
镉对两个同核异质红麻杂交种种子萌发及幼苗生长的影响
2019-09-10 07:22贾瑞星丁鑫超汤丹峰韦范常蒙蒙李增强梁志辰陈鹏
南方农业学报 2019年8期
贾瑞星 丁鑫超 汤丹峰 韦范 常蒙蒙 李增强 梁志辰 陈鹏



摘要:【目的】研究重金属镉对红麻杂交种种子萌发、幼苗生长及生理特性的影响,为揭示红麻对重金属镉胁迫的生理响应机制打下基础。【方法】以同核异质的两个红麻杂交品种C1P3A/992(简称C1)和C2P3A/992(简称C2)为材料,采用Cd2+浓度分别为0(对照)、15和30 mg/kg的CdCl2溶液处理红麻种子,测定种子的发芽率、发芽势和发芽指数;同时采用Cd2+浓度分别为0(对照)、10、20、30和50 mg/kg的CdCl2溶液处理两个红麻杂交品种幼苗,8 d后测定幼苗的株高、茎粗、根长及其鲜重和干重的相对抑制率,以及叶片抗氧化酶活性和丙二醛(MDA)含量,综合分析两个品种间的差异。【结果】不同浓度的Cd2+胁迫对两个红麻杂交种种子的萌发均有促进作用,在同一Cd2+浓度下,C2品种的发芽率、发芽势和发芽指数均显著高于C1品种(P<0.05,下同)。随着Cd2+浓度的升高,两个红麻杂交品种幼苗的株高、茎粗、根长及其鲜干重均受到明显抑制,在Cd2+浓度为50 mg/kg时抑制程度最大;在同一Cd2+浓度下,C2品种幼苗各部分鲜干重相对抑制率均显著低于C1品种。两个杂交种幼苗的叶片过氧化物酶(POD)、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活性及MDA含量总体上呈先升高后降低的变化趋势,3种酶的活性均在10 mg/kg Cd2+浓度处理时达最大值,MDA含量在30 mg/kg Cd2+浓度处理时达最大值,在10~50 mg/kg Cd2+浓度处理下,C2品种幼苗叶片的SOD和CAT活性均显著高于C1品种,而MDA含量显著低于C1品种。【结论】两个红麻杂交品种对Cd2+的敏感度不同。在0~50 mg/kg范围内,C2品种在种子萌发、幼苗生长及生理指标方面均表现出更好的适应性,对Cd2+的耐性优于C1品种,可作为母本用于配制抗镉胁迫的三系杂交种。
关键词: 红麻;重金属胁迫;镉;种子萌发;农艺性状;生理指标
中图分类号: S536.5 文獻标志码: A 文章编号:2095-1191(2019)08-1688-07
Effects of seed germination and seedling growth of two homonuclear-heterocytoplasmic kenaf hybrid
cultivars under cadmium stress
JIA Rui-xing1,2, DING Xin-chao1,2, TANG Dan-feng1,2, WEI Fan1,2,
CHANG Meng-meng1,2, LI Zeng-qiang1,2, LIANG Zhi-chen1,2, CHEN Peng1,2*
(1College of Agriculture,Guangxi University, Nanning 530004, China; 2Guangxi Colleges and Universities Key Laboratory of Plant Genetics and Breeding, Nanning 530004, China)
Abstract:【Objective】The effects of cadmium on seed germination, seedling growth and physiological characteristics of kenaf hybrids were studied. It laid a foundation for revealing the physiological response mechanism of kenaf to cadmium stress. 【Method】Two homonuclear-heterocytoplasmic kenaf hybrids,C1P3A/992(C1)and C2P3A/992(C2),were treated with CdCl2 solution with Cd2+ concentrations being 0(control),15 and 30 mg/kg,respectively. The germination percentage,germination potential and germination index of the seeds were determined. At the same time, two hybrid seedlings were treated with CdCl2 solution with Cd2+ concentrations being 0(control), 10, 20, 30 and 50 mg/kg, respectively,plant height,stem diameter,root length of kenaf seedlings,the relative inhibition rate of fresh and dry weights and antioxidant enzyme activity and malondialdehyde(MDA) content in leaves were investigated, and the difference between the two cultivars was analyzed. 【Result】Different concentrations of Cd2+ stress promoted the germination of two kenaf hybrid seeds. At the same Cd2+ concentration, the germination rate, germination potential and germination index of C2 were si-gnificantly higher than those of C1(P<0.05, the same below). With the concentration increasing of Cd2+ treatment,the plant height,stem diameter,root length,fresh wight and dry weight of the two kenaf hybrid cultivars seedlings were inhibi-ted,and the degree of inhibition was maximized when treated with 50 mg/kg Cd2+. Under the same concentration of Cd2+,the relative inhibition rate of fresh and dry weight of C2 seedlings were significantly lower than C1. The activities of pero-xidase(POD), superoxide dismutase(SOD), catalase(CAT) and MDA content in leaves of two hybrids seedlings increased first and then decreased. The activities of POD,SOD and CAT reached the maximum at the treatment of 10 mg/kg Cd2+ concentration,and the MDA content reached the maximum at the treatment of 30 mg/kg Cd2+ concentration. The SOD and CAT activities in leaves of C2 seedlings were significantly higher than those of C1 under 10-50 mg/kg Cd2+ concentration treatment, while MDA content was significantly lower than that of C1 seedlings. 【Conclusion】The sensitivity of two kenaf hybrids to Cd2+ is different. The C2 has better adaptability in seed germination,agronomic traits and physiological indexes,and has better tolerance to Cd2+ than C1 variety in the range of 0-50 mg/kg of the Cd2+ stress. C2 variety can be used as maternal parents to prepare three-line hybrid varieties resistant to cadmium stress.
