
微生物硫酸盐同化的调控及其在提高重金属抗性中的研究进展
2019-09-10 09:38王小雨
四川环境 2019年4期
毛 娟,王小雨
(东北师范大学环境学院,长春 130117)
1 引 言
硫是生物体生存的必需营养元素之一[1],生物体内的硫主要以还原态的形式在有机硫醇物质中存在[2]。硫酸盐同化是指硫酸盐在生物作用下合成为有机硫化物的过程,即首先将硫酸盐中的硫转变为还原态的硫,并将其加入到生物体的含硫代谢物,如半胱氨酸和胱氨酸等有机硫化物中,构成生物的细胞组分。大多数微生物和植物均可参与这一过程。硫酸盐同化途径的产物半胱氨酸是无机硫元素最终转变为有机硫并参与生物体代谢的中间载体[3],在生物代谢中发挥重要作用:参与生物体必需氨基酸甲硫氨酸的转变;参与硫胺素、生物素、铁硫簇、辅酶A及辅酶M的合成;是生物体重要抗氧化剂谷胱甘肽、硫蛋白等的合成原料之一[4]。此外,半胱氨酸和甲硫氨酸残基具有修复细菌细胞质和细胞膜氧化损伤的作用[5],半胱氨酸生物合成的抑制会严重干扰病原体抵抗氧化应激[6]。可见,硫酸盐同化在生物体代谢中具有重要作用。
近年来组学的研究发现硫酸盐同化参与了微生物的重金属抗性作用。已经出现了几篇关于微生物硫酸盐同化的综述,如David Mendoza-Co′zatl等人[7]综述了重金属镉胁迫对真核生物的酵母、原生植物和高等植物硫酸盐同化途径的影响及其衍生物谷胱甘肽的解毒作用;Carlo Viti等人[8]指出细菌和真菌中硫代谢相关基因在铬胁迫下显著表达;郑春丽[9]综述了微生物硫酸盐同化途径与重金属抗性的关系。然而,已有的综述多侧重硫酸盐同化途径的过程机理阐述,有关微生物硫酸盐同化途径调控的综述还很少见,尤其是重金属胁迫下微生物硫酸盐同化途径是如何调控的以及硫酸盐同化在微生物重金属抗性中如何发挥作用还未有明确的综述。因而,对微生物硫酸盐同化的综述有助于进一步了解微生物的重金属抗性机制,同时为揭示微生物硫酸盐同化途径及其与重金属抗性的关系提供参考。本文针对微生物体内硫酸盐同化途径的调控及其在重金属抗性中的作用进行总结。
2 微生物硫酸盐同化途径
2.1 微生物硫酸盐同化途径
微生物硫酸盐同化通常是指微生物将硫酸盐经活化、还原、合成半胱氨酸的过程,这一过程亦称作同化型硫酸盐还原。微生物硫酸盐同化途径是微生物体内硫代谢的重要组成部分,是合成生物体代谢所必需的含硫氨基酸的必需前提。硫酸盐同化途径在植物、真菌、细菌和古细菌之间广泛保守,不同界生物间存在细微差别[10]。在真核微生物的酿酒酵母中研究较为透彻,主要是因为酵母菌经常作为生物工程菌生产一些含硫氨基酸或发酵生产葡萄酒等;革兰氏阴性菌中的大肠杆菌中亦有相似的应用。
2.2 微生物硫酸盐同化途径代谢通路
微生物硫酸盐同化途径包括:硫酸盐的转运、活化、还原以及半胱氨酸合成,其同化机理如图1(A)。
