
水分胁迫下竹叶花椒的生理响应
2019-09-10 07:22:44王纪辉侯娜粱美
江苏农业学报 2019年3期
关键词:水分胁迫
王纪辉 侯娜 粱美



摘要:为揭示水分胁迫对竹叶花椒(Zanthoxylum armatum DC)生理指标的影响,以及竹叶花椒耐旱机制,以1年生花椒实生苗为试验材料,采用盆栽试验,对水分胁迫后竹叶花椒幼苗的超氧化物歧化酶活性、过氧化物酶活性、相对含水量、丙二醛含量、叶绿素含量、可溶性蛋白质含量的变化规律进行了研究。结果表明,干旱胁迫下竹叶花椒超氧化物歧化酶、过氧化物酶活性保持良好,丙二醛含量缓慢增加,相对含水量和叶绿素含量逐渐下降,可溶性蛋白质含量呈“V”型變化。相关性分析结果表明,部分指标间表现出显著正(负)相关,部分指标间呈一定的相关性但不显著。主成分分析结果表明,过氧化物酶活性、丙二醛含量、相对含水量、叶绿素含量、可溶性蛋白质含量与竹叶花椒耐旱性关系密切,可作为评价其耐旱性的指标。
关键词:竹叶花椒;水分胁迫;生理响应
中图分类号: Q945.78
文献标识码:A
文章编号: 1000-4440( 2019) 03-0676-06
竹叶花椒(Zanthoxylum armatum DC)是芸香科( Rutaceae)花椒属落叶小乔木,花椒中酰胺类物质和挥发油含量丰富,因而麻香味十分浓郁,是受广大消费者青睐的佐料[1]。花椒因具有温中散寒、除湿、止痛等功能,又被纳入传统中药材的行列,成为药食两用的重要植物[2]。近几年来,花椒产业发展十分迅速,已成为中国西南、西北地区的重要经济树种[3]。花芽分化是花椒产量高低的决定性因素,确保一定的花芽数量,是花椒高产稳产的基础[4]。但不良环境会减弱花椒花芽分化程度,造成花椒大幅度减产[5]。因此,研究水分胁迫对竹叶花椒生理指标的影响,为初步阐明竹叶花椒耐旱机理,解决其目前面临的干旱问题奠定基础。研究发现,干旱胁迫下作物超氧化物歧化酶( SOD)协同过氧化物酶(POD)共同发挥作用,清除自由基[6]。干旱胁迫阻碍叶绿素合成,叶绿素分解速率大于合成速率,叶绿素含量整体上表现为下降趋势;干旱胁迫导致植物细胞膜系统被破坏,膜脂发生过氧化,丙二醛(MDA)含量增加,相对含水量(RWC)逐渐下降,而可溶性蛋白质(SP)合成速率大于分解速率,其含量整体上呈增加趋势[7]。研究结果表明,水分胁迫在一定程度上能提高SOD和POD活性,加速膜脂过氧化,增加MDA含量,降低叶片相对含水量,刺激蛋白酶合成新的可溶性蛋白质来适应环境[8-10]。