Key words: kenaf; heavy metal stress; cadmium; seed germination; agronomic traits; physiological indexes
0 引言
【研究意义】重金属污染效应具有长期性、隐蔽性及不可逆性等,已成为环境污染治理的热点和难点(宋玉婷和雷泞菲,2018)。在重金属污染的修复方法中,植物修复技术因具有成本较低、无二次污染及具有一定经济价值等优点而得到国内外学者的广泛关注,筛选出对重金属耐性强、富集能力高的植物是植物修复技术应用的关键。镉是一种半衰期长、毒性高、迁移性强的重金属,微量即可严重阻碍农作物生长,是发展中国家环境污染最严重的污染物之一,其对植物的毒性极强,在重金属潜在毒性有害元素分类中位居第Ⅰ类第2位(刘红梅等,2018)。红麻(Hibiscus cannabinus L.)是锦葵科木槿属作物,生长周期较短,生命力极强(王国庆等,2006),具有较高的经济价值(苏旭中等,2018),同时,红麻是一种高产的纤维作物,具有生长速度快及抗逆境能力强等特点(廖英明等,2009)。由于红麻生物量大、对重金属有较强的耐性,在土壤重金属修复方面具有很大潜力(李文略等,2018),利用红麻复垦是改良大田土壤重金属含量的有效措施(杨煜曦等,2013)。因此,研究镉胁迫对红麻杂交种种子萌发及幼苗生长的影响,对缓解镉污染對红麻的毒害作用及探讨不同细胞质背景的红麻杂交种对重金属镉的抗性差异具有重要意义。【前人研究进展】重金属可在植株体内累积,超过植株承受范围后,植物生长会受到抑制,表现为叶片发黄、卷曲甚至脱落,植株矮小,根尖变黑发绿等。张嘉桐等(2018)发现镉对植物细胞的膜透性、光合和呼吸代谢过程、酶作用机制及遗传效应等生命过程均有毒害作用。关于重金属镉对麻类生长发育的影响,邓勇(2016)研究表明,不同品种红麻幼叶受镉毒害的程度随镉浓度的增加及胁迫时间的延长而加重,镉累积效应抑制红麻的生长,导致红麻根系活力降低、根膜通透性增加及叶绿素含量降低,幼叶抗氧化酶活性的差异可能是不同品种耐性差异的关键因素;李文略等(2018)发现镉对红麻生长影响明显,但红麻对镉有较强的耐性;姚运法等(2018)研究了不同黄、红麻品种在重金属镉胁迫下的生物产量及不同部位对镉的富集能力,发现红麻对镉的富集能力高于黄麻。杂种优势是质核互作的结果,意味着不同细胞质背景会对杂种优势产生不同影响。邹瑞昌等(2012)对芥菜细胞质雄性不育系的胞质效应和杂种优势进行研究,结果表明不同的细胞质背景对于杂种优势有重要影响。孙丽芳等(2016)研究了3个不同胞质玉米不育系杂交种的杂种优势利用情况,发现不同胞质背景对杂种优势的影响较大。在逆境胁迫环境下,杂交种红麻具有更强的抗逆性,其在农艺性状、生物量和酶活性等方面均会表现出中亲优势和超亲优势。张加强等(2011)研究了盐胁迫对红麻杂交种幼苗生长的影响,发现红麻杂交种F1代对于逆境胁迫表现出较强的中亲优势和超亲优势,其株高、茎粗、根长等指标均优于亲本。【本研究切入点】近年来与红麻抗逆性有关的研究主要体现在杂交种优势利用方面,而关于重金属镉对不同细胞质背景红麻杂交种的毒害程度和机理尚不明确,也未见不同细胞质背景红麻杂交种对重金属镉抗逆性差异的研究报道。【拟解决的关键问题】以红麻同核异质不育系杂交种C1P3A/992(简称C1)和C2P3A/992(简称C2)为试验材料,在水培条件下研究重金属镉对红麻杂交种种子萌发、幼苗生长及生理特性的影响,为揭示红麻对重金属镉胁迫的生理响应机制打下基础。
1 材料与方法
1. 1 试验材料
C1和C2分别为野生型红麻品种UG93雄性不育突变体的选育品种C1P3A、C2P3A与恢复系992的杂交F1代,由广西大学周瑞阳教授提供。镉胁迫处理使用CdCl2(分析纯,购自天津市大茂化学试剂厂),先用蒸馏水配制成10 mg/mL的母液,使用时稀释成所需浓度。霍格兰氏(Hoagland)营养液购自山东拓普生物工程有限公司。
1. 2 试验方法
