,1969)的基础上,提出了Tilman多物种集合模型。Rhodesetal.(1996)将其分为空间隐含模型和空间显含模型。
树木的死亡是森林群落中的一种普遍现象(Riceetal.,2004)。树木的死亡会使群落物种组成和群落格局等发生变化,进而影响演替进程(Mencuccinietal.,2005)。树木死亡也会造成环境因子的变化:如林窗增加、土壤成分改变、森林区域气候的变化等(Dasetal.,2007)。死亡树木的凋落物会参与到生态系统的循环中,是森林元素循环的重要环节,也是调节森林生态系统结构和动态的重要机制(McCoy&Gillooly,2008;Lewisetal.,2004)。树木的死亡一般是由多因素相互作用和相互制约而形成的(Marbaetal.,2007),长期因素如个体竞争、病虫害、气候变化、土壤水分条件变化、突发干扰等,突发因素如突发性的火灾、虫灾、天气干扰等(Brandt,2013)。
南亚热带常绿阔叶林是一种典型的热带向亚热带过渡的植被类型(彭少麟,1998)。鼎湖山南亚热带常绿阔叶林20hm2样地(简称“鼎湖山大样地”)是中国森林生物多样性监测网络的核心样地之一(叶万辉等,2008)。因为当地人文环境的影响,鼎湖山很好地保存了400多年南亚热带常绿阔叶林地带性森林群落的结构特征。针对鼎湖山大样地群落构建机制,已从群落(王志高等,2008;林国俊等,2010;叶万辉等,2008;Lietal.,2009;Wangetal.,2009)、種群遗传(Wangetal.,2012)、进化(Peietal.,2011)等方面进行了研究。魏识广等(2008)对鼎湖山大样地的格木种群动态进行了分析,结果显示该群落中的格木正处于加速增长状态。
荷木(Schimasuperba)隶属于山茶科(Theaceae)木荷属,大乔木,叶革质,椭圆形,在两面明显,边缘有钝齿;叶柄长1~2cm。花白色,生于枝顶叶腋,常多朵排成总状花序,花柄长,苞片早落;萼片半圆形,长2~3mm;花瓣长1~1.5cm。蒴果直径1.5~2cm。花期6—8月(张宏达和任善湘,1998)。荷木是鼎湖山大样地中极为重要的建群种和优势种(叶万辉等,2008),对鼎湖山森林群落的构建和其它物种的空间分布都有重要作用,故而其在鼎湖山大样地的分布和动态变化在南亚热带的森林群落构建中十分具有代表性。因此,对鼎湖山南亚热带常绿阔叶林荷木种群的死亡、分布、数量变化的研究,对于探究该群落的构建机制有十分重要的意义。本研究以鼎湖山大样地的荷木种群为对象,分析其从2005—2017年期间的种群动态变化并探究可能的原因,研究结果不仅有利于探讨鼎湖山南亚热带常绿阔叶林的群落构建机制,而且对于该地带的森林生态系统保护和恢复有重要意义。
1材料与方法
1.1研究区自然概况