张寅媛等的研究结果表明,干旱胁迫能迫使景天科植物叶片相对含水量下降,细胞膜系统遭到破坏,MDA含量增加,胁迫初期SOD活性升高、后期下降,可溶性蛋白质含量增加[11]。胡宏远等研究发现,水分胁迫能抑制叶绿素合成,加速其分解,并降低净光合速率[12]。干旱胁迫能阻碍作物光合作用,降低净光合速率[13-14]。目前,有关竹叶花椒对水分胁迫的生理响应方面的研究报道较少。本研究以1年生竹叶花椒幼苗叶片为试验材料,揭示水分胁迫下竹叶花椒的生理响应,旨在为竹叶花椒栽培、品种选择以及耐旱性能等研究提供科学依据。
1 材料与方法
1.1 试验材料
竹叶花椒1年生实生苗取自贵州大学林学院苗圃基地,属于延迟脱水耐旱类型。2016年2月在半拱形覆膜日光温室中育苗。盆栽试验于2017年2月在贵州大学林学院科研基地半拱形覆膜日光温室中进行,取园土、腐殖质、砂子按质量比4:1:1混合,装入高27 cm、直径26 cm的塑料盆中,每盆装土12 kg,土壤田间最大持水量为24.12%。每盆栽植l株,正常浇水,保持幼苗正常生长。移栽后2个月,移到遮雨棚中进行控水处理。设置4个水分梯度处理:对照( CK),土壤含水量为田间最大持水量的70% -80%;轻度胁迫(Tl),土壤含水量为田间最大持水量的50% - 60%;中度胁迫(T2),土壤含水量为田间最大持水量的40% - 50%;重度胁迫(T3),土壤含水量为田间最大持水量的30% - 40%。每个处理10盆,3次重复。采用称量法控水和补水,每天18:00称量,补充当天消耗的水分,使土壤含水量控制在设定范围之内。
1.2 测定指标与方法
达到设定的水分梯度后,维持20 d。采集各处理植株上相同节位(从基部向上第3-5节)、相同部位发育成熟的叶片,带回实验室立即测定叶片相对含水量(RWC),其余样品装入密封袋,置于4℃冰箱中保存,3d内完成所有指标测定,每个指标重复测定3次。
SOD活性采用氮蓝四唑光还原法测定,POD活性采用愈创木酚法测定,MDA含量采用硫代巴比妥酸法测定,叶片相对含水量采用烘干法测定,叶绿素含量采用高俊凤等的方法[15]测定,可溶性蛋白质含量采用考马斯亮蓝G-250染色法测定。
1.3 数据处理
采用Origin9.1软件制图,用SPSS19.0中的Duncan's进行差异显著性分析。
2 结果与分析
2.1 水分胁迫对竹叶花椒生理指标的影响
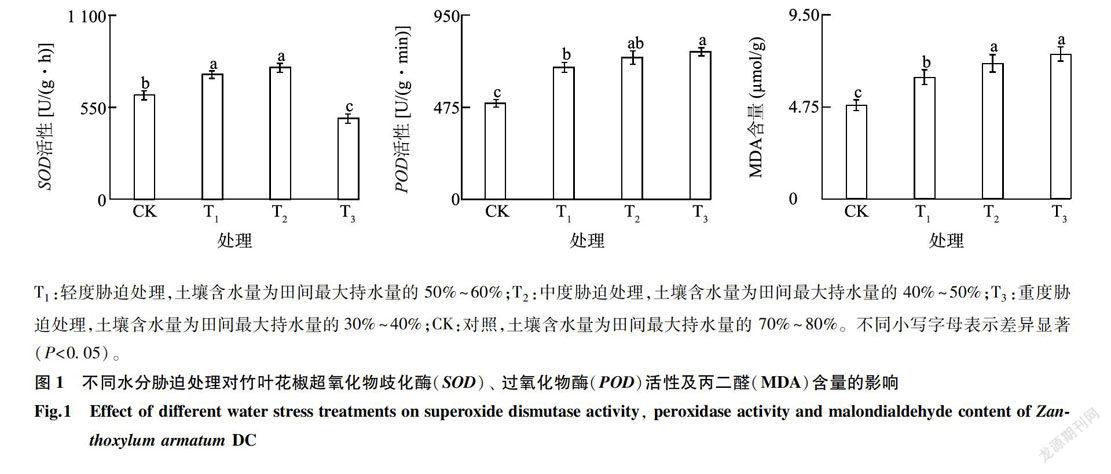