发芽试验:挑选籽粒饱满的红麻种子,放入蒸馏水中浸泡20 min,再用3% H2O2浸泡10 min,然后用蒸馏水冲洗3次以保证去除残留的H2O2;将消毒完的种子平铺在已灭菌的培养皿中(覆盖一层灭菌滤纸),加入15 mL处理液。共设3个镉胁迫处理,Cd2+浓度分别为0(蒸馏水,对照)、15和30 mg/kg,每处理3次重复,每重复100粒,于光照培养箱中进行胁迫处理。培养条件为光照16 h、黑夜8 h;昼夜温度为白天30 ℃、黑暗26 ℃。每天更换一次处理液,培养到第8 d为止。
苗期试验:种子先用蒸馏水浸泡20 min,再用3% H2O2浸泡10 min,然后用蒸馏水冲洗3次,以滤纸吸干种子表面的水分。将消毒完的种子播种于装有育秧基质的育秧盘中,待幼苗长至7~9片真叶时(约20 d)转移至水培。水培试验中,每个红麻品种分别挑取长势良好且一致的幼苗30株,分成3组,每个容器加1/2 Hoagland营养液1 L进行预培养,培养条件为光照16 h、黑夜8 h;昼夜温度为白天30 ℃、黑暗26 ℃。2 d后开始用含CdCl2的1/2 Hoagland营养液进行胁迫培养,共设5个镉胁迫处理,每处理3次重复,Cd2+浓度分别为0(蒸馏水,对照)、10、20、30和50 mg/kg,培养条件同上,此后每2 d更换一次培养液,8 d后停止胁迫。
1. 3 测定项目及方法
发芽试验:发芽第2 d开始统计发芽率,第3 d开始计算发芽势和发芽指数,发芽至第8 d为止。发芽势(%)=发芽高峰期发芽种子数/供试种子数×100,发芽指数=Σ(逐日发芽种子数/对应发芽日数)。
苗期试验:胁迫8 d后,测量红麻幼苗株高、茎粗、根长及根茎叶各部分鲜重,再将各部分鲜样置于烘箱中105 ℃杀青15 min,随后立即降温至80 ℃烘至恒重并记录各部分干重,计算鲜重和干重相对抑制率,相对抑制率(%)=处理组相应值/对照组相应值×100。同时,选取两个红麻品种苗期真叶,参照李合生(2000)的方法测定抗氧化酶活性及丙二醛(MDA)含量,其中过氧化物酶(POD)活性采用愈创木酚法测定,超氧化物歧化酶(SOD)活性采用氮蓝四唑法测定,过氧化氢酶(CAT)活性采用紫外吸收法测定,MDA含量采用硫代巴比妥酸法测定。
1. 4 统计分析
使用SPSS 18.0对试验数据进行方差分析,采用Excel 2013进行制图。
2 结果与分析
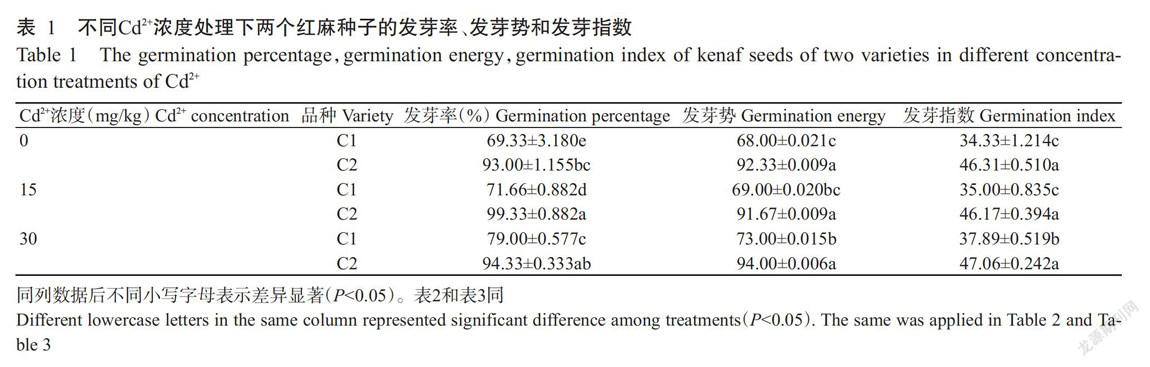
2. 1 红麻发芽试验结果
由表1可知,与对照相比,随着Cd2+浓度的升高,C1品种的发芽率、发芽势和发芽指数均逐渐升高,在Cd2+浓度为30 mg/kg时达最大值;C2品种的发芽率先升高后下降,在Cd2+浓度为15 mg/kg时达最大值,发芽势和发芽指数则先降低后升高,在Cd2+浓度为30 mg/kg时达最大值。在同一Cd2+浓度下,C2品种的发芽率、发芽势和发芽指数均显著高于C1品种(P<0.05,下同)。说明C2品种在不同Cd2+浓度胁迫下的发芽情况均优于C1品种。
2. 2 镉胁迫对不同胞质红麻幼苗农艺性状的影响
2. 2. 1 不同处理下红麻幼苗的形态观察 通过观察,发现两个红麻品种在不同Cd2+浓度胁迫下的幼苗形态相似,图1为C2品种的幼苗生长情况。由图1可看出,与对照相比,在不同Cd2+浓度处理下红麻幼苗生长均受到明显抑制,且随着Cd2+浓度升高,抑制程度逐渐加重,红麻幼苗出现植株生长缓慢、主根变短、须根量减少、叶片卷曲、失绿、脱落及叶面积减小的现象。尤其从图1-a和图1-c可看出,随着Cd2+浓度的升高,红麻幼苗和根生長受到的抑制程度逐渐加重,浓度越高叶片脱落越严重;与对照相比,在Cd2+浓度为10 mg/kg时,对根的抑制作用主要体现在抑制根系伸长,在20、30和50 mg/kg时,则主要表现为须根量减少。