(1)转运 环境中的硫酸盐通过主动运输的方式进入生物体细胞内。一般来说,优选的硫源是无机硫酸盐[11]。已知4种细菌转运蛋白可运输硫酸盐和结构类似的硫代硫酸盐:SulT家族;SulP家族;PiT家族的CysP转运蛋白和CysZ转运蛋白[12],如图1(A)。Q在微生物培养过程中硫酸盐是最受欢迎与常用的硫源,例如,硫酸铵、硫酸钠和硫酸镁等硫酸盐[13]。除硫酸盐是细菌的优选硫源外,土壤环境也含有丰富的磺酸盐和硫酸酯[14]。已知细菌中有两类有机硫转运蛋白TauABC和SsuABC,分别负责转运牛磺酸和烷基磺酸[15],如图1(B)。此外,许多微生物也可以吸收非氨基酸的有机硫化合物,如亚砜,砜,磺酸盐和硫酸酯等。这些化合物在经过硫酸酯酶水解后以硫酸盐或亚硫酸盐形式释放,再进入无机硫酸盐同化途径,以满足微生物对硫的需求。
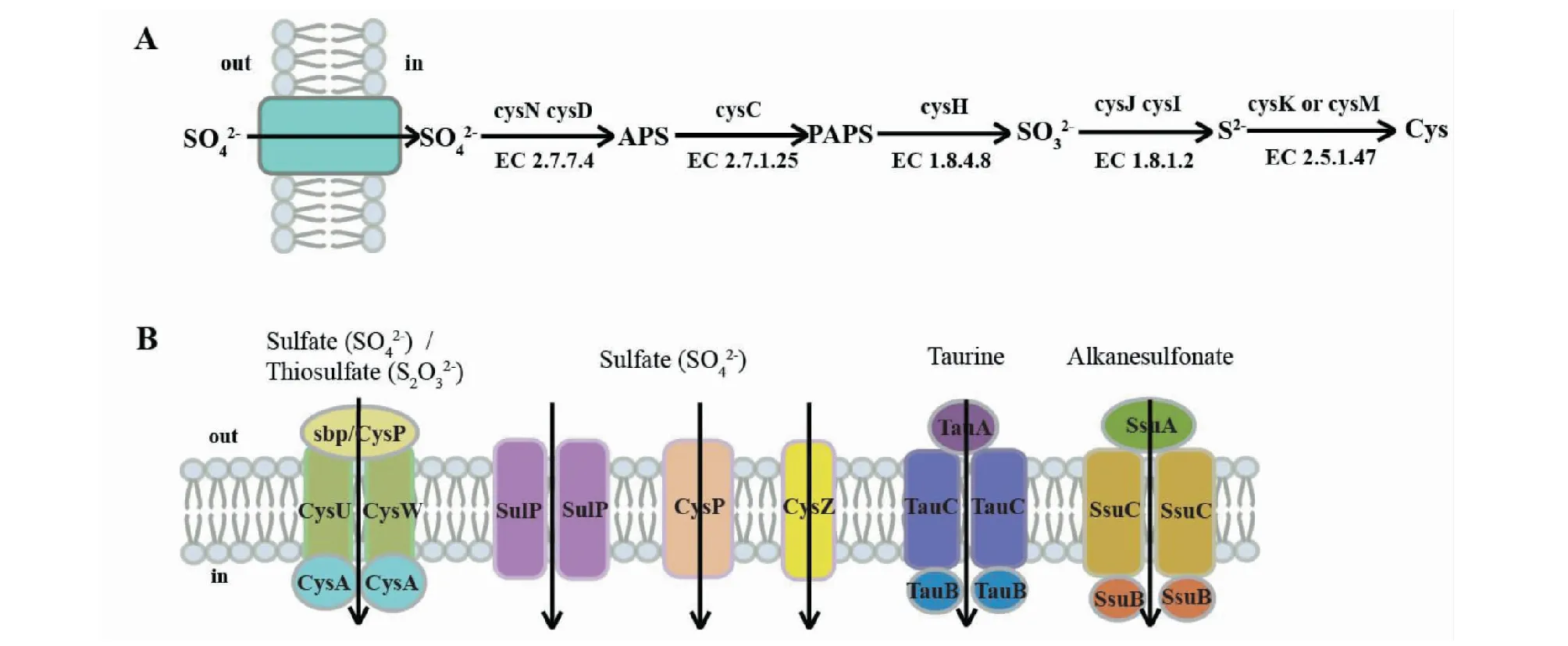
(A: 微生物无机硫酸盐同化途径通路,根据Takeshi Nakatani等人文献[17] 绘制;EC 2.7.7.4,ATP硫酰化酶; EC 2.7.1.25,APS还原酶;EC 1.8.4.8,PAPS还原酶;EC 1.8.1.2,亚硫酸盐还原酶;EC 2.5.1.47,半胱氨酸合成酶;APS:腺苷-5’-磷酰硫酸;PAPS:3′-磷酸腺苷-5′-磷酰硫酸。B: 微生物硫转运蛋白,根据Esther Aguilar-Barajas等人[12]和Eric Eichhorn等人[18]文献绘制;taurine:牛磺酸;alkanesulfonate:链烷磺酸盐。) 图1 微生物硫酸盐同化途径及硫转运蛋白Fig.1 Microbial sulfate assimilation pathway and sulfur transporter
(2)活化 进入生物体的硫酸盐在基因cysDN编码的ATPS(ATP硫酰化酶,EC 2.7.7.4)作用下活化为腺苷-5’-磷酰硫酸(APS)。许多细菌中,如大肠杆菌和肠道沙门氏菌中,cysC编码的APS还原酶(EC 2.7.1.25)进一步作用于APS,其将APS转化为3′-磷酸腺苷-5′-磷酰硫酸(PAPS)。
(3)还原 PAPS通过cysH编码的PAPS磺基转移酶(PAPS还原酶,EC 1.8.4.8)从PAPS中释放亚硫酸盐和3’,5’二磷酸腺苷酸(PAP),再通过亚硫酸盐还原酶(EC 1.8.1.2)将亚硫酸盐还原为硫化物。
(4)半胱氨酸合成 硫酸盐同化的最后一步是将硫化物整合入半胱氨酸。硫化物与O-乙酰基-L-丝氨酸(OAS)结合形成L-半胱氨酸,该反应由两种OAS裂解酶(也称半胱氨酸合成酶,EC 2.5.1.47)同功酶CysM或CysK中的一种催化[16]。
3 微生物硫酸盐同化途径的调控
3.1 微生物硫酸盐同化途径的代谢调控
