
醋栗叶片对干旱胁迫的光合生理生化响应
2019-09-10 07:22:44古丽江·许库尔汗孙雅丽巴哈尔古丽·阿优甫哈尔肯·叶尔江阿依古丽·铁木儿
南方农业学报 2019年7期
古丽江·许库尔汗 孙雅丽 巴哈尔古丽·阿优甫 哈尔肯·叶尔江阿依古丽·铁木儿

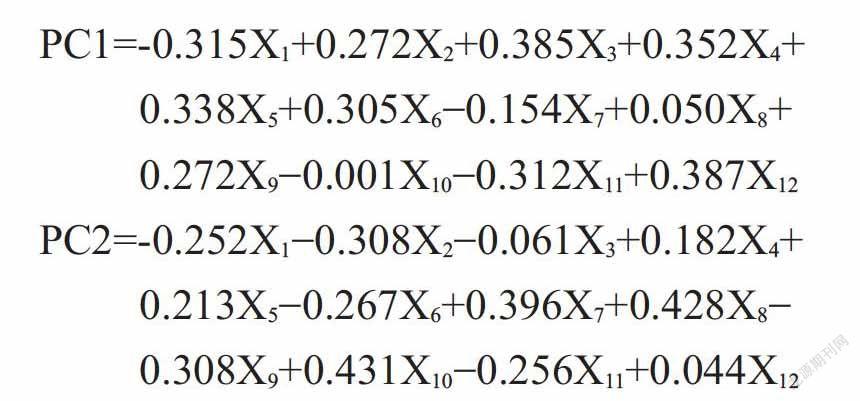

摘要:【目的】分析醋栗(Gooseberry)幼苗叶片对干旱胁迫的光合生理生化响应,为小浆果果树抗旱育种提供理论依据。【方法】采用盆栽自然干旱法控水模拟干旱,对彩虹、坠玉和红灯笼3个醋栗品种的2年生苗进行4个梯度胁迫处理(CK、土壤水含量35%、30%和25%),分析各处理的可溶性糖(SS)、脯氨酸(Pro)和丙二醛(MDA)含量、相对电导率(REC)及超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性,运用主成分分析法对其抗旱性进行综合评估。【结果】干旱胁迫导致3个醋栗品种的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和水分利用效率(WUE)不同程度下降,胞间CO2浓度(Ci)上升,其中坠玉和红灯笼的WUE下降较明显,彩虹和红灯笼的Gs显著下降(P<0.05,下同)。随干旱胁迫程度的加剧,坠玉、彩虹和红灯笼的SS、Pro、MDA含量和REC均显著增加;坠玉和彩虹的POD活性在重度干旱胁迫时保持较高水平;3个醋栗品种的SOD活性均降至低于CK水平;中度和重度干旱胁迫时红灯笼的CAT活性最高,其次为坠玉。主成分分析结果表明,3个醋栗品种的抗旱性排序为彩虹(1.886)>坠玉 (-0.289)>红灯笼(-1.596)。【结论】醋栗品种彩虹、坠玉和红灯笼的光合和生理生化指标对干旱胁迫均表现出明显的响应,但对干旱胁迫的耐受能力和适应能力存在差异,其抗旱性排序为彩虹>坠玉>红灯笼。
关键词: 醋栗;干旱胁迫;生理生化指标;抗旱性评价
0 引言
【研究意义】醋栗(Gooseberry)别称灯笼果,为虎耳草科(Saxifragaceae)茶藨子亚科醋栗属(Grossularia Mill.)落叶灌木,主要分布于我国吉林(长白山)、黑龙江(小兴安岭)、河北、山西和新疆(阿尔泰山)(李芳晓等,2014),南方省份也有引种栽培(王瑞等,2016),植株高0.8~1.2 m,小枝密,有刺毛和短刺,果实呈近球形或椭圆形,果实上有纵向维管束,成熟果实黄绿色、紫色或深红色。醋栗具有较高的药用和保健价值,其果实能加工高档果酒、果醋和果酱,是新型高经济价值小浆果。我国西北干旱半干旱地区降水量少,干旱引起的蒸发量日趋增强,严重困扰西北地区农林牧业的发展(Chaves et al.,2003;李方安等,2013)。近年来,新疆林果业得到较快发展,但北疆地区推广应用的醋栗等小浆果类果树品种抗旱性存在明显差异,抗旱性差的品种果实小或产量低,严重制约了小浆果产业的健康持续发展。因此,引种醋栗优良品种并分析其幼苗叶片光合生理生化对干旱胁迫的响应,对挖掘干旱、半干旱地区小浆果产业潜力及小浆果抗旱育种具有重要意义。【前人研究进展】郑海燕(2009)研究认为,干旱胁迫会影响林果业的产量和果实品质,甚至造成大片果树死亡;果树的抗旱性研究主要在形态结构、生理生化机制及基因工程等方面取得一定进展,揭示了果树抗旱机理,为小浆果类果树抗旱特性研究奠定了基础。张壮飞(2012)研究發现,黑穗醋栗在高温胁迫下能产生自我保护机制,在胁迫初期抗氧化酶活性升高,游离脯氨酸(Pro)含量增加,但不同品种存在明显差异。热依扎·朱木斯别克(2013)研究表明,轻度干旱胁迫(土壤水含量为60%)和中度干旱胁迫(土壤水含量为40%)黑穗醋栗品种世纪光和世纪星30 d,其叶片Pro含量增加,超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性升高,净光合速率(Pn)、蒸腾速率(Tr)和水分利用效率(WUE)的日变化均呈双峰曲线。杨玉海等(2013)研究发现,干旱胁迫下植物叶片的水势降低,并通过调节自身生理状态以提高吸水能力。田浩(2014)研究表明,植物遇到干旱胁迫时耗水量大于吸水量,发生水分亏缺,植物体内正常生理活动受到干扰。简营(2016)研究发现,逆境胁迫下植物细胞膜遭到破坏,膜透性加大,电解质外渗,电导率升高,细胞内外平衡失调,使活性氧大量积累,引发膜质过氧化,导致膜系统受损。何亚南(2017)研究认为,叶片是植物的主要光合作用和蒸腾作用器官,也是植物外部形态中对环境反应最敏感的器官,当植株遇到干旱时,叶片最先受损,出现萎蔫和枯死等现象。杜兴良等(2018)研究表明干旱和高温复合胁迫导致小麦叶片SOD和CAT活性降低。【本研究切入点】目前,鲜见综合分析醋栗品种在干旱胁迫下其体内可溶性糖(SS)和Pro等渗透调节物质含量和保护酶活性与其抗旱性关系的研究报道。【拟解决的关键问题】分析3个醋栗品种对干旱胁迫的光合生理生化响应和适应性机制,综合评价其抗旱性,为抗旱醋栗品种选育及抗旱小浆果品种栽培技术研发提供参考依据。