2. 2. 2 不同处理对红麻幼苗株高、茎粗和根长的影响 由表2可知,在苗期胁迫试验中,与对照相比,两个红麻品种的株高、茎粗和根长均随着Cd2+浓度的升高显著降低,在Cd2+浓度为50 mg/kg时达最低值;相同浓度处理下,两个品种间无显著差异(P>0.05,下同),但同一品种在不同浓度处理下,其株高、茎粗和根长差异明显。
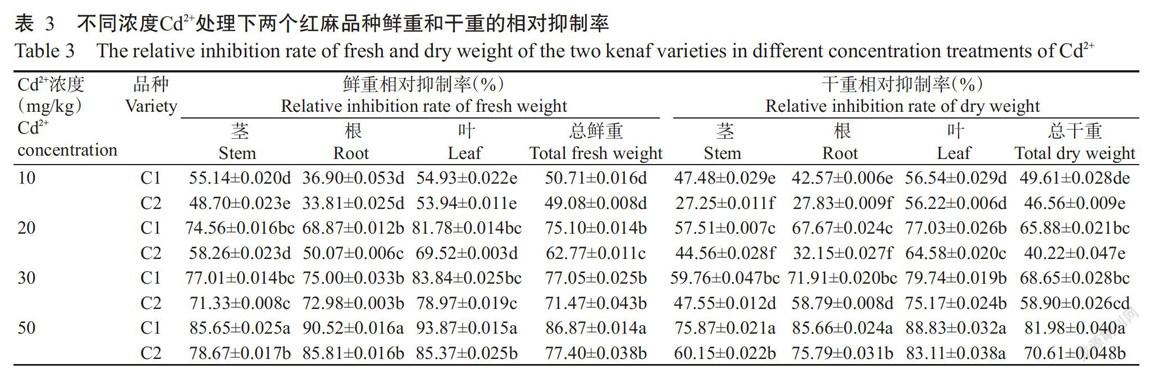
2. 2. 3 不同处理下红麻幼苗鲜、干重的相对抑制率 由表3可知,随着Cd2+浓度的升高,两个红麻品种的茎、根、叶和总鲜重的相对抑制率均显著升高,Cd2+浓度为50 mg/kg时,两个品种各部分鲜重的相对抑制率均达最大值,C1品种的茎、叶、根和总鲜重的相对抑制率均显著高于C2品种。此外,镉对红麻幼苗的生物量有明显影响,随着Cd2+浓度的升高,两个品种的茎、根、叶和总干重的相对抑制率均明显升高,Cd2+浓度为50 mg/kg时,两个品种各部分干重的相对抑制率达最大值;同一浓度处理下,C1品种的茎、叶、根和总干重的相对抑制率均高于C2品种。综上所述,在不同浓度Cd2+胁迫下,C2品种各部分鲜重和干重受到的抑制程度均低于C1品种。
2. 3 镉胁迫对不同胞质红麻幼苗叶片生理特性的影响
2. 3. 1 对POD活性的影响 POD的主要作用是清除机体内的自由基,参与多种生化反应(林杰等,2009)。由图2-A可看出,随着Cd2+浓度的升高,红麻幼苗叶片POD活性总体上呈先升高后降低的变化趋势,均在Cd2+浓度为10 mg/kg时达最高值,且显著高于其他处理组,在Cd2+浓度为30 mg/kg时达最低值。在Cd2+浓度为10和30 mg/kg时,C1品种的叶片POD活性均低于C2品种。
2. 3. 2 对SOD活性的影响 当植物受到外界环境迫害时会产生超氧负离子,SOD可使超氧负离子发生歧化反应,从而降低其毒害作用(Sreenivasulu et al.,2000)。由图2-B可看出,随着Cd2+浓度的升高,红麻幼苗叶片SOD活性总体上也呈先升高后降低的变化趋势,在Cd2+浓度为10 mg/kg时达最高值,随后逐渐缓慢降低。除对照外,同一Cd2+浓度处理下C1品种的SOD活性均低于C2品种。
2. 3. 3 对CAT活性的影响 CAT是植物防御系统的重要组成酶之一,植物在受到逆境迫害时,会通过提高CAT活性来防御逆境胁迫(张建新等,2018)。由图2-C可看出,随着Cd2+浓度的升高,CAT活性急剧升高后下降,两个品种在Cd2+浓度为10 mg/kg时CAT活性最高,除对照外,同一Cd2+浓度处理下C1品种的CAT活性均显著低于C2品种。
2. 3. 4 对MDA含量的影响 MDA是细胞膜脂质发生过氧化反应的重要产物,是反映植物膜系统损害程度的重要指标(曾小飚等,2015)。由图2-D可看出,与对照相比,Cd2+浓度为10 mg/kg时,红麻幼苗叶片的MDA含量显著降低,之后随着Cd2+浓度的升高,叶片MDA含量先急速上升,在Cd2+浓度为30 mg/kg时达最大值,当Cd2+浓度增大至50 mg/kg时MDA含量有所降低。Cd2+浓度为20、30和50 mg/kg时,C1品种的MDA含量均显著高于C2品种。
3 讨论
在所有的重金属污染元素中,镉是关注度最高、污染最严重的重金属,能影响植物种子萌发,造成发芽率、发芽势和发芽指数等指标发生改变。赵淑玲等(2018)研究发现,随着Cd2+胁迫浓度的升高,花椰菜种子的发芽率呈先升高后下降的变化趋势;李兰平等(2012)研究了镉对不育系P3A、恢复系992及其杂交F1代红麻种子萌发的影响,发现随着处理浓度的增加,不同红麻种子的发芽势、发芽率和发芽指数总体呈下降趋势,但部分品种的发芽率出现一定波动。本研究发现,随着Cd2+浓度的升高,C1品种的发芽率、发芽势和发芽指数逐渐升高,C2品种的发芽率先升高后降低,发芽势和发芽指数先降低后升高,与李兰平等(2012)、赵淑玲等(2018)的研究结果有相似之处,究其原因可能是低浓度的Cd2+有利于增强红麻幼苗体内的水解酶活性,进而提高种子活力。但高浓度处理下,C1品种的发芽率、发芽势和发芽指数并未出现降低,可能是杂种优势有助于提高红麻种子对镉的耐受性;同时,在相同Cd2+浓度条件下,C2品种的发芽率、发芽势和发芽指数均优于C1品种,表明不同红麻品种种子萌发对重金属镉的耐受性存在差异。