生物具有保持相对恒定细胞内环境的能力。生物存在代谢和遗传控制机制,以使生物体适应胞外环境的变化。细胞控制机制通过关键酶活的调节(激活或抑制)实现[7]。一种方式是通过与一些代谢物和共价酶修饰的非共价相互作用组成的短期(生物化学)机制,是酶活性的调节;另一种方式是由酶的合成和降解速率变化组成的长期(遗传)机制,是酶合成的调酶活性的调节是生物体一种快速、精细的调节方式。目前已有研究中,对微生物硫酸盐同化途径代谢调节的研究主要集中在外加(减)硫源或中间代谢物对关键酶活性的影响方面。
其次,中间代谢产物对关键酶活的影响。在结核分枝杆菌Mycobacteriumtuberculosis中,硫活化阶段的产物PAPS可在磺基转移酶的作用下产生PAP, PAP过量会抑制磺基转移酶的活性,因而抑制硫酸盐活化过程[22]。给Saccharomycescerevisiae提供富含半胱氨酸、甘氨酸的培养基导致GSH合成速度短期快速增加,但GSH合成酶基因表达下降。几小时后GSH1表达水平和GSH合成速率恢复基础水平。这些结果表明,较高浓度的GSH合成的控制发生在酶水平而不是转录水平。另一方面,培养基中半胱氨酸的补充导致硫酸盐同化途径相关基因下调[23]。这表明硫酸盐同化途径一般受其中间代谢产物或半胱氨酸衍生物的抑制。
3.2 微生物硫酸盐同化途径的遗传调控
除了硫源或中间代谢物对硫酸盐同化途径关键酶活的影响,微生物硫酸盐同化途径也受到相关功能基因遗传调控。
大量研究通过构建突变体的方式结合微生物硫源利用能力分析进行研究。Tomas Linder[24]研究了巴斯德毕赤酵母Pichiapastoris无机硫酸盐同化途径中的两个关键基因MET3(编码酶ATP硫酸化酶)和MET5(编码亚硫酸盐还原酶),通过构建缺失突变体检测利用氨基磺酸盐和其他硫源的能力。MET3基因的缺失不影响对L-甲硫氨酸,亚硫酸盐,甲磺酸盐或牛磺酸的利用,但在硫酸盐,甲基硫酸盐和氨基磺酸盐上的生长抑制。缺失MET5基因菌株在除L-甲硫氨酸以外的所有测试硫源上均不生长。这些结果表明,氨基磺酸的分解代谢在其同化前通过硫酸盐中间体进行。Zu-Jun Lu[25]等人通过突变体创建和单一硫源(硫酸盐和甲硫氨酸)利用分析的方法识别了费氏中华根瘤菌SinorhizobiumfrediiHN01中一个关于硫酸盐同化的操纵子,其中基因cysG,cysI与其他根瘤菌同源,并且新注释一个基因cysII。Marco Fischer等人[26]也通过这种方式识别了放线杆菌StreptomycescoelicolorA3中的sirA基因,其用于编码亚硫酸盐还原酶。S.frediiWGF03的SiRI 突变菌株不能用利用亚硫酸盐作为唯一硫源,而互补菌株CSiRI 与野生型菌株却可以,结果证实了亚硫酸还原酶确实与亚硫酸盐同化途径有关[27]。宋张扬等人[28-29]利用转座子插入法构建了S.fredii15142的cysDN突变菌株,在利用硫源时受到限制,不能利用硫酸盐,能利用其他无机硫源(亚硫酸盐和硫代硫酸盐)和有机硫源(半胱氨酸和甲硫氨酸),而互补菌株基本不受影响。