1 材料与方法
1. 1 试验材料
试验材料为2年生坠玉(Zhuiyu)、彩虹(Caihong)和红灯笼(Hongdenglong)醋栗苗,均从黑龙江省尚志市引进。采用盆栽方法将3个醋栗品种种苗定植于高28.0 cm、口径32.0 cm的塑料花盆中,每盆栽1株。试验用土按森林土∶园土=4∶1混合而成,每盆装土4.0 kg。
1. 2 试验方法
1. 2. 1 试验设计 试验于2017—2018年在新疆林业科学院树木园试验地进行。2017年10月底釆用盆栽自然干旱法,人工模拟干旱胁迫3个醋栗品种种苗。为防止参试材料被雨淋,下雨前用塑料布覆盖。2018年春季,在各品种醋栗苗萌芽前,选留离地面高25~30 cm枝条进行常规管理(其余部分进行平茬)。参照贺康宁等(2003)的方法,利用烘干法测得土壤绝对水含量约44%;设4个土壤干旱胁迫梯度处理,分别为:土壤绝对水含量40%(对照,CK),盆中土壤重量降至3.6~3.7 kg;土壤绝对水含量35%(轻度胁迫),盆中土壤重量降至3.3~3.4 kg;土壤绝对水含量30%(中度胁迫),盆中土壤重量降至3.0~3.1 kg;土壤绝对水含量25%(重度胁迫),盆中土壤重量降至2.6~2.7 kg。2018年6月28日将各品种醋栗盆栽苗一次性浇透水,让其自然干旱。待各盆栽苗土壤重量降至预设重量时,开始测定盆栽苗各项光合参数及生理指标。每处理3盆,每次随机在各盆醋栗苗中上部取2~3片功能叶进行测定,重复3次。
1. 2. 2 测定指标及方法 Pn、Tr、胞间CO2浓度(Ci)和气孔导度(Gs)采用CIRAS-Ⅱ型便携式光合作用测定系统(美国PPsystems公司)进行测定,并计算WUE。测定时间为上午10:00—12:00,有效辐射强度为1200 mol/(m2·s);SS和Pro含量采用蒽酮比色法(邹琦,2004)进行测定,相对电导率(REC)采用电导率法(郑炳松,2006)进行测定,丙二醛(MDA)含量采用硫代巴比妥酸法(刘家尧,2010)进行测定,SOD、POD和CAT活性参照田浩(2014)的方法进行测定。
1. 3 统计分析
试验数据采用Excel 2003进行整理,以SPSS 16.0进行差异显著性分析,采用主成分分析法对3个醋栗品种的抗旱性进行综合评价。
2 结果与分析
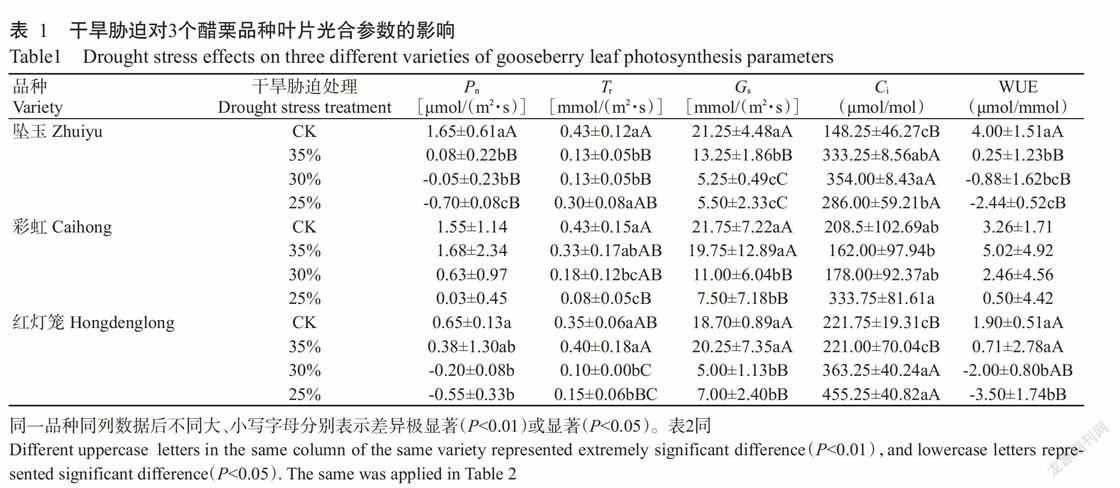
2. 1 干旱胁迫对醋栗叶片主要光合参数的影响
2. 1. 1 对Pn的影响 由表1可知,坠玉叶片的Pn在轻度干旱胁迫时降至0.08 μmol/(m2·s),中度干迫胁迫时降至-0.05 μmol/(m2·s),重度干旱胁迫时降至 -0.70 μmol/(m2·s),均极显著低于CK(P<0.01,下同);彩虹叶片的Pn在轻度干旱胁迫时[1.68 μmol/(m2·s)]稍高于CK,在中度和重度干旱胁迫时分别降至0.63和0.03 μmol/(m2·s),但与CK均无显著差异(P>0.05,下同);红灯笼叶片的Pn在轻度干旱胁迫时降至0.38 μmol/(m2·s),与CK差异不显著,在中度和重度干旱胁迫时分别降至-0.20和-0.55 μmol/(m2·s),与轻度干旱胁迫的Pn差异不显著,但均显著低于CK(P<0.05,下同)。在不同程度干旱胁迫下,3个醋栗品种叶片的Pn总体上呈下降趋势,但彩虹叶片的Pn始终高于其余2个品种,表现出较强的耐旱性。
2. 1. 2 对Tr的影响 从表1可看出,坠玉叶片的Tr在轻度和中度干旱胁迫时降至0.13 mmol/(m2·s),极显著低于CK,在重度干旱胁迫时升至0.30 mmol/(m2·s),低于CK,差异不显著,但显著高于轻度和中度干旱胁迫;彩虹叶片的Tr在轻度干旱胁迫时降至0.33 mmol/(m2·s),与CK差异不显著,在中度干旱胁迫时降至0.18 mmol/(m2·s),显著低于CK,在重度干旱胁迫时降至0.08 mmol/(m2·s),极显著低于CK;红灯笼叶片的Tr在轻度胁迫时升至0.40 mmol/(m2·s),与CK差异不显著,在中度干旱胁迫时降至0.10 mmol/(m2·s),与CK差异极显著,在重度干旱胁迫时降至0.15 mmol/(m2·s),与CK差异极显著。可见,干旱胁迫期间3个醋栗品种的Tr总体上呈下降趋势,其叶片通过减少水分蒸腾和气体交换阻止叶片水势下降,但重度干旱胁迫时坠玉叶片的Tr高于其余品种,表现出较强的耐旱性。