鼎湖山国家级自然保护区(112°30′39″—112°33′41″E,23°09′21″—23°11′30″N)地处广东省肇庆市东北部。该地区位于亚热带季风气候区南端,接近热带(吴厚水,1982)。其年平均气温为20.9℃,雨季月降水量均超过200mm,旱季月降水量不到100mm,土壤类型为赤红壤、黄壤和山地灌丛草甸土(叶万辉等,2008)。彭少麟等(1998)将鼎湖山森林群落的演替划分为六个阶段:(1)针叶林;(2)以针叶树种为主的针阔混交林;(3)以阳生阔叶树种为主的针阔混交林;(4)以阳生植物为主的常绿阔叶林;(5)以中生植物为主的常绿阔叶林;(6)中生群落。本研究中的南亚热带常绿阔叶林群落处于鼎湖山森林群落演替的第五阶段。
1.2研究样地概况
1.2.1大样地的设置鼎湖山南亚热带常绿阔叶林20hm2监测样地于2005年根据CTFS(CentreforTropicalScience)样地建设标准(Condit,1998)在鼎湖山国家级自然保护区的核心地区建立。鼎湖山大样地是我国第一批森林生物多样性固定监测样地之一(叶万辉等,2008)。
1.2.2大样地五种生境的划分将鼎湖山大样地划分为五种生境类型,参考因素有海拔(H)、坡度(S)的中值、和凹凸度(C)(练琚愉等,2015)。具体如下:高坡(higherslope,HS),S>median(S),≥median(H),C>0;高谷(highergully,HG),S>median(S),H≥median(H),C<0;
山脊(mountainridge,MR),S≤median(S),H≥median(H);低坡(lowerslope,LS),H0;低谷(lowergully,LG),H
1.3野外调查
2005年鼎湖山大样地建立之后完成了首次调查,随后分别于2010年、2015年进行了两次复查。野外调查中,标记样地内胸径(DBH)≥1cm的全部植物个体,记录其编号、胸径、坐标、生长情况等信息。本研究中荷木的全部个体信息均提取自这三次调查的数据库。另外,为了记录荷木种群的现况,2017年对鼎湖山大样地中所有胸径(DBH)≥1cm的荷木个体进行了补充调查,同样记录上述信息。
1.4数据统计分析
所有数据用Excel13.0和SPSS21.0进行统计与分析,其中计算的指标包括:死亡率(mortalityrate)=(本次存活个体数-上次存活个体数)/(上次存活个体数×年数);相对生长率(relativegrowthrate,RGR)=[lg(本次胸径)-lg(上次胸径)]/[lg(上次胸径)×年数];死亡率的变异系数(coefficientofvariation,CV)=(标准差/平均值)×100%。
使用Pearson相关分析法检验荷木死亡率与土壤因子(酸碱度、有机质、有效钾、总磷、总氮和有效氮)的相关性,P<0.05表示有显著相关性,P<0.01表示有极显著相关。
2结果与分析
2.1鼎湖山大样地荷木种群的整体动态
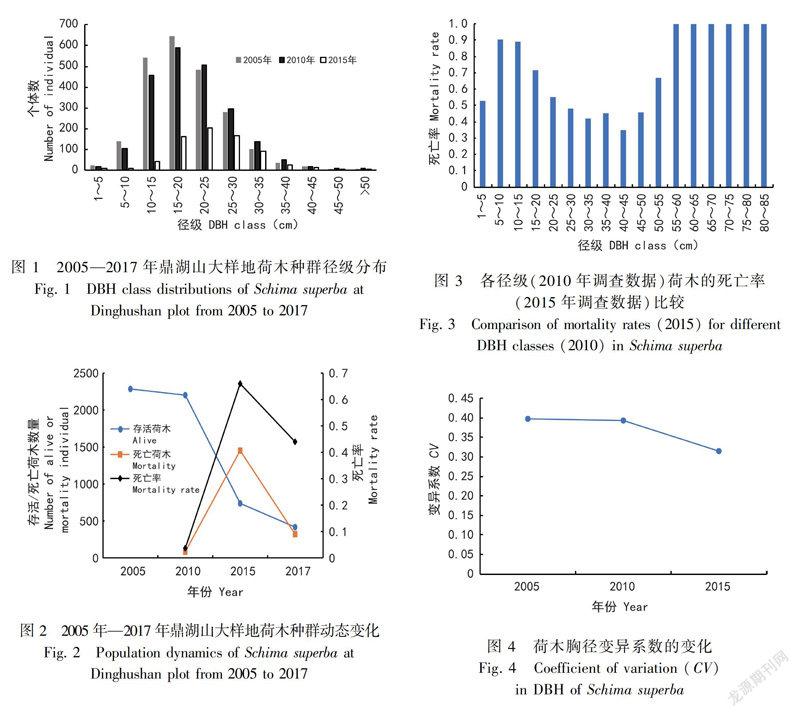