在S.frediiWGF03[30]、S.frediiHN01、苜蓿中华根瘤菌14 500(均为快生根瘤菌)亦有相似结果,但是在大豆慢生根瘤菌15606(属慢生根瘤菌)呈现不同趋势,其cysDN突变株利用硫酸盐能力并未改变。推测这两种根瘤菌的硫活化机制存在不同。这些研究表明,对微生物硫酸盐同化途径相关基因进行突变后,微生物硫源利用能力发生改变,从而证实了相关基因在硫酸盐同化途径中的功能。
另外,近年来相关基因或蛋白调控硫酸盐同化途径的也有报道。丁香假单胞菌Pseudomonassyringae和恶臭假单胞菌Pseudomonasputida中fprA对硫酸盐同化具有调控作用[31]。伯克霍尔德氏菌Botkholderiaphytofirmans中PRF在硫酸盐同化中发挥作用[32]。大肠杆菌中cysB对S-硫代半胱氨酸转运子YdjN有调控作用;同时cysB也调控硫酸盐同化过程[33],不动杆菌中亦如此[16]。
4 重金属胁迫下微生物硫酸盐同化途径的调控
4.1 微生物硫酸盐同化途径在重金属抗性中的调控作用
金属的二重性(微生物对金属的代谢需求和金属对微生物的潜在毒性)使得微生物发展出一系列金属稳态和抗性系统,以保持体内金属离子微妙的平衡。
相较于传统研究方法,例如测量生物群中的浓度和生化、生理和行为变化,组学方法可以提供对生物途径和调节细胞命运、发育和进程的分子更深入的见解。近年来,随着组学方法的传播,特别是转录组和蛋白质组学方法的传播,提供了研究微生物全基因组和蛋白质表达响应的可能性[8]。组学方法阐明了细胞过程,到目前为止,已有多篇组学相关文章指出硫酸盐同化在重金属抗性中具有重要作用。Rodrigo J. Alma?rcegui等人[34]通过定量蛋白组学分析40 mM CuSO4存在下,嗜酸氧化亚铁硫杆菌AcidithiobacillusferrooxidansATCC23270中的蛋白差异表达,结果表明CysN、CysD-2、CysJ、CysNC、CysI表达上调。Camila Salazar等人[35]对嗜酸氧化亚铁硫杆菌菌株A.ferrooxidansD2对325 mM铜胁迫的响应的微阵列研究中,结果表明cysD,cysI,cysN分别上调了4.39,2.61,3.36倍。Garrett H.等人[36]研究了金属球菌Metallosphaerasedula对多种金属(Co,Cu,Ni,Zn,U)的反应,发现4和8 mM铜刺激cysNHI基因显着上调,而其他金属刺激不上调。S.cerevisiae在镉胁迫下,蛋白组分析表明,亚硫酸盐还原酶(SiRB)亚基增加了2.5倍[37]。本实验室对一株铜抗性菌的转录组测序研究表明,硫酸盐同化途径在铜胁迫下显著性表达并上调,另外铜胁迫的菌株培养基中添加硫酸盐有利于菌株抗性的增强。这些研究表明,硫酸盐同化途径在重金属抗性中具有重要作用。