由图1可知,随着水分胁迫加深,SOD活性先升高后下降,从对照到中度胁迫,SOD活性逐渐增大,之后下降且下降幅度较大。在中度胁迫(T2)下SOD活性达到最高值785.03 U/(g.h),在重度胁迫(T3)下SOD活性达到最低值481. 97U/(g.h),与对照相比分别增加和降低26.44%、22.38%。轻度胁迫(Tl)和T2处理之间无显著差异,它们与CK、T3之间均存在显著差异。随水分胁迫加深,POD活性呈现逐渐升高的变化趋势。重度胁迫下POD活性达到762.75U/(g.min),与CK相比增加53.83%。随着土壤水分含量的降低,MDA含量逐渐增加。胁迫初期MDA含量增加速率较快,随着胁迫程度的增加,MDA含量增加趋势有所减缓,重度胁迫下丙二醛含量为7. 48 μmol/g,与CK相比增加54.55%。在水分胁迫过程中,SOD、POD活性虽有所增大,但并不能完全清除细胞组织内积累的自由基和过氧化物,因此在水分胁迫逐渐加大时MDA含量逐渐增加,但速率下降。
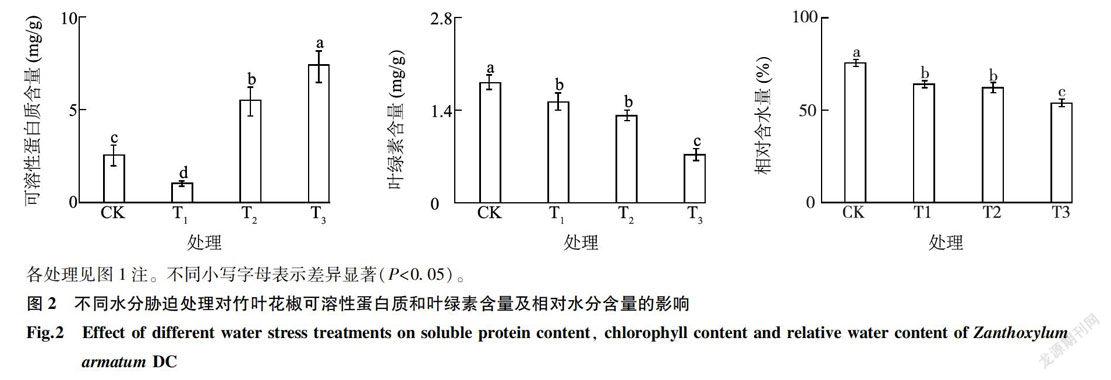
随着水分胁迫程度逐渐增加,可溶性蛋白质含量呈现增加趋势(图2)。从对照到轻度胁迫过程中,可溶性蛋白质含量下降;从轻度胁迫到重度胁迫过程中,可溶性蛋白质含量逐渐增加,且增加幅度较大。其原因可能是在胁迫初期植物体内正常的蛋白质合成被抑制,随着水分胁迫加剧进而诱导植物自身合成新的蛋白质,导致可溶性蛋白质含量增加。在水分胁迫过程中,叶绿素含量逐渐降低。胁迫初期,叶绿素含量下降幅度较小,差异不显著。重度胁迫下叶绿素含量显著下降至1 mg/g以下,原因可能是水分胁迫加速了叶绿体的降解,并且进一步抑制了叶绿素合成酶的活性,导致叶绿素合成量下降。花椒植株受到水分胁迫时,叶片相对含水量逐渐降低,对照花椒叶片相对含水量为80. 69%,T1、T2及T3处理叶片相对含水量分别为68.41%、66.43%、57.57%,与对照相比,叶片相对含水量分别下降17.95%,21.47%、40.16%,差异均达到显著水平。
2.2 竹叶花椒生理指标间的相关性分析
由表1可知,不同处理对竹叶花椒叶绿素含量的影响达显著水平,对其余5个生理指标的影响达极显著性水平。由表2可知,相对含水量(RWC)与POD活性、MDA含量呈显著负相关关系,相关系数分别为-0.967*、-0.990*( P
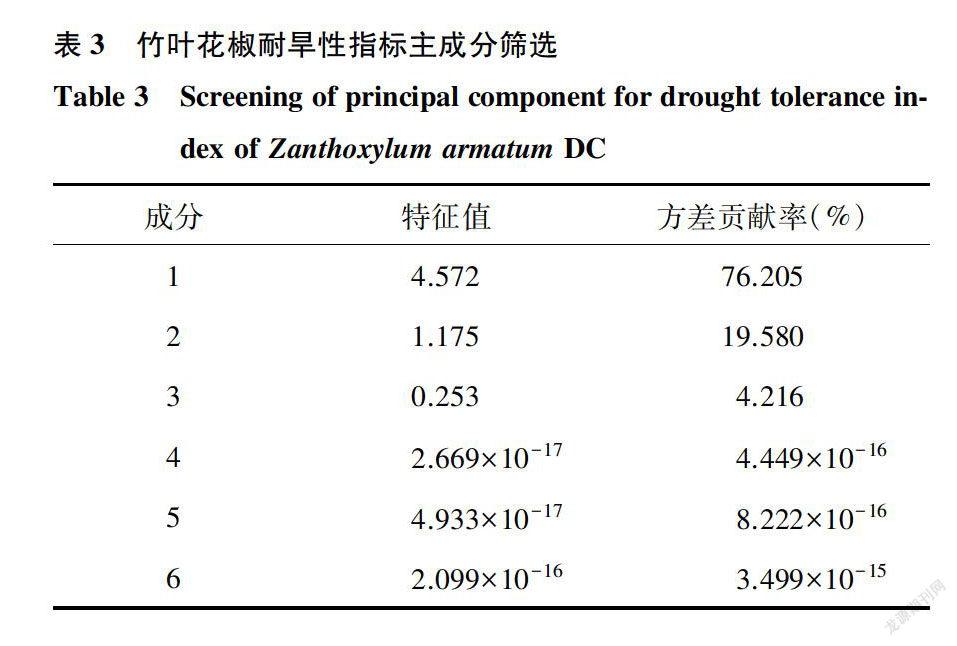