2. 1. 3 对Gs的影响 由表1可知,坠玉叶片的Gs在轻度干旱胁迫时降至13.25 mmol/(m2·s),在中度和重度干旱胁迫时分别降至5.25和5.50 mmol/(m2·s),極显著低于CK;彩虹叶片的Gs在轻度干旱胁迫时降至19.75 mmol/(m2·s),与CK差异不显著,在中度和重度干旱胁迫时分别降至11.00和7.50 mmol/(m2·s),极显著低于CK;红灯笼叶片的Gs在轻度干旱胁迫时[20.25 mmol/(m2·s)]稍高于CK,但差异不显著;在中度和重度干旱胁迫时分别降至5.00和7.00 mmol/(m2·s),均极显著低于CK。说明干旱胁迫使3个醋栗品种叶片的Gs总体上呈下降趋势,叶片通过关闭气孔阻止其体内水分蒸发,但彩虹叶片的Gs在重度干旱胁迫时高于其余2个品种,表现出较强的耐旱性。
2. 1. 4 对Ci的影响 从表1可看出,坠玉叶片的Ci在轻度干旱胁迫时升至333.25 μmol/mol,在中度和重度干旱胁迫时分别升至354.0和286.0 μmol/mol,均极显著高于CK;彩虹叶片的Ci在轻度和中度干旱胁迫时分别降至162.0和178.00 μmol/mol,与CK无显著差异,在重度干旱胁迫时升至333.75 μmol/mol,高于CK但差异不显著;红灯笼叶片的Ci在轻度干旱胁迫时降至221.0 μmol/mol,与CK无显著差异,在中度和重度干旱胁迫时分别升至363.25和455.25 μmol/mol,极显著高于CK。可见,干旱胁迫期间3个醋栗品种叶片的Ci总体上呈上升趋势,但轻度干旱胁迫时彩虹和红灯笼叶片的Ci有下降趋势(Pn也稍有下降),是由气孔因素所引起;重度干旱胁迫时3个醋栗品种叶片的Ci上升(同时Pn均下降),是由非气孔因素引起。
2. 1. 5 对WUE的影响 从表1可看出,坠玉叶片的WUE在轻度干旱胁迫时降至0.25 μmol/mmol,在中度和重度干旱胁迫时分别降至-0.88和-2.44 μmol/mmol,均极显著低于CK;彩虹叶片的WUE在轻度干旱胁迫时升至5.02 μmol/mmol,在中度干旱胁迫时降至2.46 μmol/mmol,在重度干旱胁迫时降至0.50 μmol/mmol,与CK均无显著差异;红灯笼叶片的WUE在轻度干旱胁迫时降至0.71 μmol/mmol,与CK差异不显著,在中度干旱胁迫时降至-2.00 μmol/mmol,显著低于CK,在重度干旱胁迫时降至-3.50 μmol/mmol,极显著低于CK。说明干旱胁迫期间3个醋栗品种叶片气孔开度减小,水分散发少,WUE提高,其中彩虹叶片的WUE高于其余2个品种。
2. 2 干旱胁迫对不同醋栗品种叶片生理指标的影响
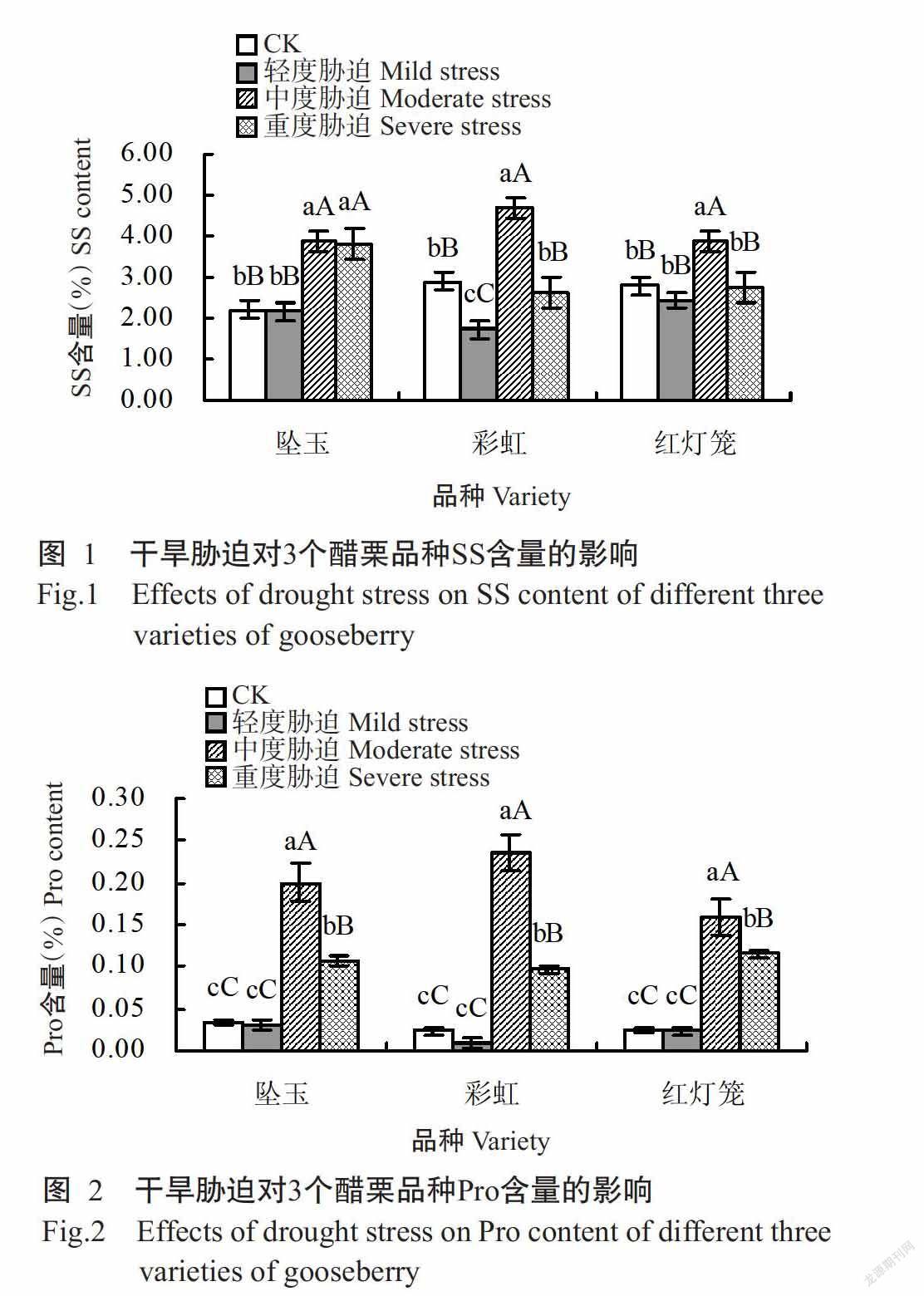
2. 2. 1 对SS含量的影响 从图1可看出,坠玉叶片的SS含量在轻度干旱胁迫时降至2.17%,与CK无显著差异,在中度和重度干旱胁迫时分别升至3.89%和3.81%,极显著高于CK;彩虹叶片的SS含量在轻度干旱胁迫时降至1.73%,极显著低于CK,在中度干旱胁迫时升至4.67%,极显著高于CK,在重度干旱胁迫时迅速降至2.64%,低于CK但差异不显著;红灯笼叶片的SS含量在轻度干旱胁迫时降至2.45%,与CK差异不显著,在中度干旱胁迫时升至3.89%,极显著高于CK,在重度干旱胁迫时降至2.73%,低于CK但差异不显著。说明彩虹叶片的SS含量在中度干旱胁迫时高于其余2个品种,即其对中度干旱胁迫具有较强的耐受性。
2. 2. 2 对Pro含量的影响 从图2可看出,坠玉叶片的Pro含量在轻度干旱胁迫时与CK相同,均为0.03%,在中度干旱胁迫时升至0.20%,极显著高于CK,在重度干旱胁迫时降至0.11%,仍极显著高于CK;彩虹叶片的Pro含量在轻度干旱胁迫时降至0.01%,与CK差异不显著,在中度干旱胁迫时升至0.24%,极显著高于CK,在重度干旱胁迫时降至0.10%,仍极显著高于CK;红灯笼叶片的Pro含量在轻度干旱胁迫时降至0.02%,与CK差异不显著,在中度和重度干旱胁迫时分别升至0.16%和0.12%,极显著高于CK。可见,在中度干旱胁迫时彩虹叶片的Pro含量明显高于其他2个品种,在重度干旱胁迫时3个品种的Pro含量相近,即3个醋栗品种通过积累Pro,自身的抗旱性均得以增强。