当环境中的Cd2+达一定浓度时会影响植物的生长,造成植株生长速度缓慢、植株矮化,根系生长受到抑制,严重时会造成植物死亡(闫寒等,2017)。陈顺钰等(2018)研究表明,镉胁迫处理下,枫香幼苗的根长、株高和生物量等均受到抑制,抑制程度与镉胁迫浓度呈正相关。本研究结果表明,Cd2+会抑制两个红麻杂交种的株高、茎粗、根长及各部分鲜重、干重,且随Cd2+浓度的增加,抑制程度逐渐加重。与陈顺钰等(2018)的研究结果相似,当植物受到镉胁迫时,首先会抑制植物根系生长,导致植物对水分和营养物质的吸收速率下降,继而影响地上部分的生长;其次,在低浓度Cd2+处理下,两个红麻品种的叶片出现变黄失绿等现象,其原因可能是镉破坏了植物的光合系统,导致叶绿素含量下降,但在高浓度时叶片与对照差异不明显,可能是高浓度处理下大部分的Cd2+被红麻根系吸收固定,从而减轻其对叶片的破坏作用。本研究中,在不同Cd2+浓度处理下,两个同核异质红麻杂交种在各部分鲜重、干重相对抑制率方面均表现为C2品种低于C1品种,表明C2品种的幼苗能更好地适应镉胁迫环境。
SOD、POD和CAT是植物抗氧化酶系统的重要成员,对植物清除自身过氧化物质起重要作用,其活性变化与植物体内氧化胁迫有直接关系(李丽锋等,2014)。本研究发现,在不同浓度的Cd2+胁迫处理下,3种酶活性整体上均呈先上升后下降的变化趋势,两个红麻杂交种的3种酶活性均在Cd2+浓度为10 mg/kg时达最高值,随后活性有所下降,与李正文等(2013)、简敏菲等(2017)的研究结果基本一致,即在低浓度胁迫时,植株体内会产生大量的POD、SOD和CAT以消除逆境下体内产生的有毒物质,对植物起保护作用,但Cd2+浓度达一定范围后,就会超过植物的耐受限度,导致活性氧自由基含量增多,此时活性氧物质通常会对抗氧化酶产生氧化损伤,从而导致POD、SOD和CAT活性降低。通过比较同一处理下两个不同杂交种幼苗的酶活性发现,C2品种的抗氧化酶活性更高,相对于C1品种能更好地适应胁迫环境。
植物受到重金属胁迫时会产生大量的过氧化物(ROS),而ROS过量会导致细胞质膜系统损伤,通透性发生改变,使膜脂过氧化产生MDA(周妍英和罗正明,2018),MDA含量越高表示膜脂过氧化程度越高,细胞膜受到的损伤越严重(余沛东等,2019)。本研究发现,在10~50 mg/kg范围内,随着Cd2+浓度的升高,两个红麻幼苗叶片的MDA含量呈先升高后降低的变化趋势,与孙刚(2013)、刘丽欣等(2017)的研究结果基本一致,即在重金属胁迫下,细胞膜的受损程度随胁迫浓度的升高而升高,细胞壁遭受破坏,内溶物外泄,MDA等物质在细胞内积累,但浓度过高(50 mg/kg)时,MDA含量出现下降,此时红麻幼苗的生理活动受到严重影响,细胞结构遭到严重破坏,开始出现细胞死亡。同时,在同一中、高Cd2+浓度(20~50 mg/kg)处理下,发现C1品种的MDA含量均显著高于C2,说明C1品种在逆境下更容易发生膜质过氧化,对镉胁迫的敏感性更高。
4 结论
两个红麻杂交品种对Cd2+的敏感度不同。在0~50 mg/kg范围内,C2品种在种子萌发、幼苗生长及生理指标方面均表现出更好的适应性,对Cd2+的耐性优于C1品种,可作为母本用于配制抗镉胁迫的三系杂交种。
参考文献:
陈顺钰,韩航,薛凌云,张韵,侯晓龙,蔡丽平,周垂帆. 2018. Pb、Cd和酸胁迫对枫香种子萌发、幼苗生长及体内抗氧化酶活性的影响[J]. 农业环境科学学报,37(4):647-655. [Chen S Y,Han H,Xue L Y,Zhang Y,Hou X L,Cai L P,Zhou C F. 2018. Effects of Pb,Cd and acid stress on seed germination,seedling growth and antioxidant enzyme activities of Liquidambar formosana[J]. Journal of Agro-Environment Science,37(4):647-655.]