对竹叶花椒植株SOD活性、POD活性、MDA含量等6项生理指标进行主成分分析。结果(表3)表明,特征值大于l的主成分有2个,这2个主成分的累积贡献率达到95.784%。依据各生理指标在主成分1及主成分2中载荷系数绝对值大小,判断主成分包含的生理指标。第一主成分在POD活性、MDA含量、相对含水量、叶绿素含量、可溶性蛋白质含量5项指标上有较大的载荷系数,因此该主成分与这5项指标有较强的相关性:第二主成分在SOD活性上有较大的载荷系数,该主成分与SOD活性有较强的相关性(表4)。可见,POD活性、MDA含量、相对含水量、叶绿素含量、可溶性蛋白质含量这5项生理指标贡献率最大,其综合作用结果对竹叶花椒耐旱性的影响显著,可作为评价其耐旱性指标。胡曼筠等[16]对华中冬青相对电导率、丙二醛含量、可溶性蛋白质含量、可溶性糖含量、游離脯氨酸含量、超氧化物歧化酶活性、叶绿素含量及类胡萝卜素含量8项生理指标进行主成分分析,结果表明相对电导率、丙二醛含量、可溶性蛋白质含量、超氧化物歧化酶活性、叶绿素含量5项指标与华中冬青抗逆性关系密切。这与本研究结果相似,但又有所不同,可能与植物种类不同有关。
3 讨论
植物处于正常环境之中时,其细胞内自由基的产生和清除处于动态平衡状态,然而植物一旦受到不良环境侵害,这种动态平衡就会被破坏,自由基大量积累,细胞代谢紊乱,导致植物遭到伤害,而此时植物体内会迅速启动保护系统,保护酶活性被激发,催化新陈代谢产生的超氧阴离子自由基歧化为02和H202,进而缓解因不良环境胁迫而引起的细胞伤害[17-18]。本研究中,水分胁迫下SOD活性呈先升后降的趋势变化,这与张翠梅等[19]以及贾鑫等[20]的研究结果一致。SOD活性的这种变化说明竹叶花椒体内防御机制被刺激,抗性生理活动加快,SOD活性升高以应对体内超氧阴离子的增加以免细胞受损,胁迫后期可能是超出了SOD歧化能力,SOD活性下降;POD活性缓慢升高,这与刘世鹏等[21]在枣树上的研究结果相似。说明干旱胁迫发生时,竹叶花椒通过及时提高抗氧化酶活性积极适应外界环境变化,然而不同抗氧化酶的响应速度不尽相同,SOD反应迅速,其酶活性增加较快,而POD活性则是稳步上升。干旱胁迫可破坏类囊体膜结构,导致竹叶花椒叶片产生更多自由基(02和H202),使膜脂中不饱和脂肪酸过氧化,MDA等过氧化物质积累。张翠梅等[19]及潘昕等[22]的研究结果表明,水分胁迫能引起植物细胞膜破坏,导致MDA含量增加。