2. 2. 3 对MDA含量的影响 从图3可看出,坠玉叶片的MDA含量在轻度干旱胁迫时降至0.022 µmol/gFW,极显著低于CK,在中度干旱胁迫时升至0.027 µmol/gFW,与CK持平,在重度干旱胁迫时升至0.074 µmol/gFW,极显著高于CK;彩虹叶片的MDA含量在轻度和中度干旱胁迫时分别降至0.033和0.026 µmol/gFW,显著低于CK,在重度干旱胁迫时升至0.067 µmol/gFW,极显著高于CK;红灯笼叶片的MDA含量在轻度和中度干旱胁迫时分别降至0.029和0.028 µmol/gFW,显著低于CK,在重度干旱胁迫时升至0.074 µmol/gFW,极显著高于CK。说明随干旱胁迫强度的增加,3个醋栗品种叶片的MDA含量总体上呈增加趋势,质膜透性逐渐增大,其中轻度和中度干旱胁迫的MDA含量和质膜透性增幅相对较小,而重度干旱胁迫时的MDA含量增幅较大,质膜透性也相应增大,即重度干旱胁迫对3个醋栗品种细胞膜的伤害均重于轻度和中度干旱胁迫,但对彩虹叶片细胞膜的伤害程度小于其余品种。
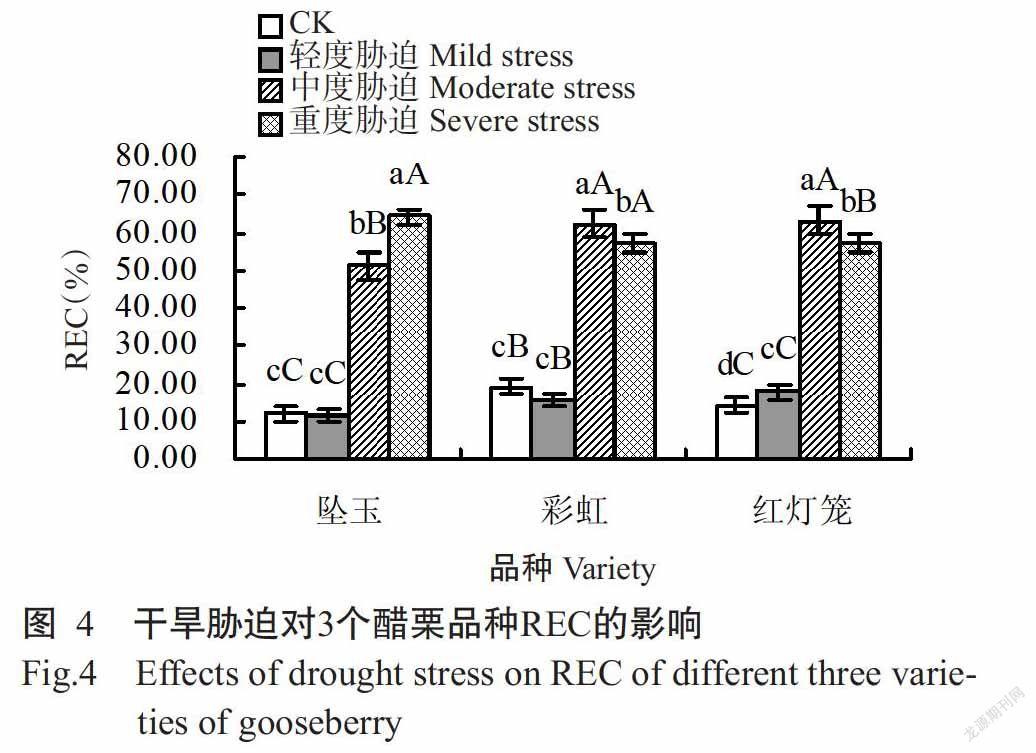
2. 2. 4 对REC的影響 由图4可看出,坠玉叶片的REC在轻度干旱胁迫时降至11.57%,与CK差异不显著,在中度和重度干旱胁迫时分别升至51.25%和64.15%,极显著高于CK;彩虹叶片的REC在轻度干旱胁迫时降至15.57%,与CK差异不显著,在中度和重度干旱胁迫时分别升至62.44%和57.37%,极显著高于CK;红灯笼叶片的REC在轻度干旱胁迫时升至17.7%,与CK差异显著,在中度和重度干旱胁迫时分别升至63.13%和57.26%,与CK差异极显著。重度干旱胁迫时坠玉叶片的REC稍高于彩虹和红灯笼,即重度干旱胁迫时坠玉叶片的细胞膜完整性较彩虹和红灯笼稍差。
2. 3 干旱胁迫对3个醋栗品种叶片抗氧化酶活性的影响
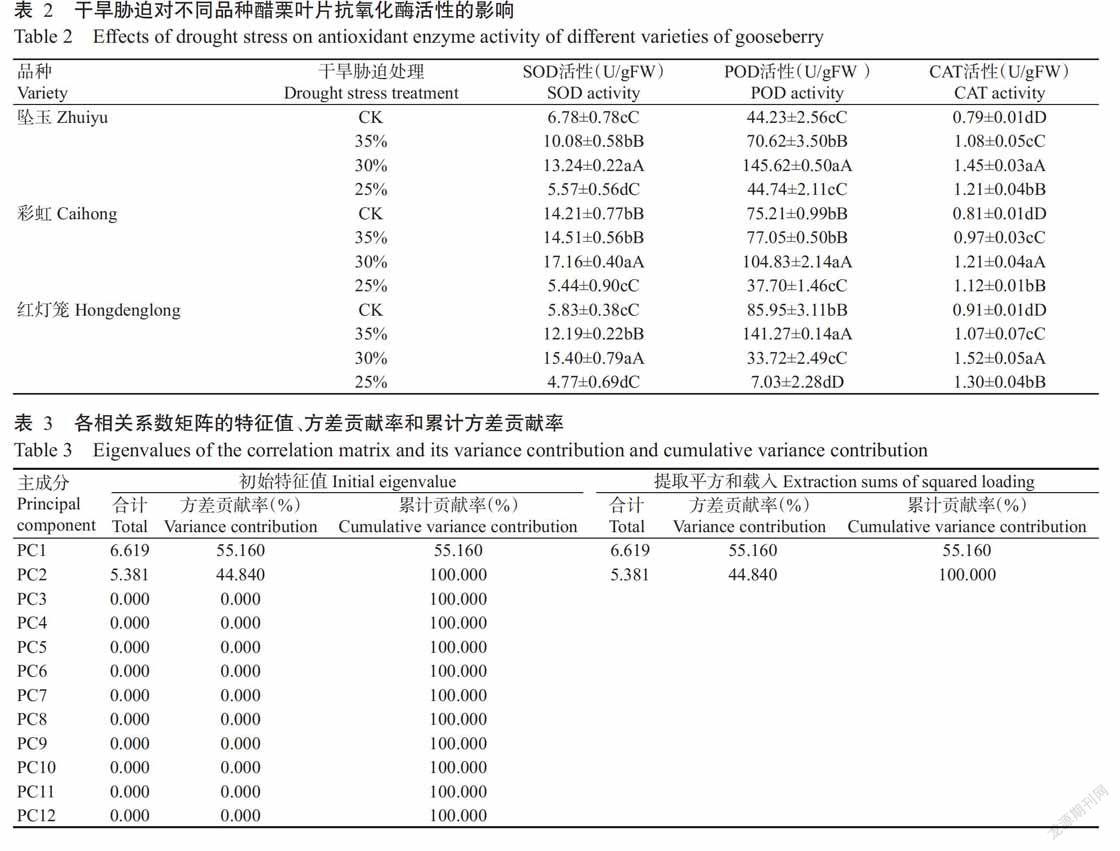
2. 3. 1 对SOD活性的影响 由表2可知,坠玉叶片的SOD活性在轻度和中度干旱胁迫时分别为10.08和13.24 U/gFW,极显著高于CK,在重度干旱胁迫时降至5.57%,显著低于CK;彩虹叶片的SOD活性在轻度干旱胁迫时升至14.51 U/gFW,与CK差异不显著,在中度干旱胁迫时升至17.16 U/gFW,极显著高于CK,在重度干旱胁迫时降至5.44 U/gFW,极显著低于CK;红灯笼叶片的SOD活性在轻度和中度干旱胁迫时分别为12.19和15.40 U/gFW,极显著高于CK,在重度干旱胁迫时降至4.77 U/gFW,显著低于CK。由表2还可看出,在干旱胁迫期间3个醋栗品种叶片的SOD活性总体上均呈先上升后下降的变化趋势,其中,中度干旱胁迫的SOD活性均极显著高于CK,尤其是彩虹叶片的SOD活性最高,说明中度干旱胁迫促使醋栗叶片产生大量SOD以清除自由基,保护细胞膜免受活性氧胁迫的伤害,而重度干旱胁迫使醋栗叶片的SOD活性显著或极显著下降,表明其细胞膜已受到活性氧胁迫的伤害,但彩虹细胞膜受到的伤害小于其余2个品种。
2. 3. 2 对POD活性的影响 由表2可知,坠玉叶片的POD活性在轻度和中度干旱胁迫时分别为70.62和145.62 U/gFW,极显著高于CK,在重度干旱胁迫时降至44.74 U/gFW,高于CK但差异不显著;彩虹叶片的POD活性在轻度干旱胁迫时升至77.05 U/gFW,与CK差异不显著,在中度干旱胁迫时升至104.83 U/gFW,极显著高于CK,在重度干旱胁迫时降至37.7 U/gFW,极显著低于CK;红灯笼叶片的POD活性在轻度干旱胁迫时升至141.27 U/gFW,极显著高于CK,在中度和重度干旱胁迫时分别降至33.72和7.03 U/gFW,极显著低于CK。由表2还可看出,干旱胁迫期间3个醋栗品种叶片的POD活性总体上呈先上升后下降的变化趋势,尤其在轻度干旱胁迫时红灯笼叶片的POD活性(141.27 U/gFW)明显高于坠玉和彩虹,即红灯笼叶片较坠玉和彩虹叶片更易受轻度干旱胁迫而衰老;在中度和重度干旱胁迫时坠玉叶片的POD活性(分别为145.62和44.74 U/gFW)明显高于彩虹和红灯笼,即坠玉叶片较彩虹和红灯笼更易受中度和重度干旱胁迫而衰老。可见,彩虹的耐干旱胁迫性相对强于坠玉和红灯笼。
2. 3. 3 对CAT活性的影响 由表2可知,在干旱胁迫期间3个醋栗品种叶片的CAT活性总体上呈先上升后下降的变化趋势,但均极显著高于CK。其中,轻度、中度和重度干旱胁迫下坠玉叶片的CAT活性分别为1.08、1.45和1.21 U/gFW,分别比CK提高26.9%、45.5%和34.7%;轻度、中度和重度干旱胁迫下彩虹叶片的CAT活性分别为0.97、1.21和1.12 U/gFW,分别比CK提高16.5%、33.1%和27.7%;轻度、中度和重度干旱胁迫下红灯笼叶片的CAT活性分别为1.07、1.52和1.30 U/gFW,分别较CK提高14.9%、40.1%和30.0%。从表2还可看出,轻度干旱胁迫时3个醋栗品种叶片的CAT活性排序为坠玉>红灯笼>彩虹,中度和重度干旱胁迫时3个醋栗品种的CAT活性排序均为红灯笼>坠玉>彩虹。说明彩虹对干旱胁迫的耐受能力强于红灯笼和坠玉,抗旱性更强。