邓勇. 2016. 红麻耐镉生理响应相关机制的研究[D]. 长沙:湖南农业大学. [Deng Y. 2016. The study of physiological response mechanism in kenaf under cadmium stress conditions[D]. Changsha:Hunan Agricultural University.]
简敏菲,史雅甜,陈涛,何琦瑜,崔妍,杜恺翔. 2017. 重金属镉、铅胁迫下湿地植物丁香蓼的生理生化特征研究[J]. 江西师范大学学报(自然科学版),41(1):93-98. [Jian M F,Shi Y T,Chen T,He Q Y,Cui Y,Du K X. 2017. The physiological and biochemical characteristics of the wetland plant Ludwigia prostrata Roxbunder the heavy metal pollution stress of cadmiumand lead[J]. Journal of Jiangxi Normal University(Natural Science Edition),41(1):93-98.]
李合生. 2000. 植物生理生化實验原理和技术[M]. 北京:高等教育出版社. [Li H S. 2000. Principles and techniques of physiological and biochemical experiments in plants[M]. Beijing:Higher Education Press.]
李兰平,李正文,杨海霞,周琼,周瑞阳,李志刚. 2012. 铅、镉对红麻种子萌发生长的影响[J]. 南方农业学报,43(9):1291-1296. [Li L P,Li Z W,Yang H X,Zhou Q,Zhou R Y,Li Z G. 2012. Effects of lead and cadmium on seed germination and growth of kenaf(Hibiscus cannabinus L.)[J]. Journal of Southern Agriculture,43(9):1291-1296.]
李丽锋,卢佳,苏芳莉. 2014. 镉(Cd)胁迫对芦苇叶片SOD、POD活性影响及动力学分析[J]. 沈阳农业大学学报,45(3):326-330. [Li L F,Lu J,Su F L. 2014. Effects of cadmium stress on SOD,POD activities in reed leaf and their dynamic models[J]. Journal of Shenyang Agricultural University,45(3):326-330.]
李文略,金关荣,骆霞虹,安霞,李苹芳,朱关林,陈常理. 2018. 不同红麻品种的土壤重金属污染修复潜力对比研究[J]. 农业环境科学学报,37(10):2150-2158. [Li W L,Jin G R,Luo X H,An X,Li P F,Zhu G L,Chen C L. 2018. Comparative study on the potential of a kenaf(Hibiscus cannabinus)variety for remediating heavy metal contaminated soils[J]. Journal of Agro-Environment Scien-ce,37(10):2150-2158.]