微生物硫酸盐同化途径在金属胁迫下的调控作用的主要体现在硫酸盐同化途径的激活和抑制两方面,如图2。
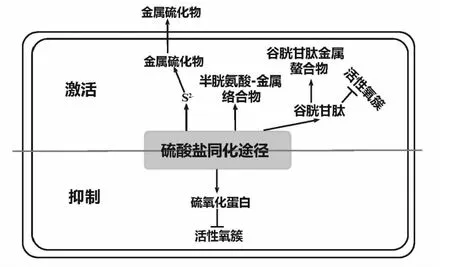
(↑相关产物的增强;↓相关蛋白的减弱;┤抑制、清除) 图2 微生物硫酸盐同化途径在重金属抗性中的调控Fig.2 Regulation of microbial sulfate assimilation pathwray in heavy metal resistance
(1)激活作用
一些金属可与还原态S2-形成硫化物。由于硫化物是生物学上不可利用的沉淀物,硫化物是金属解毒态的优选形式。沼泽红假单胞菌Rhodopseudomonaspalustris具有通过硫酸盐同化途径合成半胱氨酸的能力,在低浓度Pb2+、Cd2+存在时,半胱氨酸脱硫酶酶活性增强,可诱导半胱氨酸生成S2-,继而形成PbS、CdS沉淀,然后排出体外[38]。在一些光合自养微生物亦有相似研究,如绿藻类,红藻类,蓝藻等在受到金属胁迫时,均能够在有氧条件下合成硫化物,形成ZnS[39]、CdS[40]和HgS[41]等。同时在这些生物中半胱氨酸脱硫酶活性亦有增强,向培养物中补充某一种硫源(硫酸盐或亚硫酸盐或半胱氨酸)时会促进菌体的生长与金属硫化物沉淀的生成。这些研究表明微生物在受到金属胁迫时会通过增多金属硫化物沉淀的形成来达到解毒的目的,而半胱氨酸脱硫酶活性的增强说明形成金属硫化物的硫来自于半胱氨酸,硫源的补充实验说明,硫酸盐同化途径是半胱氨酸的主要来源。因而,微生物的硫酸盐同化途径在重金属的抗性与解毒中可通过形成金属硫化物沉淀的方式发挥作用。
为了限制游离金属浓度,特定螯合剂的合成和金属络合物的隔离是非常重要的[42]。一般,金属对硫醇(-SH)基团具有高亲和力,生物体内常见硫醇物质有半胱氨酸、谷胱甘肽、金属硫蛋白和植物螯合剂等。通常,半胱氨酸的巯基与几乎所有金属均能强烈结合,使游离半胱氨酸成为金属离子的有效螯合剂。半胱氨酸的巯基可与Hg2+、Cu2+和Ag+等重金属离子结合形成不溶性硫醇盐[43]。一些蛋白中的半胱氨酸残基也能与金属结合,如已有研究表明FtsL在大肠杆菌E.coliK12细胞分裂中发挥结构作用,但FtsL蛋白的半胱氨酸残基对于细胞分裂并不是必需的,却影响菌株对Zn的抗性,推测缺乏半胱氨酸残基的FtsL蛋白通过增加细胞质膜的渗透性导致菌株的Zn敏感性[44]。亦有研究发现半胱氨酸能有效络合土壤中的Zn,且这种能力强于谷胱甘肽,但由于半胱氨酸的不稳定性,这种络合会随时间衰减。另外,当半胱氨酸与具有氧化还原活性的金属结合时,被迅速氧化,还原的金属可能发生芬顿反应并形成有毒性作用的羟基自由基[42]。由于游离半胱氨酸可能是有毒的,转化为肽使得它在细胞中保持低浓度。通过半胱氨酸与谷氨酸、甘氨酸的偶联形成GSH的方式,细胞可以含有低浓度的半胱氨酸和相对高浓度的GSH,而不会引发有害的芬顿反应[45]。