渗透调节物质的积累是植物适应干旱胁迫的主要方式之一。大量研究结果显示可溶性蛋白质含量与植物抗旱性呈正相关关系。本研究中,可溶性蛋白含量在胁迫前期下降,在中度和重度胁迫下,其含量逐渐升高,这与张庆华等[9]研究结果存在差异。胁迫前期可溶性蛋白含量下降的原因可能是轻度干旱胁迫下蛋白质的分解速率大于合成速率。中度和重度胁迫下可溶性蛋白质含量增加意味着在干旱响应中有新的蛋白质合成[23]。
干旱胁迫不仅会对竹叶花椒生长产生不利影响,还会对光合色素造成损害.进而影响光合能力[24]。目前关于逆境胁迫下叶绿素含量变化的研究结果尚存在一定差异。本研究中随水分胁迫加深,叶绿素含量逐渐降低,这与李阳等[25]研究结果有一定差异。叶绿素含量下降的原因可能是干旱胁迫下,在类囊体膜上产生较多的活性氧簇( ROS),这些活性氧簇可破坏光合色素。
植物叶片中相对含水量是植物重要的耐旱性能指标,在一定程度上反映叶片持水保水能力[23]。随水分胁迫加深,竹叶花椒叶片相对含水量呈逐渐下降趋势,且不同胁迫程度之间存在显著差异,与张寅媛等[8]研究结果稍有差异,这可能与叶片结构不同有关。
综上所述,水分胁迫下,竹叶花椒生理指标表现出不同的生理响应,不同胁迫程度之间呈现显著差异。胁迫初期竹叶花椒通过提高SOD、POD活性增强植株耐旱性能,合成新的可溶性蛋白质适应外界环境变化。水分胁迫引发细胞膜系统发生膜脂过氧化,导致MDA含量增加,引起叶片相对含水量降低,破坏叶绿素分子结构及叶绿体膜系统和光合反应中心。
参考文献:
[l]王纪辉,陈应福,侯娜,等,源白不同产地青花椒主要特征品质分析[J].食品工业,2018,39(8):322-325.
[2]王纪辉,梁美,侯娜.青花椒良种黔椒2号的选育及栽培技术[J].南方农业学报,2018,49(7):1383-1388.
[3] 吕小军,杨途熙,何小红,等,冬季低温对花椒抗寒性生理指标的影响[J].西北农业学报,2013,22(7):143-148.
[4] 刘淑明,孙丙寅,刘杜玲.不同种源花椒抗寒性研究[J].西北农业学报,2010,19( 11):119-124.
[5]杨途熙,魏安智,李晓,等,越冬过程中花椒抗寒性与组织水和渗透调节物质的变化[J].植物生理学通讯,2010,46(6):579-582.
[6]杨艳丽,杨勇,李大红,等,转桃PpCuZnSOD基因大豆的耐旱性[J].江苏农业学报,2018,34(5):978-983.
[7]刘杜玲,刘淑明,不同花椒品种抗旱性比较研究[J].干旱地区农业研究,2010,28(6):183-189.