李正文,李兰平,周琼,李志刚,周瑞阳. 2013. 不同浓度Cd、Pb对红麻抗逆生理特性的影响[J]. 湖北农业科学,52(15):3568-3571. [Li Z W,Li L P,Zhou Q,Li Z G,Zhou R Y. 2013. Effect of different concentrations of Cd and Pb on the stress-resistance physiological characte-ristics of Hibiscus cannabinus L.[J]. Hubei Agricultural Sciences,52(15):3568-3571.]
廖英明,祁建民,张广庆,李点姣,林荔辉,方平平. 2009. 我国红麻雄性不育系选育及对不育机理研究的思考[J]. 中国麻业科学,31(2):125-129. [Liao Y M,Qi J M,Zhang G Q,Li D J,Lin L H,Fang P P. 2009. Discussion on breeding and sterile mechanism of kenaf male sterility in China[J]. Plant Fiber Sciences in China,31(2):125-129.]
林杰,柯金炼,魏道军. 2009. 铅胁迫对香根草叶片膜系统及抗氧化系统的影响[J]. 福建农业学报,24(3):254-257. [Lin J,Ke J L,Wei D J. 2009. Effect of lead stress on membrane and antioxidant systems in Vetiveria zizanioides leaves[J]. Fujian Journal of Agricultural Sciences,24(3):254-257.]
刘红梅,杨凯,肖正午. 2018. 土壤镉污染治理及外源调控研究进展[J]. 作物研究,32(5):449-453. [Liu H M,Yang K,Xiao Z W. 2018. Research progress of soil cadmium pollution control and exogenous regulation[J]. Crop Research,32(5):449-453.]
刘丽欣,迟鑫,王建,刘明飞,徐欢,李铭. 2017. 重金属Cd对粟米生理生化指标的影响[J]. 安徽农学通报,23(7):27-30. [Liu L X,Chi X,Wang J,Liu M F,Xu H,Li M. 2017. Effect of Cd on physiological and biochemical indicators of corn[J]. Anhui Agricultural Science Bulletin,23(7):27-30.]
宋玉婷,雷泞菲. 2018. 我国土壤镉污染的现状及修复措施[J]. 西昌学院学报(自然科学版),32(3):79-83. [Song Y T,Lei N F. 2018. China’s cadmium pollution land status and restoration measures[J]. Journal of Xichang University(Natural Science Edition),32(3):79-83.]
蘇旭中,顾秦榕,赵超,刘新金. 2018. 麻织物服用性能探讨[J]. 上海纺织科技,46(9):14-15. [Su X Z,Gu Q R,Zhao C,Liu X J. 2018. Discussion on the wearing properties of hemp fabric[J]. Shanghai Textile Science & Techno-logy,46(9):14-15.]
孙刚. 2013. 镉、铅胁迫对千屈菜种子萌发、幼苗生长及生理生化的影响[D]. 长春:吉林农业大学. [Sun G. 2013. Effects of cadmium and lead stress on the seed germination and seedling growth in Lythrum salicaria L.[D]. Chang-chun:Jilin Agricultural University.]
孙丽芳,邓杰,王霞,赵伟,杨克军,苗兴芬,徐莹莹,高树仁. 2016. 玉米不同雄性不育胞质杂种优势分析[J]. 玉米科学,24(6):8-11. [Sun L F,Deng J,Wang X,Zhao W,Yang K J,Miao X F,Xu Y Y,Gao S R. 2016. Analysis of different cytoplasmic male sterile heterosis in maize[J]. Journal of Maize Sciences,24(6):8-11.]
王国庆,李敏,骆永明,宋静,赵其国. 2006. 红麻对Cu和Cu-EDDS的吸收和富集[J]. 土壤,38(5):626-631. [Wang G Q,Li M,Luo Y M,Song J,Zhao Q G. 2006. Plant uptake and accumulation of Cu and Cu-EDDS by kenaf (Hibiscus Cannabinus L.)[J]. Soils,38(5):626-631.]
闫寒,宋杭霖,张丽,闫晶,石贤吉,朱诗苗,刘璐,李虎林. 2017. 镉胁迫对烤烟农艺性状及生理生化指标的影响[J]. 作物杂志,(5):156-161. [Yan H,Song H L,Zhang L,Yan J,Shi X J,Zhu S M,Liu L,Li H L. 2017. Effects of cadmium stress on agronomic traits and physiological and biochemical indexes of flue-cured tobacco[J]. Crops,(5):156-161.]
杨煜曦,卢欢亮,战树顺,邓腾灏博,林庆祺,王诗忠,杨秀虹,仇荣亮. 2013. 利用红麻复垦多金属污染酸化土壤[J]. 应用生态学报,24(3):832-838. [Yang Y X,Lu H L,Zhan S S,Deng T H B,Lin Q Q,Wang S Z,Yang X H,Qiu R L. 2013. Using kenaf(Hibiscus cannabinus) to reclaim multi-metal contaminated acidic soil[J]. Chinese Journal of Applied Ecology,24(3):832-838.]