2. 4 3个醋栗品种抗旱性的综合评价
2. 4. 1 抗旱指标的主成分分析 由表3可知,前2个主成分已能代表醋栗品种12个抗旱指标(Pn、Tr、Ci、Gs、WUE、SS和Pro含量、REC、MDA含量及SOD、POD和CAT活性)的大部分信息。因此,可将不同醋栗品种的12个抗旱指标综合成2个主成分。
由各抗旱指标在2个主成分中的载荷(表4)可知,第一主成分(PC1)主要代表Ci、Tr、Gs、Pn、WUE、REC、CAT和SOD活性的信息;第二主成分(PC2)主要代表SS、Pro、MDA含量和POD活性的信息。
2. 4. 2 不同醋栗品种抗旱性的综合评价结果 通过主成分分析,提取出2个主成分(λ>1),用主成分载荷矩阵中的数据计算特征向量式中,Bi为主成分载荷矩阵,SQR为平方根函数,λ为特征值],得出的特征向量矩阵如表5所示,各特征向量与标准化后的数据相乘,得到各主成分的表达式。
以每个主成分的特征值占所提取主成分总特征值的比例为权重,计算主成分综合模型F=6.619F1+5.381F2。运用此数学模型对不同醋栗品种抗旱性进行综合评价及排序。综合得分越高,表明该样品的抗旱性越强。由表6可知,3个醋栗品种抗旱性综合得分排序为彩虹>坠玉>红灯笼。说明彩虹对干旱胁迫的耐受能力和适应能力最强,坠玉次之,红灯笼稍弱。
3 讨论
3. 1 干旱胁迫下醋栗光合参数的变化
已有研究表明,植物在干旱胁迫下其Pn降低(Jerry et al.,2001,魏孝荣等,2005),抗旱性强的植物Pn降低程度小于抗旱性弱的植物(Ao and Zhang,2007),即抗旱性较强植物的Pn较高,因此,Pn可作为判断植物抗旱性的可靠指标(周忆堂等,2008;吉小敏等,2012)。本研究中,彩虹叶片的Pn高于坠玉和红灯笼,说明彩虹的抗旱性强于坠玉和红灯笼。Jerry等(2001)研究认为,干旱胁迫下拥有较高WUE的植物具有更高生产力和生存能力。魏孝荣等(2005)研究发现,叶片的WUE是评价植物耐旱能力的重要指標之一,该指标受Tr和Pn的共同影响。本研究结果与Jerry等(2001)、魏孝荣等(2005)的研究结果相似,坠玉和红灯笼的WUE在中度干旱胁迫时降为负值(分别为-0.88和-2.00 μmol/mmol),而彩虹叶片的WUE虽然降低,但仍维持正值(2.46 μmol/mmol),明显高于坠玉和红灯笼,说明彩虹较坠玉和红灯笼具有更强的耐旱能力。本研究还发现,在干旱胁迫过程中3个醋栗品种叶片的Pn、Tr、Gs和WUE总体上呈下降趋势,Ci呈上升趋势,与迟琳琳(2017)对科尔沁沙地小叶鼠李、叶底珠、东北连翘和杠柳4种灌木的研究结果相似。虽然坠玉叶片的Ci在中度干旱胁迫时升至最高,但在重度干旱胁迫时呈下降趋势,可能是随着干旱胁迫的加剧,植株消耗的CO2增多,Ci被消耗后得不到及时补充所致。
3. 2 干旱胁迫下醋栗生理生化指标的变化
SS是植物渗透调节中的重要有机溶质,随着干旱胁迫的持续,植物体能通过自身代谢调控积累一些有机小分子物质以调节渗透胁迫,SS含量上升(许翩翩和王建柱,2018),细胞原生质浓度、细胞膜的稳定性和植株的保水性得到提高,从而增强植物的抗旱能力(姜英淑等,2009)。本研究中,中度干旱胁迫时彩虹叶片的SS含量显著高于CK,且明显高于其余2个品种,但在重度干旱胁迫时降至与CK水平相近,可能是干旱胁迫程度超出彩虹叶片忍耐范围,致使其SS含量降至接近CK水平的缘故;重度干旱胁迫时坠玉叶片的SS含量高于彩虹和红灯笼,说明不同品种对干旱胁迫程度的耐受能力存在差异。
已有研究认表明,干旱胁迫下植物的SS含量增加到一定程度后会诱发Pro大量生成(田浩,2014),而Pro的大量积累能防止水分散失,直接影响蛋白的稳定性,有利于植物抗旱(郑海燕,2009),且干旱胁迫越严重,Pro含量越高,即在干旱胁迫下Pro积累越多的植物其抗旱性越强(Yamada et al.,2005)。本研究中,中度干旱胁迫时彩虹叶片的Pro含量明显高于其他2个品种,且均极显著高于CK,重度干旱胁迫时3个品种的Pro含量相近,但均极显著高于CK,说明3个醋栗品种通过积累Pro,使自身的抗旱性得以增强。