在2005年、2010年和2015年三次调查中,鼎湖山大样地的荷木种群个体径级结构(图1)均为正态分布,表明荷木的个体大部分位于中径级阶段,而小径级与大径级的个体数则相对较少,即中径级个体储备型(叶万辉等,2008)。
2005年鼎湖山大样地的荷木有2287株个体,随后于2010年略微减少到2199株,但2015年迅速下降到745株,死亡率显著增加。2015—2017年期间荷木死亡率有所下降,但其个体数量依然在減少。
2.2不同径级的荷木个体数量的动态变化
按照径级分段并计算死亡率,可以看出鼎湖山大样地荷木的各径级的个体死亡率总体呈U型(图3),即小径级(0~25cm)和大径级(50~85cm)的死亡率较大,均超过0.5,而中等径级(25~50cm)个体的死亡率较小,均小于0.5。其中,55~85cm径级中的大径级个体在2010年存活数量较少,在2015年全部死亡,故死亡率达到1。
对鼎湖山大样地历年荷木个体胸径计算变异系数CV=SD/mean(图4),发现呈逐渐下降趋势,特别是在2010—2015年迅速下降。
2.3不同生境中荷木个体数量的动态变化
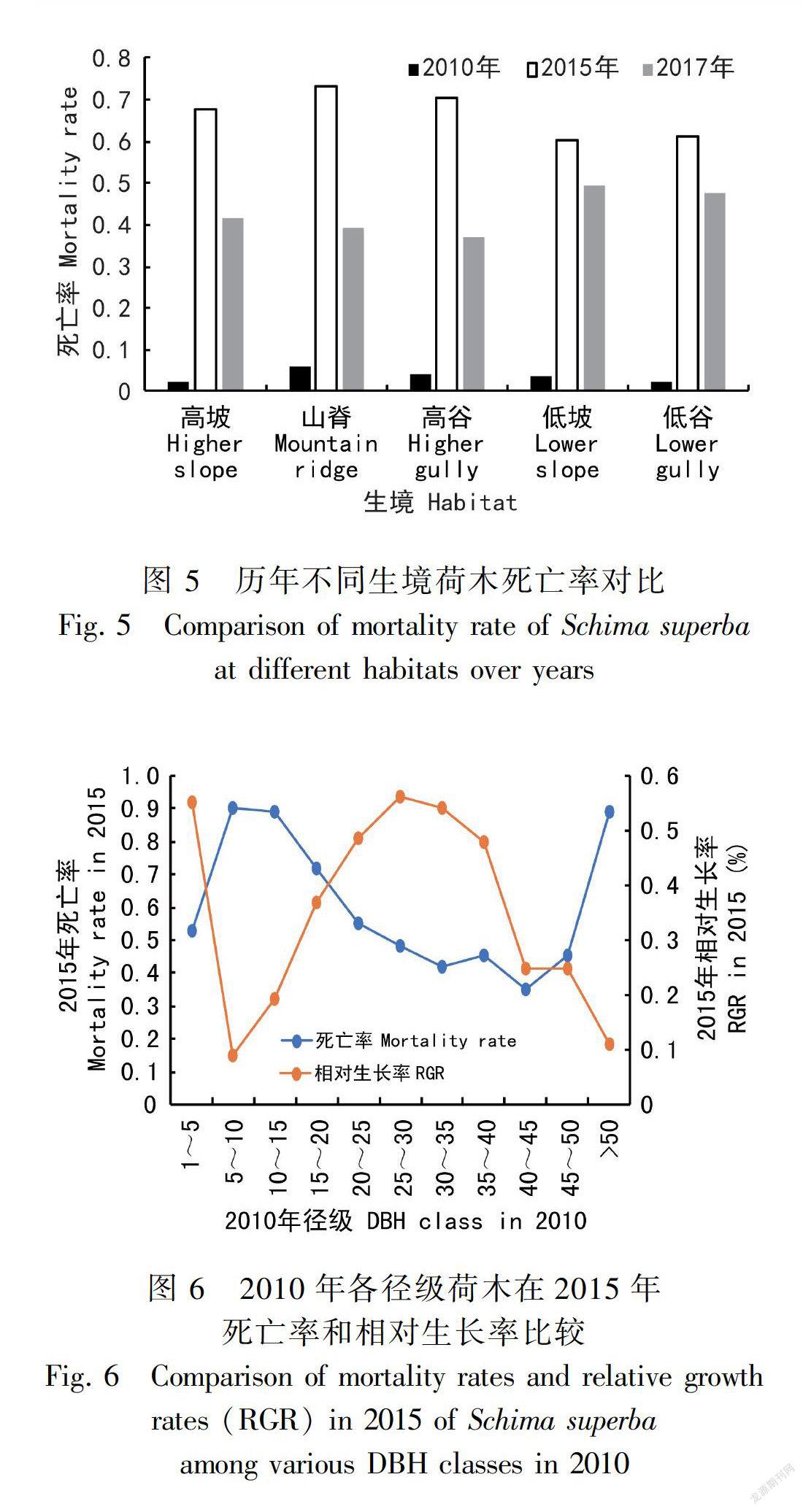
从图5可以看出,2010年各个生境死亡率相差不大,2015年开始出现差异,高海拔生境(高坡、山脊、高谷)死亡率大于低海拔(低坡、低谷),2017年低海拔生境大于高海拔。这是因为高海拔的荷木先受到虫害的影响,竞争力较弱的树已经死亡,环境筛选中剩下的荷木存活能力强,由此造成2017年统计的时候,其死亡率较低海拔的小。
2.4荷木种群动态变化原因的初步探究
2.4.1荷木个体死亡率和相对生长率的关系对荷木死亡率和相对生长率按照径级分组并作折线图(图6)。从图6可以看出,(1)径级在1~10cm范围的荷木个体相对生长率逐渐下降至最低点,死亡率上升达到第一个峰值;(2)径级在10~50cm范围的荷木相对生长率减速上升至峰值后开始下降,而死亡率不断降低;(3)在50cm以上的径级范围内,荷木个体的相对生长率小,而死亡率大。
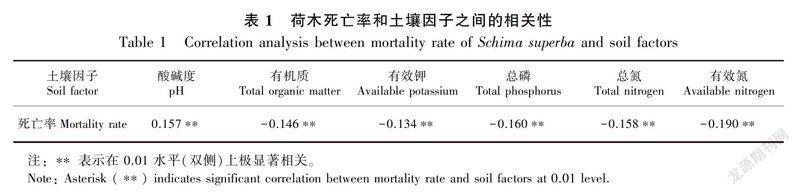
2.4.2荷木个体死亡率和土壤因子的关系对荷木死亡率和环境因子做Pearson相关分析,结果见表1。由表1可知,荷木个体的死亡率和土壤中酸碱度为正相关关系,而和土壤有机质、总氮、总磷、有效钾、有效氮含量呈负相关。说明荷木适宜在酸性土壤中生存,同时土壤中的有机质、有效钾、总磷、总氮、有效氮含量越高,越有利于荷木的存活。
3讨论与结论
3.1鼎湖山大样地荷木种群的整体动态变化
在森林群落中,乔木优势种和建群种能够很好地反映出其演替状况,这是因为优势种和建群种的动态变化能够决定其群落的构建和特征(Ni,2001)。张宏达等(1955)指出,由于荷木是阳性树种,不耐阴,只能作为先锋树种向林缘发展,而不能在鼎湖山森林群落中发展下去,说明该群落发展处于不平衡状态,即荷木已达到成熟的阶段,并开始走下坡路。这与其幼苗的更新能力较差有关。彭少麟和方炜(1995)提出了鼎湖山常绿阔叶林的演替趋势为黄果厚壳桂-锥栗-厚壳桂-荷木群落向黄果厚壳桂-厚壳桂群落方向进行。本研究进一步发现,在2005—2017年之间,鼎湖山大样地荷木的个体数量持续降低,特别是2015年统计的下降速度最快,死亡率最高。其重要值由2005年的第2位(叶万辉等,2008)降为2017年的第8位,这些均证实了张宏达等(1955)及彭少麟和方炜(1995)的推论。