GSH是一种具有特殊生物学功能的由谷氨酸、半胱氨酸及甘氨酸组成的三肽,含有氨基、巯基、羧基和酰胺基等多种配位基团。一方面,GSH 是细胞内可以直接作用金属离子的螯合剂,具有强烈亲硫特征的金属元素,如镉,汞,铅和砷,可以和GSH 分子中的巯基结合,从而达到解毒的作用。另一方面,GSH可以清除因重金属胁迫产生的活性氧簇(ROS)[46]。
目前,金属胁迫条件下关于硫酸盐同化途径的研究,主要是从硫化物生成、硫醇物质含量、相关酶活性和相关基因表达水平几方面进行。A.ferrooxidans在Zn2+存在下,硫酸盐同化途径的基因(cysDEMH)与相应的酶表达均显著性提高,胞内cys和GSH也翻倍增加;在高浓度Cd2+胁迫下,硫酸盐同化途径相关基因普遍显著上调,cys和GSH随Cd2+浓度和暴露时间上升,半胱氨酸合成酶活性随Cd2+浓度呈现先升高再下降的趋势[47-48]。S.cerevisiae在Cd2+胁迫下,其硫转运蛋白SULl、SUL2和ATP硫酸化酶(ATPS)、APS还原酶METl4(APSK)、PAPS还原酶METl6(PAPSR)的表达均有上调现象[49]。深色有隔真菌在Cd胁迫条件下,75%的硫酸盐同化途径的基因上调,表明其参与了Cd的抗性机制,推测这种上调有利于谷胱甘肽硫转移酶与金属硫蛋白的合成,以绑定胞内多余游离Cd[50]。在这些案例中,菌体受到重金属胁迫后,硫酸盐同化途径相关酶的活性与基因表达均呈现增强与上调趋势,这预示硫酸盐同化途径的增强,同时胞内半胱氨酸、谷胱甘肽的含量也增多,这表明半胱氨酸与谷胱甘肽通过胞内螯合参与重金属抗性,同时谷胱甘肽还能清除由重金属产生的ROS以解毒。
(2)抑制作用
另外,重金属胁迫会抑制微生物的硫酸盐同化途径。A.caldus在高浓度的铜离子存在情况下,亚硫酸盐氧化酶和APS还原酶的活性会呈现下调趋势[51]。沙门肠杆菌Salmonellaenterica在160 μM 钴胁迫下,亚硫酸盐还原酶活性受到强烈抑制,钴还与cysG中的Fe原子发生竞争抑制硫酸盐同化途径;菌体内GSH含量降低。另外谷胱甘肽合成基因gshA缺失菌株无法正常合成谷胱甘肽,对钴的抗性降低[52]。这两个案例中,由于金属的胁迫,硫酸盐同化途径受到了抑制,这可能是由于这些金属到对相关的硫酸盐同化途径的蛋白产生毒性。在最近的研究中,大肠杆菌在0.1 mM Ag胁迫下,硫酸盐同化途径活性(cysCDHI)降低,因而减少了半胱氨酸生物合成等过程所需的硫化氢的量,故Ag与硫化氢的相互作用几率降低。蛋白CysH可与硫氧还蛋白相互作用,因此CysH的减少可以释放还原态硫氧还蛋白,从而可能提高在Ag胁迫下产生的活性氧存在下的抗性。
不同微生物硫酸盐同化途径对重金属胁迫有不同程度的响应,这可能与金属毒性和菌种耐受性相关。但是,仍可以初步了解到硫酸盐同化途径确实在重金属抗性中具有重要作用。