MDA对逆境反应非常敏感,在发生干旱胁迫时,植物细胞膜的透性增大,膜系统发生膨胀、破损(田浩,2014),因此,其含量与植物细胞膜受损程度呈正相关,MDA含量越高,植物受干旱胁迫损害越大,抗旱性越弱(时连辉等,2005;姜英淑等,2009)。本研究中,3个醋栗品种受到干旱胁迫时体内产生大量膜脂过氧化物,MDA含量增加并超出醋栗叶片的耐受范围,致使其细胞膜系统受到损害,电解质外渗,REC升高。但由于叶片含有防止膜脂过氧化的POD和SOD等保护酶类,其活性在干旱胁迫下得以增强,有效降低脂膜过氧化产物MDA含量,因此彩虹和红灯笼在中度干旱胁迫时MDA含量降低;随干旱胁迫的加剧,各醋栗品种保护酶系统的活性被钝化或降低(李叶峰等,2011),不能发挥正常功能,所以重度干旱胁迫导致各醋栗品种MDA含量升至最高。在干旱胁迫期间3个醋栗品种的REC变化趋势与MDA含量变化趋势基本相同,但在重度胁迫时出现彩虹和红灯笼叶片的REC小幅下降情况,与曹建华等(2014)对油棕幼苗的研究结果相似。
3. 3 干旱胁迫下醋栗酶活性的变化
本研究中,3个醋栗品种的SOD活性均在中度干旱胁迫时升至最高,其中彩虹为17.16 U/gFW、红灯笼为15.40 U/gFW、坠玉为13.20 U/gFW,在重度干旱胁迫时极显著降低,说明彩虹叶片SOD活性增加可清除过多的活性氧,降低其叶片细胞膜的损害程度,与徐建欣等(2014)研究认为干旱胁迫下陆稻可通过提高抗氧化酶活性、增强抗氧化能力以减轻干旱胁迫伤害的观点一致。3个醋栗品种中,红灯笼的POD活性在轻度干旱胁迫时最高,在中度干旱胁迫时下降,说明轻度干旱胁迫时红灯笼叶片的细胞为适应干旱随即产生保护反应,导致清除活性氧能力降低;坠玉和彩虹的POD活性在中度干旱胁迫时出现高峰(其中坠玉的POD活性高于彩虹),在重度干旱胁迫时下降,说明随着干旱胁迫的加剧,坠玉和彩虹叶片细胞中自由基的产生和清除平衡失调,导致SOD和POD活性下降,与Gill和Tuteja(2010)对金心吊兰的研究结果相似。3个醋栗品种中,彩虹叶片的CAT活性下降程度低于其他2个品种,表明彩虹的耐旱能力强于坠玉和红灯笼,与韩建秋(2009)对白三叶的研究结果一致。
本研究结果表明,干旱胁迫导致彩虹、坠玉和红灯笼叶片的Pn、Tr、Gs和WUE不同程度下降, Ci总体上上升。其中,坠玉和红灯笼的WUE和Gs下降最显著;随干旱胁迫的加剧,坠玉、彩虹和红灯笼叶片的SS、Pro、MDA含量和REC均增加,彩虹叶片的SS和Pro含量增加最明显,红灯笼叶片的SS和Pro含量增幅较坠玉小,说明彩虹对干旱胁迫的耐受能力最强,而红灯笼对干旱胁迫的耐受能力最弱。
4 结论
醋栗品种彩虹、坠玉和红灯笼的光合和生理生化指标对干旱胁迫均表现出明显的响应,但对干旱胁迫的耐受能力和适应能力存在差异,其抗旱性排序为彩虹>坠玉>红灯笼。
参考文献:
曹建华,李静,陶忠良,陈俊明,谢贵水. 2014. 油棕幼苗对低温胁迫的生理响应及其抗寒力评价[J]. 热带农业科学,34(8):8-12. [Cao J H,Li J,Tao Z L,Chen J M,Xie G S. 2014. Physiological responses of oil palm seedlings to low temperature stress and cold hardiness evaluation[J]. Chinese Journal of Tropical Agriculture, 34(8):8-12.]
迟琳琳. 2017. 科尔沁沙地4种灌木对干旱胁迫的生理响应[J]. 干旱区资源与环境,31(5):158-162. [Chi L L. 2017. The physiological response to drought stress for four landscape-economical shrubs in Horqin sandy land[J]. Journal of Arid Land Resources and Environment,31(5):158-162.]
杜兴良,兰盼龙,张皓帆,赵光伟,李华,汪月霞,赵会杰. 2018. 一氧化氮对高温与干旱复合胁迫下小麦叶片Rca基因表达及Rubisco活性的影响[J]. 河南农业大学学报,52(6):868-873. [Du X L,Lan P L,Zhang H F,Zhao G W,Li H,Wang Y X,Zhao H J. 2018. Regulating effect of nitric oxide on expression of Rca gene and activity of Rubisco in wheat leaves under combined stress of heat and drought[J]. Journal of Henan Agricultural University,52(6):868-873.]