3.2不同径级的荷木个体数量动态变化
树木的径级和年龄呈正相关关系,因此在野外调查和研究中,出于条件的限制和成本的考虑,常常会使用径级代表年龄森林群落的树木个体年龄。而不同径级的树木死亡率往往并不均衡。刘万德等(2010)对海南霸王岭林区的热带雨林的研究表明,树木死亡主要集中于胸径小于30cm的树木。梁军等(2011)对昆嵛山天然林13a演替动态的研究表明,死亡树木中,胸径小于10cm的幼树所占比例较高。
本研究结果表明鼎湖山大样地荷木中径级小于25cm的幼树和大于50cm的老树死亡率较高。径级大于55cm时,死亡率为100%。这符合Nepstadetal.(2007)在亚马逊森林的研究结果:大树的死亡率高于小树的死亡率。大树因为繁殖消耗和自然衰老使得大树更易受到自然灾害的影响(Muelleretal.,2005)。但根据梁军(2011)的研究,在虫灾的干扰下,幼树的死亡情况会更严重。在2010—2015年期间,鼎湖山大样地森林受到虫灾干扰。因此在本文的研究结果中,幼树和老树死亡率高,而青壮年树木得以在干扰中存活,使得荷木种群的变异系数逐年变小,胸径离散度缩小。
3.3不同生境中荷木个体数量的动态变化
在生境异质性强的森林群落中,种群的分布格局和生存状态往往与其所处的生境密切相关。不同的生境下植物群落会呈现不同的群落特征(王立龙等,2015)。生境因子会导致群落树木死亡率的变化和功能群结构的变化从而改变群落结构和物种生存状态(刘万德等,2010;周光霞等,2016)。本研究结果表明,尽管2010年各个生境中荷木的个体死亡率相差不大,但是到2010年荷木死亡率急剧增高,而且高海拔生境(山脊、高坡、高谷)的死亡率明显大于低海拔生境(低坡、低谷),这可能与2005—2015年期间鼎湖山地区爆发的虫灾有关。鼎湖山大样地高海拔生境的林窗平均面积和林窗空隙率均大于低海拔生境(隋丹丹等,2017),而已有研究表明林窗生境的荷木遭受虫食损失率更大(王宏伟和蔡永立,2009),这可能是导致本研究中高海拔生境荷木在遭受虫灾后死亡率更大的原因。而在2017年,低海拔生境的荷木个体死亡率大于高海拔生境,这一方面与计算死亡率的基数有关,因为2010年高海拔生境幸存下来的荷木个体显著减少;另一方面低海拔生境的虫灾影响相比高海拔生境相对滞后于低海拔生境;最后还可能与荷木的阳性特征(不耐阴)密切相关(张宏达等,1955)。
3.4荷木种群数量变化原因的初步探究