[8]李雪妹,刘畅,单羽,等.PEG预处理对水分胁迫下水稻根系抗氧化酶同T酶及其表达的影响[J].江苏农业科学,2018, 46(7):54-57.
[9] 张庆华,向发云,曾祥国,等.干旱胁迫下不同品种草莓生理响应及抗旱性评价[J].北方园艺,2018(2):30-37.
[10]李佳,刘立云,李艳,等,保水剂对干旱胁迫槟榔幼苗生理特征的影响[J].南方农业学报,2018,49(1):104-108.
[11]张寅媛,刘英,白龙.干旱胁迫对4种景天科植物生理生化指标的影响[J].草业科学,2014,31(4):724-731.
[12]胡宏远,王振平.水分胁迫对赤霞珠葡萄光合特性的影响[J].节水灌溉,2016(2):18-22,27.
[13]曹岩坡,戴素英,代鹏,等,水分胁迫对不同抗旱性豇豆生长及光合特性的影响[J].河北农业科学,2018,22(1):11-14.
[14]王玉丽,孙居文,荀守华,等,干旱胁迫对东岳红光合特性、叶绿素荧光参数及叶片相对含水量的影响[J].山东农业科学,2017,49(4):46-50.
[15]高俊凤,孙群,曹翠玲.植物生理学实验指导[M].西安:陕西科技出版社,2006:29-30.
[16]胡曼筠,金晓玲,曾雯,等,华中冬青雌雄株抗寒生理指标筛选及评价[J].中南林业科技大学学报,2018,38(5):59-64.
[17] CONCEICAO V S.Regulation of chlorophyll biosynthesis and deg-radation by salt stress in sunflower levaes[J].ScientiaHorticulture ,2004, 103:93-99.
[18] JANDA T,KSA E,SZALAI G,et al.lnvestigation of antioxidantactivity in mize during low temperature stress[J].Acta BiologicaSzegedrensis, 2005 ,49( 1/2): 53-54.
[19]张翠梅,师尚礼,吴芳,干旱胁迫对不同抗旱性苜蓿品种根系生长及生理特性影响[J].中国农业科学,2018,51(5):868-882。
[20]贾鑫,孙窗舒,李光跃,等,干旱胁迫对蒙古黄芪生长和生理生化指标及其黄芪甲苷积累的影响[J].西北植物学报,2018,38(3):501-509.
[21]劉世鹏,刘济明,陈宗礼,等.模拟干旱胁迫对枣树幼苗的抗氧化系统和渗透调节的影响[J].西北植物学报,2006,26(9): 1781-1787.
[22]潘昕,邱权,李吉跃,等.干旱胁迫对青藏高原6种植物生理指标的影响[J].生态学报,2014,34( 13):3558-3567.
[23]冯晓敏,张永清.水分胁迫对糜子植株苗期生长和光合特性的影响[J].作物学报,2012,38(8):1513-1521.
[24]王婷,海梅荣,罗海琴,等.水分胁迫对马铃薯光合生理特性和产量的影响[J].云南农业大学学报,2010,25(5):737-742.
[25]李阳,齐曼·尤努斯,祝燕,水分胁迫对大果沙枣光合特性及生物量分配的影响[J].西北植物学报,2006,26( 12): 2493 -2499.
(责任编辑:张震林)
猜你喜欢
农业研究与应用(2017年2期)2017-07-01 18:51:07
天津农业科学(2017年6期)2017-06-10 20:17:46
江苏农业科学(2017年7期)2017-05-23 08:41:04
江苏农业科学(2016年1期)2017-05-17 11:45:21
科技创新与应用(2017年1期)2017-05-11 09:03:04
江苏农业科学(2016年10期)2017-02-05 14:52:07
江苏农业科学(2016年3期)2016-05-03 15:31:15
安徽农学通报(2015年12期)2015-07-09 11:06:39
长江蔬菜·学术版(2015年2期)2015-04-27 13:29:40
江苏农业科学(2014年10期)2014-11-22 05:24:29