韩建秋. 2009. 水分胁迫对白三叶光合特性的影响[J]. 上海应用技术学院学报(自然科学版),9(2):106-120. [Han J Q. 2009. Effect of water stress on photosynthesis of Trifolium repens[J]. Journal of Shanghai Institute of Technology(Natural Science Edition),9(2):106-120.]
何亚南. 2017. 苹果不同砧穗组合抗旱生理特性的比较研究[D]. 杨凌:西北农林科技大学. [He Y N. 2017. Compa-rative study of the physiological characteristics on drought resistance in different applerrootstock-scion combinations[D]. Yangling:Northwest A & F University.]
贺康宁,田阳,史常青,宋吉红,张光灿. 2003. 黄土半干旱区集水造林条件下林木生長适宜的土壤水分环境[J]. 林业科学,39(1):10-16. [He K N,Tian Y,Shi C Q,Song J H,Zhang G C. 2003. The suitable soil moisture condition for forest growthin catchment afforestation in semia-rid regionon loess plateaus[J]. Scientia Silvae Sincae,39(1):10-16.]
吉小敏,宁虎森,梁继业,高明月,李磊. 2012. 不同水分条件下梭梭和多花柽柳苗期光合特性及抗旱性比较[J]. 中国沙漠,32(6):57-62. [Ji X M,Ning H S,Liang J Y,Gao M Y,Li L. 2012. Comparison of drought resistance and photosynthetic characteristics of Haloxylon ammodendron and Tamarix hohenackeri at seedling stage under different moisture conditions[J]. Journal of Desert Research,32(6):57-62.]
简营. 2016. 遮阴和胁迫处理对莲雾相关生理指标的影响[D]. 广州:华南农业大学. [Jian Y. 2016. Effects of sha-ding,drought stress and water logging on related physiological index of wax-apple[D]. Guangzhou:South China Agricultural University.]
姜英淑,陈书明,王秋玉,卢宝明. 2009. 干旱胁迫对2个欧李种源生理特征的影响[J]. 林业科学,45(6):6-10.[Jiang Y S,Chen S M,Wang Q Y,Lu B M. 2009. Effects of the drought stress on physiological characteristics of two Cerasus humilis provenances[J]. Scientia Silvae Sincae,45(6):6-10.]
李方安,孙歆,张利霞,易小艳. 2013. 干旱胁迫对大豆幼苗期抗逆性的影响[J]. 西南农业学报,26(4):1406-1409. [Li F A,Sun X,Zhang L X,Yi X Y. 2013. Effects of drought on stress-resistance characteristics of soybean seedlings[J]. Southwest China Journal of Agricultural Scien-ces,26(4):1406-1409.]
李芳晓,秦栋,李曙雷,战歌,霍俊伟,王欢欢,赵毅,韩伟,步鹏志. 2014. 黑穗醋栗AsA含量及其代谢酶活性差异分析[J]. 南方农业学报,45(7):1237-1241. [Li F X,Qin D,Li S L,Zhan G,Huo J W,Wang H H,Zhao Y,Han W,Bu P Z. 2014. Ascorbic acid content and difference of related enzyme activities in fruit of blackcurrant[J]. Journal of Southern Agriculture,45(7):1237-1241.]