如前文所示,鼎湖山大样地荷木种群数量在2005—2017年之间发生了巨大变化。树木的死亡一般是一个由量变到质变的过程(Franklinetal.,1987)。导致荷木死亡的原因有生物因子和非生物因子。在鼎湖山大样地的荷木种群中,主要生物影响因素有植食者、种间和种内竞争等,以及非生物因素包括地形和土壤因子。
在研究中,径级在1~10cm范围的荷木相对生长率大,死亡率大。这是因为幼年荷木細胞活性强,相对生长率大,而不如成年树木稳定(吴可等,2010)。径级在5~10cm范围的荷木相对生长率不断下降,而死亡率不断上升,可能是由于这个年龄的荷木对虫灾等环境干扰比较敏感。径级10~50cm的荷木相对生长率减速上升至峰值后开始下降,而死亡率不断降低。这是因为青壮年荷木对干扰的抵抗能力最强,幸存荷木个体的生长性能得到恢复。而老年荷木,即50cm以上的径级范围的荷木数量少、死亡率大,相对生长率小。随着年龄增加,树木的功能器官逐渐衰老,相对生长率变小,不仅对外界干扰的抵抗力差,而且也会随着自然衰老而死亡。
鼎湖山大样地的荷木受到环境中土壤因子的影响很大。荷木的死亡率与土壤中酸碱度呈正相关,和土壤有机质、总氮、总磷、有效钾、有效氮含量呈负相关。这与安树青等(1997)对南京紫金山森林群落植物的研究结果类似。安树青等(1997)研究发现土壤酸碱度、厚度、含水量、有效氮和速效磷对北亚热带森林群落有着重要影响。而地处珠江三角洲边缘的鼎湖山自然保护区降水中氮沉降量也由20世纪80年代的36kg·hm-2·a-1(黄忠良等,1998)上升到了90年代的38kg·hm-2·a-1(周国逸和闫俊华,2001),这一沉降量与欧洲和北美的一些高氮沉降区的氮沉降量相当。而氮沉降可能会加剧亚热带森林土壤酸化(Luetal.,2014)。因此荷木死亡的可能原因是受到鼎湖山土壤环境的干扰,这些土壤因子会影响土壤中微生物的群落结构以及荷木根部的根际环境(刘菊秀等,2003),从而间接影响荷木对水分和养分的吸收,影响荷木的生长能力和防御能力。
在当前气候环境下,对于鼎湖山大样地荷木种群13a来的动态变化和影响因子的研究可以让我们能够更好地理解南亚热带森林的群落构建和演替过程,对于预测种群发展,提高森林对外界干扰的抵抗能力有十分重要的价值。
参考文献:
ANSQ,WANGZF,ZHUXL,etal.,1997.Effectsofsoilfactorsonspeciesdiversityinsecondaryforestcommunities[J].JWuhanBotRes15(2):143-150.[安树青,王峥峰,朱学雷,等,1997.土壤因子对次生森林群落物种多样性的影响[J].武汉植物学研究,15(2):143-150.]
ANDRZEJCZYKT,BRZEZIECKIB,1995.Thestructureanddynamicsofold-growthPinussylvestris(L.)standsintheWigryNationalPark,north-easternPoland[J].Vegetatio,117(1):81-94.