4.2 微生物硫酸盐同化途径调控在重金属污染微生物修复中的潜在应用
微生物长期在重金属存在的不良条件下会进化出一系列抗性机制来保证自身存活而不会对其生长和代谢产生影响。这些机制包括生物吸附、生物转化、生物累积、胞外沉淀和外排作用等。这些抗性机制是应用微生物修复重金属污染的基础,同时具有这些独特特征的微生物被证明是重金属污染环境场所的生物修复的理想材料。近年来的研究表明,硫酸盐同化也是微生物重金属抗性机制的一部分,在微生物重金属抗性中发挥作用。
目前,有关微生物硫酸盐同化途径在重金属抗性中的调控作用已有很多报道,如A.ferrooxidans在多种重金属胁迫下硫酸盐同化途径的不同调控模式。这表明微生物硫酸盐同化途径在重金属污染的生物修复中具有应用潜力。Clifford L. Wang等人[53-54]通过构建突变体的方式,使得大肠杆菌的硫酸盐同化途径能过量产生半胱氨酸,并且过量的半胱氨酸可被半胱氨酸脱硫酶转化为硫化物。这种大肠杆菌工程菌可在有氧条件下将镉沉淀为硫化镉沉积在细胞表面上,并且这种硫化镉沉淀是在细胞壁表面形成的。这项研究是应用硫酸盐同化途径的遗传工程菌以将重金属沉淀为金属硫化物的首次报道,但是相关的应用研究还不多。现在通过构建基因工程菌的方式强化微生物硫酸盐同化途径应用于重金属的生物修复这种途径还不是一种即刻修复环境金属污染的实用方法。如果基因工程途径用于从污染环境中去除重金属,可以考虑使用与环境更相关的生物。此外,还需要其他手段技术的应用以使这种基因工程生物在异质微生物环境中具有生长优势与竞争力。同时,硫化物具有结晶比较细小, 难以沉降的特点[55],如何改善硫化物的沉降性能值得进一步研究[56]。
5 结论与展望
微生物硫酸盐同化途径是微生物获得硫源的有效途径,关于微生物硫酸盐同化的研究主要集中在机理调控。而近年来组学的发展,使得其在微生物金属抗性中的潜在作用得以挖掘。微生物在重金属胁迫下硫酸盐同化途径呈现了不同的表达趋势。已有一些研究针对微生物硫酸盐同化途径与重金属抗性的相互关系进行了研究,但还不够全面。因而,对于微生物硫酸盐同化在重金属胁迫条件下的深入研究一方面对于补充该微生物的金属抗性机制具有重要意义,另一方面对更广泛的应用遗传工程菌在处理环境重金属污染方面也具有指导意义。
猜你喜欢
食品与生物技术学报(2022年4期)2023-01-12
核农学报(2021年7期)2021-12-07
建材发展导向(2021年15期)2021-11-05
当代水产(2021年3期)2021-07-20
食品界(2019年2期)2019-03-10
陶瓷学报(2019年5期)2019-01-12
中南大学学报(自然科学版)(2016年2期)2017-01-19
中国煤炭(2016年1期)2016-05-17
中国学术期刊文摘(2016年4期)2016-02-13
中国资源综合利用(2016年7期)2016-02-03