刘家尧. 2010. 植物生理学实验教程[M]. 北京:高等教育出版社. [Liu J Y. 2010. Plant physiology experiment course[M]. Beijing:Higher Education Press.]
李叶峰,王宁,王采晨,盛亚丹,陆小平. 2011. 盆栽勋章菊对自然干旱胁迫的生理响应研究[J]. 北方园艺,(2):85-88. [Li Y F,Wang N,Wang C C,Sheng Y D,Lu X P. 2011. Study on physiological responses of Gazania rigens L. under nature drought stress[J]. Northern Horticulture,(2):85-88.]
热依扎·朱木斯别克. 2013. 黑加仑抗旱生理特性及果实品质研究[D]. 乌鲁木齐:新疆农业大学. [Reyiza Zhumusibieke. 2013. Study on physological chracteristics of drought resistance and fruit quality of blackcurrant[D]. Urumqi:Xinjiang Agricultural University.]
时连辉,牟志关,姚健. 2005. 不同桑树品种在土壤水分胁迫下膜伤害和保护酶活性变化[J]. 蚕业科学,31(1):13-17. [Shi L H,Mu Z G,Yao J. 2005. Cell membrance damage and change of protective enzymes activity in different mulberry varieties under soil water stress[J]. Acta Sericologica Sinica,31(1):13-17.]
田浩. 2014. 3个枣品种抗旱性生理的研究[D]. 太原:山西农业大学. [Tian H. 2014. Studies on drought resistance phy-
siology ofthree varieties of Jujube[D]. Taiyuan:Shanxi
Agricultural University.]
王瑞,于洪俠,祁永会. 2016. 黑穗醋栗开发利用现状及发展前景[J]. 现代农业研究,(1):30. [Wang R,Yu H X,Qi Y H. 2016. Development and utilization of black currant and its development prospects[J]. Modern Agricultural Research,(1):30.]
魏孝荣,郝明德,张春霞,王旭刚. 2005. 土壤干旱条件下外源锌、锰对夏玉米光合特性的影响[J]. 作物学报,31(8):1101-1104. [Wei X R,Hao M D,Zhang C X,Wang X G. 2005. Effects of zinc and manganese fertilizers on maize photosynthetic performance undersoil drought condition[J]. Acta Agronomica Sinica,31(8):1101-1104.]
徐建欣,杨洁,刘实忠,王云月. 2014. 干旱胁迫对云南陆稻幼苗生理特性的影响[J]. 中国农学通报,30(27):145-152. [Xu J X,Yang J,Liu S Z,Wang Y Y. 2014. Effect of drought stress on physiological characteristics of upland riceseedling in Yunnan Province[J]. Chinese Agricultural Science Bulletin,30(27):145-152.]
许翩翩,王建柱. 2018. 三种常见边坡植物对模拟干旱环境抗旱性能的研究[J]. 草业学报,27(2):36-47. [Xu P P,Wang J Z. 2018. Drought resistance of three common slope pants determined in a simulated drought experiment[J]. Acta Prataculturae Sinica,27(2):36-47.]
杨玉海,李卫红,陈亚宁,朱成刚,马建新. 2013. 极端干旱区自然环境下胡杨幼株对土壤渐进式干旱的生理响应[J]. 林业科学,49(11):171-176. [Yang Y H,Li W H,Chen Y N ,Zhu C G,Ma J X. 2013. Physiological response of populus euphratica seedlings to progressive soil drought under natural environment in an extreme arid area[J]. Scientia Silvae Sinicae,49(11):171-176.]
邹琦. 2004. 植物生理学实验指导[M]. 北京:中国农业出版社. [Zou Q. 2004. Plant physiology experiment guide[M]. Beijing: China Agricultural Press.]
张壮飞. 2012. 高温胁迫对黑穗醋栗(Ribes nigrum L.)幼生理生物特性影响[D]. 哈尔滨:东北农业大学. [Zhang Z F. 2012. The effect of high temperature stress on photosyn
thesis and hysiological and biochemical characteristics of
black currant seedlings(Ribes nigrum L.)[D]. Harbin:
Northeast Agricultural University.]
郑海燕. 2009. 野生樱桃李抗寒生理特性研究[D]. 乌鲁木齐:新疆农业大学. [Zheng H Y. 2009. The research of physiological characteristics of drought resistance on wild cherry plum(Prunus divaricata Ldb)[D]. Urumqi:Xinjiang Agricultural University.]
郑炳松. 2006. 现代植物生理生化技术研究[M]. 北京:气象出版社. [Zheng B S. 2006. Research on physiology and
biochemistry of modern plants[M]. Beijing:Meteorologi-
cal Publishing House.]
周憶堂,马红群,梁丽娇,洪鸿,胡丽涛,孙敏,吴能表. 2008. 不同光照条件下长春花的光合作用和叶绿素荧光动力学特征[J]. 中国农业科学,41(11):3589-3595. [Zhou Y T,Ma H Q,Liang L J,Hong H,Hu L T,Sun M,Wu N B. 2008. Photosynthetic characteristics and chlorophyll fluorescence in leaves of Catharanthus roseus grown under different light intensities[J]. Scientia Agricultura Sinica,41(11):3589-3595.]
Ao H,Zhang Y. 2007. Effects of water stress on photosynthe-tic characteristics of spruce[J]. Bulletin of Botanical Research,27(4):445-451.
Chaves M M,Maroco J P,Pereira J S. 2003. Understanding plant responses to drought-from genes to the whole plant[J]. Functional Plant Biology,30:239-264.
Gill S S,Tuteja N. 2010. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants[J]. Plant Physiology and Biochemistry,48(12):909-930.
Jerry L,Thomas J,Prueger H. 2001. Managing soils to achieve greater water use efficiency[J]. Agronomy Journal,93(2):271-290.
Yamada M,Morishita H,Urano K,Noriko S,Kazuko Y S,Kazuo S,Yoshu Y. 2005. Effects of free proline accumulation in petunias underdrought stress[J]. Journal of Experimental Botany,56(417):1975-1981.
(责任编辑 思利华)
猜你喜欢
天津农林科技(2023年3期)2023-06-27 00:59:44
大自然探索(2021年5期)2021-08-20 05:43:23
中国果业信息(2019年10期)2019-01-05 07:42:30
中国果业信息(2018年1期)2018-01-17 23:36:16
江苏农业科学(2016年8期)2017-02-15 19:38:57
安徽农学通报(2016年24期)2017-01-12 21:06:57
山东农业科学(2016年11期)2016-12-17 21:16:16
现代园艺(2016年2期)2016-03-15 16:05:02
江苏农业科学(2015年8期)2015-09-10 15:56:05
湖北农业科学(2015年15期)2015-09-09 22:21:54