BRANDTAJ,KROONH,REYNOLDSHL,etal.,2013.Soilheterogeneitygeneratedbyplant-soilfeedbackshasimplicationsforspeciesrecruitmentandcoexistence[J].JEcol,101(2):277-286.
CHANGHT,RENSX,1998.FloraReipublicaePopularisSinicae[M].Beijing:SciencePress,49(3):1-251.[张宏达,任善湘,1998.中国植物志[M].北京:科学出版社,49(3):1-251.]
CHANGHT,WANGBS,ZHANGCC,etal.,1955.ThestudyontheplantcommunityofDinghuMountaininGuangdongProvince[J].ActaSciNatUnivSunyatseni,(3):159-227.[张宏达,王伯荪,张超常,等,1955.广东高要鼎湖山植物群落之研究[J].中山大学学报(自然科学版),(3):159-227.]
CONDITR,1998.Tropicalforestcensusplots:MethodsandresultsfromBarroColoradoIsland,Panamaandacomparisonwithotherplots[M].Berlin:Springer:23-55.
DASAJ,BATTLESJJ,STEPHENSONNL,etal.,2007.Therelationshipbetweentreegrowthpatternsandlikelihoodofmortality:AstudyoftwotreespeciesintheSierraNevada[J].CanJForRes,37(3):580-597.
FRANKLINJF,SHUGARTHH,HARMONME,1987.Treedeathasanecologicalprocess[J].Bioscience,37(8):550-556.
GUYY,ZHANGSQ,LIXY,etal.,2013.RelationshipbetweendiameteratbreastheightandageofendangeredspeciesPopuluseuphraticaOliv[J].JTarimUniv,25(2):66-69.[顾亚亚,张世卿,李先勇,等,2013.濒危物种胡杨胸径与树龄关系研究[J].塔里木大学学报,25(2):66-69.]
HUANGZL,KONGGH,WEIP,1998.PlantspeciesdiversitydynamicsinDinghuMountainforests[J].ChinBiodivers,6(2):116-121.[黄忠良,孔国辉,魏平,1998.鼎湖山植物物种多样性动态[J].生物多样性,6(2):116-121.]
LEVINSR,1969.Somedemographicandgeneticconsequencesofenvironmentalheterogeneityforbiologicalcontrol[J].EntomolSocAmBull,15(3):237-240.
LEWISSL,PHILLIPSOL,SHEILD,etal.,2004.Tropicalforesttreemortality,recruitmentandturnoverrates:Calculation,interpretationandcomparisonwhencensusintervalsvary[J].JEcol,92(6):929-944.
LIL,HUANGZL,YEWH,etal.,2009.SpatialdistributionsoftreespeciesinasubtropicalforestofChina[J].Oikos,118(4):495-502.
LIQH,GAOTT,LIUJF,etal.,2009.TheagestructureandlifetableofrareeremophyteHelianthemumordosicumpopulation[J].BullBotRes,29(2):176-181.[李清河,高婷婷,刘建锋,等,2009.荒漠珍稀灌木半日花种群的年龄结构与生命表分析[J].植物研究,29(2):176-181.]
LIANJY,CHENC,HUANGZL,etal.,2015.Communitycompositionandstandageinasubtropicalforest,southernChina[J].BiodiversSci,23(2):174-182.[练琚愉,陈灿,黄忠良,2015.鼎湖山南亚热带常绿阔叶林不同成熟度群落特征的比较[J].生物多样性,23(2):174-182.]
LIANGJ,SUNZQ,ZHUYP,etal.,2011.13-yearssuccessiondynamicofKunyushannaturalforest[J].JCentSUnivForTechnol,31(1):9-17.[梁军,孙志强,朱彦鹏,等,2011.昆嵛山天然林13年演替动态[J].中南林业科技大学学报,31(1):9-17.]
LINGJ,HUANGZL,ZHUL,etal.,2010.BetadiversityofforestcommunityonDinghushan[J].ActaEcolSin,30(18):4875-4880.[林國俊,黄忠良,竺琳,等,2010.鼎湖山森林群落β多样性[J].生态学报,30(18):4875-4880.]
LIUJX,ZHOUGY,ZHANGDQ,2003.ThecumulativeeffectsofacidrainonthesoilandresponsesofSchimasuperbaatDinghushan[J].ChinEnvironSci,23(1):90-94.[刘菊秀,周国逸,张德强,2003.酸雨对鼎湖山土壤的累积效应及荷木的反应[J].中国环境科学,23(1):90-94.]