姚运法,赖正锋,林碧珍,练冬梅,洪建基. 2018. 耐镉黄、红麻品种筛选试验[J]. 福建热作科技,43(3):1-4. [Yao Y F,Lai Z F,Lin B Z,Lian D M,Hong J J. 2018. Choice test of cadmium proof vute,kenaf[J]. Fujian Science & Technology of Tropical Crops,43(3):1-4.]
余沛东,陈银萍,闫志强,师小平,丁浚刚,颉海帆. 2019. 硝普钠对铅胁迫娃娃菜种子萌发及其幼苗生理特性的影响[J]. 南方农业学报,50(3):607-614. [Yu P D,Chen Y P,Yan Z Q,Shi X P,Ding J G,Xie H F. 2019. Effects of sodium nitroprusside on seed germination and seedling physiological characteristics of Brassica pekinensis under lead stress[J]. Journal of Southern Agriculture,50(3):607-614.]
曾小飚,李毅,陈冠喜,莫远明. 2015. 重金属镍胁迫对麻风树幼苗生理生化特性的影响[J]. 广东化工,42(21):232-234. [Zeng X B,Li Y,Chen G X,Mo Y M. 2015. Effect of heavy metal nickel stresson physiological and biochemical characteristics in seedling of Jatropha Carcas L.[J]. Guangdong Chemical Industry,42(21):232-234.]
张加强,潘凤英,廖小芳,周瑞阳,杨健,黄龙. 2011. 红麻杂交种幼苗生长对盐胁迫的响应[J]. 华中农业大学学报,30(5):552-557. [Zhang J Q,Pan F Y,Liao X F,Zhou R Y,Yang J,Huang L. 2011. Response of hybrid kenaf seedling to salt stress[J]. Journal of Huazhong Agricultural University,30(5):552-557.]
張嘉桐,关颖慧,司莉青,彭霞薇,孟丙南,周金星. 2018. Pb2+、Cd2+复合胁迫对桑树光合作用的影响[J]. 北京林业大学学报,40(4):16-23. [Zhang J T,Guan Y H,Si L Q,Peng X W,Meng B N,Zhou J X. 2018. Effects of Pb2+ and Cd2+ combined stress on photosynthesis of Morus alba[J]. Journal of Beijing Forestry University,40(4):16-23.]
张建新,张婷婷,龙昌莲,崔乐怡,张航,苟秋,吴玉环,刘鹏. 2018. 重金属镉胁迫下臭牡丹的生理響应及富集研究[J]. 安徽农业大学学报,45(4):703-709. [Zhang J X,Zhang T T,Long C L,Cui L Y,Zhang H,Gou Q,Wu Y H,Liu P. 2018. Physiological response and bioconcentration of Clerodendrum bungei Steud. to cadmium[J]. Journal of Anhui Agricultural University,45(4):703-709.]
赵淑玲,王瀚,王让军,陈强,杨小录,王一峰. 2018. 重金属Cd2+对花椰菜种子的萌发及幼苗生理生化的影响[J]. 种子,37(1):100-102. [Zhao S L,Wang H,Wang R J,Chen Q,Yang X L,Wang Y F. 2018. Effect of Cd2+ stress on seed germination and seedling physiological and biochemical characteristics of cauliflower[J]. Seed,37(1):100-102.]
周妍英,罗正明. 2018. 镉对中华圆田螺抗氧化酶活性和脂质过氧化的影响[J]. 食品工业科技,39(15):43-47. [Zhou Y Y,Luo Z M. 2018. Effects of cadmium on antioxidant enzymes activities and lipid peroxidation in mudsnail(Cipangopaludina cathayensis)[J]. Science and Technology of Food Industry,39(15):43-47.]
邹瑞昌,万正杰,徐跃进,施展,傅廷栋. 2012. 叶用芥菜细胞质雄性不育系0912A的胞质效应和杂种优势分析[J]. 植物科学学报,30(3):261-268. [Zou R C,Wan Z J,Xu Y J,Shi Z,Fu T D. 2012. Cytoplasmic effects and heterosis in cytoplasmic male sterile line of leafy Mustard 0912A[J]. Plant Science Journal,30(3):261-268.]
Sreenivasulu N,Grimm B,Wobus U,Weschke W. 2000. Di-fferential response of antioxidant compounds to salinity stress in salt-tolerant and salt-sensitive seedlings of foxtail millet(Setaria italica)[J]. Physiologia Plantarum,109(4):435-442.
(责任编辑 王 晖)
猜你喜欢
吉林农业·下半月(2017年1期)2017-02-16
安徽农学通报(2017年1期)2017-02-15
物联网技术(2016年12期)2017-01-21
天津农业科学(2016年12期)2017-01-11
现代农业科技(2016年20期)2016-12-20
中国中药杂志(2016年20期)2016-11-19