LIUWD,ZANGRG,DINGY,etal.,2010.MortalityofwoodyplantsintropicalmonsoonrainforestsofBawanglingNationalNatureReserveonHainanIsland,SouthChina[J].ChinJPlantEcol,34(8):62-72.[刘万德,臧润国,丁易,等,2010.海南岛霸王岭热带季雨林树木的死亡率[J].植物生态学报,34(8):62-72.]
LUXK,MAOQG,GILLIAMFS,etal.,2014.Nitrogendepositioncontributestosoilacidificationintropicalecosystems[J].GlobChangeBiol,20(12):3790-3801.
MARBAN,DUARTECM,AGUSTIS,2007.Allometricscalingofplantlifehistory[J].ProcNatlAcadUSA,104(40):15777-15780.
MCCOYMW,GILLOOLYJF,2008.Predictingnaturalmortalityratesofplantsandanimals[J].EcolLett,11(7):710-716.
MENCUCCINIM,MARTINEZ-VILALTAJ,VANDERKLEIND,etal.,2005.Size-mediatedageingreducesvigourintrees[J].EcolLett,8(11):1183-1190.
MUELLERRC,SCUDDERCM,PORTERME,etal.,2005.Differentialtreemortalityinresponsetoseveredrought:Evidenceforlong-termvegetationshifts[J].JEcol,93(6):1085-1093.
NEPSTADDC,TOHVERIM,RAYD,etal.,2007.MortalityoflargetreesandlianasfollowingexperimentaldroughtinanAmazonforest[J].Ecology,88(9):2259-2269.
NIJ,2001.PlantfunctionaltypesandbiomesofChinaataregionalscale[J].ActaBotSin,43(4):419-425.[倪健,2001.区域尺度的中国植物功能型与生物群区[J].植物学报,43(4):419-425.]
PEINC,LIANJY,ERICKSONDL,etal.,2011.Exploringtree-habitatassociationsinaChinesesubtropicalforestplotusingamolecularphylogenygeneratedfromDNAbarcodeloci[J].PLoSONE,6(6):e21273.
PENGSL,FANGW,1995.StudiesondynamicsofCastanopsischinensisandSchimasuperbapopulationinforestsuccessionofDinghushanmountain[J].ActaPhytoecolSin,19(4):311-318.[彭少麟,方炜,1995.鼎湖山植被演替过程中椎栗和荷木种群的动态[J].植物生态学报,19(4):311-318.]
PENGSL,FANGW,RENH,etal.,1998.ThedynamicsonorganizationinthesuccessionalprocessofDinghushanCryptocaryacommunity[J].ActaPhytoecolSin,22(3):245-249.[彭少麟,方炜,任海,等,1998.鼎湖山厚壳桂群落演替过程的组成和结构动态[J].植物生态学报,22(3):245-249.]
RHODESOE,CHESSERRK,SMITHMH,1996.Populationdynamicsinecologicalspaceandtime[M].Chicago:UniversityofChicagoPress:122-123.
RICEKJ,MATZNERSL,BYERW,etal.,2004.PatternsoftreediebackinQueensland,Australia:theimportanceofdroughtstressandtheroleofresistancetocavitation[J].Oecologia,139(2):190-198.
SUIDD,WANGY,LIANJY,etal.,2017.Gapdistributionpatternsinthesouthsubtropicalevergreenbroad-leavedforestofDinghushan[J].BiodiversSci,25(4):382-392.[隋丹丹,王悅,练琚愉,等,2017.鼎湖山南亚热带常绿阔叶林林窗分布格局及其成因[J].生物多样性,25(4):382-392.]
TILMAND,LEHMANCL,YINC,1997.Habitatdestruction,dispersal,anddeterministicextinctionincompetitivecommunities[J].AmNat,149(3):407-435.
TILMAND,MAYRM,LEHMANCL,1994.Habitatdestructionandtheextinctiondebt[J].Nature,371:65-66.
WANGHW,CAIYL,2009.Spatialvariationsininsectherbivoryleavesinsubtropicalevergreenbroadleavedforest[J].JShanghaiInstTechnol(NatSciEd),9(2):110-114.[王宏伟,蔡永立,2009.昆虫取食常绿阔叶林木荷叶片的空间变化[J].上海应用技术学院学报(自然科学版),9(2):110-114.]
WANGLL,WANGL,ZHANGLF,etal.,2015.StructureanddynamiccharacteristicsofGymnocarposprzewalskiiindifferenthabitats[J].ChinJPlantEcol,39(10):980-989.[王立龙,王亮,张丽芳,等,2015.不同生境下濒危植物裸果木种群结构及动态特征[J].植物生态学报,39(10):980-989.]
WANGZF,LIANJY,HUANGGM,etal.,2012.GeneticgroupsinthecommonplantspeciesCastanopsischinensisandtheirassociationswithtopographichabitats[J].Oikos,121(12):2044-2051.
WANGZG,YEWH,CAOHL,etal.,2008.SpatialdistributionofspeciesdiversityindicesinamonsoonevergreenbroadleavedforestatDinghushanMountain[J].BiodiversSci,16(5):454-461.[王志高,葉万辉,曹洪麟,等,2008.鼎湖山季风常绿阔叶林物种多样性指数空间分布特征[J].生物多样性,16(5):454-461.]
WANGZG,YEWH,CAOHL,etal.,2009.Species-topographyassociationinaspecies-richsubtropicalforestofChina[J].BasicApplEcol,10(7):648-655.
WEISG,LIL,LIUHG,etal.,2008.AnalysesofthedynamicstateofErythrophleumfordiipopulation[J].EcolEnviron,17(1):285-289.[魏识广,李林,刘海岗,等,2008.鼎湖山格木种群动态分析[J].生态环境学报,17(1):285-289.]
WUHS,1982.DinghuMountainNatureReserveinhydrothermalconditionanditsrelationshipwithecologicalenvironment[J].TropGeogr,2(4):14-20.[吴厚水,1982.鼎湖山自然保护区水热状况及其与生态环境的关系[J].热带地理,2(4):14-20.]
WUK,YINMF,ZHOUYB,etal.,2010.CurrentannualgrowthrateofforestvolumeinBaishilaziNationalNatureReserve[J].JNWForUniv,25(6):203-206.[吴可,殷鸣放,周永斌,等,2010.白石砬子国家级自然保护区森林蓄积连年生长率的研究[J].西北林学院学报,25(6):203-206.]
WUYG,HANJX,1988.Aspectralanalysisofthepopulationdynamicsofkoreanpineinthemixedbroad-leavedPinuskoraiensisforest[J].ChinJEcol,7(1):19-23.[伍业钢,韩进轩,1988.阔叶红松林红松种群动态的谱分析[J].生态学杂志,7(1):19-23.]
YEWH,CAOHL,HUANGZL,etal.,2008.Communitystructureofa20hm2lowersubtropicalevergreenbroadleavedforestplotinDinghushan,China[J].JPlantEcol,32(2):274-286.[叶万辉,曹洪麟,黄忠良,等,2008.鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究[J].植物生态学报,32(2):274-286.]
ZHOUGX,HUANGLX,ZANGXW,etal.,2016.EffectsofhabitatheterogeneityoncommunityfunctionaldiversityofDinghuMountainevergreenbroad-leavedforest[J].Guihaia,36(2):127-136.[周光霞,黄立新,臧晓蔚,等,2016.生境异质性对鼎湖山常绿阔叶林群落功能多样性的影响[J].广西植物,36(2):127-136.]
ZHOUGY,YANJH,2001.TheinfluencesofregionalatmosphericprecipitationcharacteristicsanditselementinputsontheexistenceanddevelopmentofDinghushanforestecosystems[J].ActaEcolSin,21(12):2002-2012.[周国逸,闫俊华,2001.鼎湖山区域大气降水特征和物质元素输入对森林生态系统存在和发育的影响[J].生态学报,21(12):2002-2012.]
猜你喜欢
广西林业科学(2022年4期)2022-11-07 01:50:34
林业调查规划(2022年3期)2022-06-13 09:31:04
广西林业科学(2021年6期)2022-01-21 07:39:10
中老年保健(2021年4期)2021-08-22 07:07:02
今日农业(2021年5期)2021-05-22 01:32:38
落叶果树(2021年6期)2021-02-12 01:29:10
科学之谜(2020年6期)2020-08-11 07:37:21
当代水产(2019年8期)2019-10-12 08:57:56
体育科技(2016年2期)2016-02-28 17:06:07
西北植物学报(2015年9期)2015-07-05 07:03:42
